外源MT和EBR对干旱胁迫下烤烟幼苗的缓解效应
2019-11-14李冬王艳芳申洪涛马宜林王丽君赵世民刘领
李冬,王艳芳,申洪涛,马宜林,王丽君,赵世民,刘领*
1 河南科技大学 农学院,河南洛阳开元大道263号 471023;2 河南中烟工业有限责任公司技术中心,郑州市经开区第3大街9号 450000;3 河南省烟草公司洛阳市公司技术中心,河南省洛阳市开元大道246号 471023
干旱作为植物生长发育进程中遭受的主要逆境形式之一,对农作物造成的损失约占所有非生物胁迫造成的损失的一半,仅次于生物胁迫中病虫害所造成的损失[1]。尤其是21世纪以来,干旱的频率和强度增加已成为制约现代农业生产高效发展的全球性问题[2-3]。干旱胁迫是影响植物生长和代谢的主要逆境因子之一,会抑制细胞分裂,致使细胞膜出现龟裂空隙且透性增大,使细胞内产生过量自由基,打破活性氧的代谢平衡系统,加剧膜脂过氧化程度,造成细胞内蛋白质与酶等生物大分子失活,而损坏细胞膜的结构与功能[4]。此外,干旱胁迫会阻碍大气中的CO2进入叶片,导致植物的光合作用受到抑制,降低叶绿体对光能的吸收和转化效率,吸收的光能超出光合机构的利用范围,过剩的光能使光合机构受到破坏,最终导致植物生长受到限制乃至死亡[5-6]。烤烟是我国重要的经济作物之一,起源于雨量充沛的热带、亚热带地区,整个生育期内对干旱胁迫颇为敏感,而我国大部分烟区受地理位置和气候环境限制,处于水资源短缺状态,尤其幼苗移栽期处于干旱频发时段,烟苗常因遭受水分胁迫而无法正常生长,严重制约了烤烟的产量和品质[7-8]。因此,研究如何提高烤烟幼苗抵御干旱胁迫的能力,已成为烤烟生产中亟需解决的问题[9]。
应用外源物质调控植物抗逆性是目前最为高效易行的方法之一。近年来,植物生长调节剂如褪黑素、油菜素内酯等在改善作物品质、提高作物抗逆性等方面的研究日益受到人们的关注。油菜素内酯(brassinolide, BR)最初是从油菜花粉中分离出来的一种具有极强生理活性的物质,属于甾醇类植物激素,已被公认是第六大植物激素,具有增强植物抗性的生理功能[10]。BR能够调节植物根、叶细胞的分裂和伸长,维持较高的叶绿素含量,解除光对生长的抑制作用,提高光合性能,促进作物体内蛋白质及核酸的合成,延缓植物的衰老以及提高植物的抗逆性[11-13]。王雨婷等[14]研究表明,2, 4-表油菜素内酯能够通过提高葡萄幼苗抗氧化酶的活性,减少脂质过氧化作用,进而增强植株的抗旱能力。褪黑素(melatonin, MT)是一种吲哚类色胺,化学名称为N-乙酰-5-甲氧基色胺,分子式为C13H16N2O2,是一种类激素物质,普遍存在于动植物和微生物体内,已被证实是目前已知抗氧化能力最强的内源性自由基清除剂[15-16]。褪黑素具有较高的亲脂性和亲水性,能够很容易地穿过细胞质进入细胞核发挥抗氧化作用,与羟基自由基和过氧自由基发生反应,从而清除植物体内的活性氧,维持细胞膜功能的稳定,促进光合作用,缓解不同逆境胁迫对植物体的损伤,增加其对干旱环境的适应性[17-19]。
外源物质褪黑素与油菜素内酯均为新型的植物生长调节物质,虽然在提高作物抗逆性方面的研究已有大量报道,但是两者在缓解作物干旱胁迫方面是否具有协同效应的研究尚未见报道。鉴于此,本研究以烤烟幼苗为材料,在营养液水培中,通过喷施适宜浓度的MT与EBR,探究两者互作对干旱胁迫下烤烟幼苗生长、生理生化及光合荧光特性的影响,以期揭示MT与EBR协同作用对植物抗旱性的调节机制,为农业生产中施用外源物质提高作物抗旱性提供理论依据和新思路。
1 材料与方法
1.1 材料和试剂
供试烤烟品种为‘豫烟6号’,烟草种子由洛阳市烟草公司提供。2, 4-表油菜素内酯(2, 4-epibrassinolide, EBR)购于上海源叶生物科技有限公司,褪黑素(MT)购于美国Sigma公司,试验用模拟干旱胁迫的聚乙二醇-6000(PEG-6000)购于上海国药集团化学试剂有限公司。
1.2 试验设计
本试验在河南科技大学农学院旱作与节水生理生态重点实验室进行。试验种子用10% H2O2表面消毒10 min,蒸馏水冲洗后浸种8 h,然后在育苗盘中发芽,放入人工气候培养箱中,设置光照强度为4000 Lx,昼/夜温度(28/20±2)℃,光照周期为14 h/10 h,当烟苗生长至四叶期时,选取形态长势基本一致的烟草幼苗,用蒸馏水洗净根部后移入盛有Hoagland营养液(pH5.7±0.2)的不透明水培箱中(规格:41 cm×31 cm×14.5 cm),每盆定苗20株,待烤烟幼苗培养至五叶一心期进行处理。
试验共设5个处理:(1) CK:(Hoagland营养液+喷施蒸馏水);(2) T1:干旱处理(Hoagland营养液+15% PEG-6000+喷施蒸馏水);(3) T2:(Hoagland营养液 +15% PEG-6000+ 喷施 0.2 mmol∙L-1MT);(4) T3:(Hoagland营 养 液 +15% PEG-6000+喷 施0.2 mg∙L-1EBR);(5) T4:(Hoagland营养液+15%PEG-6000+ 喷 施 0.2 mmol∙L-1MT+ 喷 施 0.2 mg∙L-1EBR)。每处理20株,重复3次。Hoagland营养液中加入含15%(W∙ V-1)的PEG-6000以模拟中度干旱胁迫(渗透势约为-0.3 MPa),每2 d更换一次处理液。外源MT和EBR浓度均为前期预备试验筛选确定出的最适浓度。外源MT、EBR溶液或蒸馏水于干旱处理当天均匀喷施在叶片正反两面,至叶面完全湿润且无溶液下滴为宜,EBR溶液喷施时间为上午8:00,为防止强光降解MT,MT溶液喷施时间定于20:00,每天喷施1次。于处理6 d后,取烤烟幼苗自上而下第2~3片真叶进行光合荧光参数及各项生理生化指标的测定,每个指标测定重复3次,取平均值。
1.3 测定项目与方法
1.3.1 生物量及根系形态指标测定
每处理随机选取5株烟苗,整株从盆中取出,用吸水纸将叶片和根系水分吸干,从根茎结合处剪断,分别称量地上部鲜重和地下部鲜重。采用EPSON 1680型扫描仪(Seiko Epson, Japan)及WinRHIZ0 2008根系分析软件(Regent, Canada)对根系总长度、根系表面积、根系平均直径和根尖数进行分析。
1.3.2 叶绿素相对含量的测定
采用SPAD-502叶绿素仪(Konica Minolta,Japan)测定叶绿素相对含量(SPAD值)。
1.3.3 光合参数的测定
采用LI-6400型光合作用仪(LI-COR, USA)于上午09:00-11:00进行测量。测定时选择Li-6400-02B红蓝光光源叶室,温度为25℃,使用开放式气路,空气相对湿度为50%~70%,设定有效光合辐射(PAR)为 800 μmol∙ m-2∙ s-1,CO2浓度为 400 μmol∙ mol-1。净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、蒸腾速率(transpiration,Tr)和胞间CO2浓度(intercellular CO2concentration,Ci)由光合仪直接测定;并计算瞬间水分利用率(water use efficiency, WUE)即Pn/Tr,气孔限制值(stomatal limitation index,Ls)即1-Ci/Co(Co为设定的CO2浓度)。

表1 外源MT和EBR对干旱胁迫下烤烟幼苗生长的影响Tab.1 Effects of exogenous MT and EBR on growth of flue-cured tobacco seedlings under drought stress
1.3.4 叶绿素荧光参数的测定
采用PAM—2100便携式调制荧光仪(Walz,Germany)进行测定。先测定光下最大荧光(Fm')、稳态荧光(Fs)和最小荧光(Fo');然后将同部位烤烟幼苗叶片暗适应20 min,测定初始荧光(Fo)、最大荧光(Fm)。通过以上测定的叶绿素荧光参数计算出光系统Ⅱ(PSⅡ)最大光化学效率(Fv/Fm)=(Fm-Fo)/Fm、PSⅡ实际光化学效率(ΦPSⅡ)=(Fm'-Fs)/Fm'、光化学猝灭系数(qP)=(Fm'-Fs)/(Fm'-Fo')和非光化学猝灭系数(NPQ)=(FM-Fm')/Fm'。测定日期同光合参数。
1.3.5 生理生化指标的测定
根系活力采用氯化三苯基四氮唑(TTC)法测定,超氧化物歧化酶(superoxide dismutase, SOD)活性采用氮蓝四唑光化还原法测定,过氧化物酶(peroxidase, POD)活性采用愈创木酚法测定,过氧化氢酶(catalase, CAT)活性采用紫外吸收法测定,丙二醛(malondialdehyde, MDA)含量采用硫代巴比妥酸比色法测定[20],超氧阴离子产生速率参照王爱国等的方法[21]。
1.4 数据统计分析
采用Microsoft Excel 2016进行数据处理和分析,采用IBM SPSS 22.0软件进行方差分析,并运用Duncan's 法进行差异显著性检验(α=0.05),以平均值±标准误(Means±SE)表示,用Origin 9.0软件进行绘图。
2 结果与分析
2.1 外源MT和EBR及其互作对干旱胁迫下烤烟幼苗生物量及根系形态的影响
由表1可以看出,与CK相比,各处理烤烟幼苗的地上部鲜重、地下部鲜重、总根长、总根表面积、根平均直径和根尖数均显著降低,说明干旱胁迫抑制了烤烟幼苗的生长。与T1相比,T2、T3和T4地上部鲜重分别提高了13.34%、22.14%和30.93%;地下部鲜重分别提高了18.97%、24.14%和29.31%;总根长分别提高了25.19%、31.97%和37.50%;总根表面积分别提高了23.41%、35.87%和53.67%;根平均直径分别提高了50.00%、63.64%和86.36%;根尖数分别提高了43.40%、44.11%和53.14%。说明,MT、EBR单独和复配均能显著缓解干旱胁迫对烤烟幼苗生长的抑制作用,促进根系的生长发育,且以MT与EBR复配处理效果最为显著,具有协同效应,其中EBR对植株生长的促进效应强于MT。
2.2 外源MT和EBR及其互作对干旱胁迫下烤烟幼苗叶绿素相对含量的影响
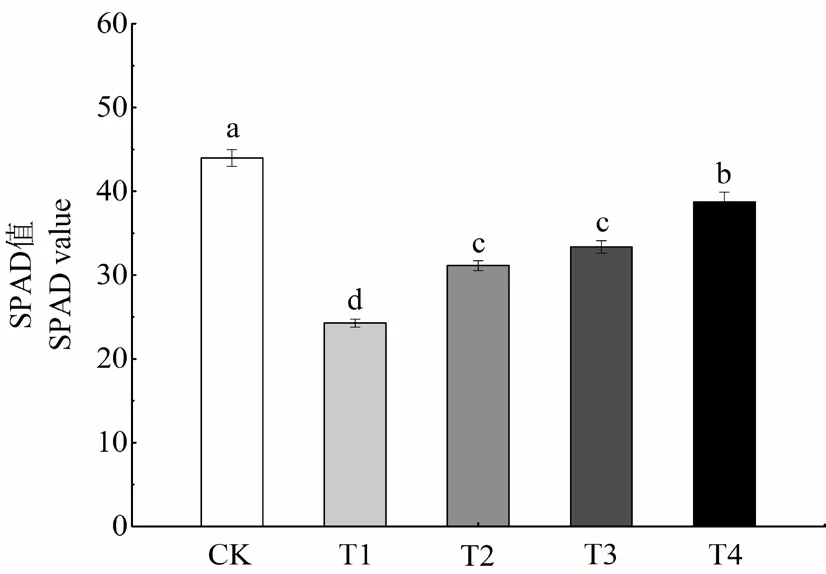
图1 外源MT和EBR对干旱胁迫下烤烟幼苗SPAD的影响Fig.1 Effects of exogenous MT and EBR on SPAD of flue-cured tobacco seedlings under drought stress
由图1可知,与CK相比,T1的叶绿素含量降低了44.81%,说明干旱胁迫显著降低了烤烟幼苗的叶绿素相对含量。与T1相比,T2、T3和T4的叶绿素含量分别提高了28.30%、37.50%和59.62%。说明外源MT、EBR单独和复配处理均能显著缓解干旱胁迫对烤烟幼苗的叶绿体结构的破坏作用,提高叶片光合色素含量,促进植株对光能的吸收及利用,且以MT和EBR复配处理的效果最佳。
2.3 外源MT和EBR及其互作对干旱胁迫下烤烟幼苗光合参数的影响
由图2可知,干旱胁迫抑制了烤烟幼苗的光合作用能力。与CK相比,T1的Pn、Tr、Gs、Ls和WUE分别降低了79.88%、52.26%、77.32%、53.82%和64.32%,Ci提高了52.12%。与T1相比,T2、T3和T4的Pn分别提高了210.87%、183.27%和293.09%;Tr分别提高了67.78%、52.68%和78.11%;Gs分别提高了229.29%、285.11%和284.74%;Ls分别提高了79.60%、73.88%和94.18%;WUE分别提高了77.31%、70.70%和122.80%;Ci分别降低了21.72%、23.40%和27.69%。说明,外源MT、EBR单独和复配处理均能有效提高干旱胁迫下烤烟幼苗的光合能力,且以MT和EBR复配处理的效果最佳。
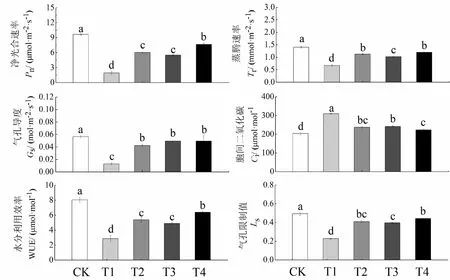
图2 外源MT和EBR对干旱胁迫下烤烟幼苗光合特性的影响Fig.2 Effects of exogenous MT and EBR on photosynthetic characteristics of flue-cured tobacco seedlings under drought stress

图3 外源MT和EBR对干旱胁迫下烤烟幼苗叶绿素荧光参数的影响Fig.3 Effects of exogenous MT and EBR on chlorophyll fluorescence parameters of flue-cured tobacco seedlings under drought stress
2.4 外源MT和EBR及其互作对干旱胁迫下烤烟幼苗叶绿素荧光参数的影响
由图3可知,与CK相比,T1的PSⅡ最大光化学效率(Fv/Fm)、PSⅡ实际光化学效率(ΦPSⅡ)和光化学猝灭系数(qP)分别降低了10.71%、31.70%和15.54%,非光化学猝灭系数NPQ提高了54.88%,说明干旱胁迫对烤烟叶片的光合机构造成了一定程度的损伤。与T1相比,T2、T3和T4的PSⅡ最大光化学效率(Fv/Fm)别提高了6.59%、6.12%和9.22%;PSⅡ实际光化学效率(ΦPSⅡ)分别提高了24.04%、24.01%和32.02%;光化学猝灭系数(qP)分别提高了9.62%、9.60%和12.11%;非光化学猝灭系数NPQ分别降低了19.72%、16.64%和22.53%。说明,外源MT、EBR单独和复配处理均能有效减缓干旱胁迫对烤烟幼苗的光抑制效应,提高光能的吸收利用效率和光合电子传递速率,且以MT和EBR复配处理的效果最佳。
2.5 外源MT和EBR及其互作对干旱胁迫下烤烟幼苗根系活力的影响
由图4可知,干旱胁迫显著降低了烤烟幼苗的根系活力,与CK相比,T1的根系活力降低了73.69%。与T1相比,T2、T3和T4的根系活力分别提高了72.59%、109.34%和139.25%。说明,外源MT、EBR单独和复配处理均能显著提高烤烟幼苗的根系活力,促进根系对水分、养分的吸收,以维持植株正常生长发育的能力,且以MT和EBR复配处理的效果最佳,其中EBR对根系活力的提升效果比MT显著。

图4 外源MT和EBR对干旱胁迫下烤烟幼苗根系活力的影响Fig.4 Effects of exogenous MT and EBR on root activity of fluecured tobacco seedlings under drought stress
2.6 外源MT和EBR及其互作对干旱胁迫下烤烟幼苗丙二醛含量和超氧阴离子产生速率的影响
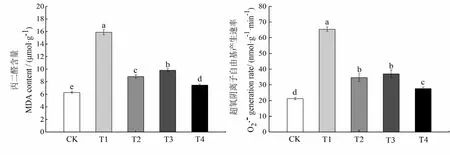
图5 外源MT和EBR对干旱胁迫下烤烟幼苗MDA含量和产生速率的影响Fig.5 Effects of exogenous MT and EBR on MDA content and production rate of flue-cured tobacco seedlings under drought stress
2.7 外源MT和EBR及其互作对干旱胁迫下烤烟幼苗抗氧化酶活性的影响
由图6可知,与CK相比,T1的POD和SOD活性分别提高了41.63%和12.96%,CAT活性无明显变化,说明干旱胁迫下烤烟幼苗体内的抗氧化酶系统有所增强。与T1相比,T2、T3和T4的POD活性分别提高了162.12%、138.48%和193.64%;CAT活性分别提高了54.66%、33.30%和89.70%;SOD活性分别提高了44.04%、28.97%和67.05%;说明,外源MT、EBR单独和复配处理均能有效提高干旱胁迫下烤烟幼苗的抗氧化酶活性,缓解氧化胁迫,其中以MT对抗氧化酶活性的提升效果大于EBR,MT和EBR复配处理的效果最佳。

图6 外源MT和EBR对干旱胁迫下烤烟幼苗SOD、POD和CAT活性的影响Fig.6 Effects of exogenous MT and EBR on SOD, POD and CAT activities of flue-cured tobacco seedlings under drought stress
3 讨论
干旱环境下植株为维持体内的水分,保持存活,通常会改变分子、细胞和生理代谢等方面来调节生长,在这一系列改变中植株的形态变化表现最为直观[22-23]。根系是植物吸收矿质营养和最先感应养分含量变化的器官,同时也是植物在干旱胁迫中最直接的受害部位[24]。根系活力是体现植物根系吸收功能、合成能力、氧化还原能力以及生长发育情况的综合指标[25]。李杰等[26]研究发现外源EBR能显著提高辣椒幼苗的地上部干鲜重和根系活力,增加根系总长、根表面积和分根数。叶君等[27]研究表明,外源MT能改善植株水分状况,显著增加小麦幼苗的生物量,促进干旱胁迫下小麦幼苗的生长发育。本试验结果表明,干旱胁迫下烤烟幼苗的生长受到显著抑制,外源MT和EBR单独及复配均能显著增强干旱胁迫下烤烟幼苗的根系活力,促进根系伸长,提高地上部与地下部的生物量,其中EBR的处理效果强于MT,且MT与EBR的复配效果最佳,表明MT与EBR之间存在显著的协同效应,对干旱胁迫起双重缓解的作用。推测其原因可能在于BR是调控植物根系生长发育和共生等的重要激素[28],而MT具有和生长素相同的前体物质及相似的生理功能[29],会通过诱导生长素的合成来促进植株体内源BR的生成,以此来增强对植物体内生理代谢活动的调节作用,保护根系结构免遭破坏,促进根系对水分和养分的吸收利用,抵御干旱胁迫对植株造成的损伤。
干旱胁迫能够通过渗透胁迫的方式对植物造成损害,导致植物体内超氧阴离子等活性氧(ROS)在细胞内过量积累,当积累量超过植物忍耐范围时,便会引起膜脂质的过氧化,丙二醛(MDA)作为质膜过氧化的最终产物,会破坏核酸、蛋白质和细胞膜等结构功能,从而对植物造成伤害甚至死亡[30-31]。逆境胁迫使植物体内活性氧产生增加,同时会刺激植物体内SOD、POD和CAT等抗氧化酶活性增加,以清除体内过多的自由基。本研究表明,干旱胁迫下,烤烟幼苗的MDA含量和产生速率均显著增加,POD和SOD活性显著提高,CAT活性无明显变化,说明干旱胁迫下,烤烟幼苗尽管能通过自身调节机制,提高抗氧化系统相关酶活性,却依旧无法清除体内过量的活性氧,而使其不可避免地遭受到氧化胁迫损伤。外源EBR能在一定程度上降低产生速率和MDA含量,提高POD、CAT和SOD活性,减轻膜脂过氧化程度,说明外源EBR能显著提高抗氧化酶活性,缓解干旱胁迫对烤烟幼苗造成的氧化伤害作用,提高细胞的渗透调节能力,保护细胞膜的完整性,与吴雪霞等[32]在高温胁迫下茄子幼苗上的研究结果相似,这可能是由于EBR能通过增强植物抗性相关基因的表达,提高植株抗氧化能力,减少自由基的含量,维持细胞膜结构的稳定性[33-34]。叶片喷施外源MT后能显著提高叶片POD、CAT和SOD活性,降低超氧阴离子产生速率和MDA含量,这与前人对甜瓜幼苗[35]的研究结果相一致,且缓解效果较EBR处理更为显著,其原因可能是MT能够通过诱导抗氧化酶相关基因的表达,减少生物大分子的降解来提高抗氧化酶活性,从而增强其对活性氧的清除能力[36]。此外,本研究发现,外源MT和EBR复配处理具有明显的互作关系,能显著提高烤烟幼苗的抗氧化酶活性,清除活性氧的产生,缓解干旱胁迫对烤烟幼苗细胞膜的损伤,减轻膜脂过氧化水平,从而增强植物抵抗逆境的能力,具体内在机制目前尚不清楚,可能是MT与EBR通过彼此共同作用来增强植物体内信号网络的传导或参与其它内源激素的合成,诱导抗逆基因的表达,从而抵御逆境胁迫对植物的生长抑制。
光合作用是植物生长发育的基础,也是植物合成有机物质和获得能量的根本来源[37]。叶绿素是植物进行光合作用的重要物质,在光合作用中起着吸收和转换光能的作用。叶绿素含量的高低在很大程度上反映了叶片光合作用的强弱,对植物生长发育具有重要作用[38]。干旱胁迫下光合作用受抑制是由多种因素共同作用的结果,原因可分为气孔限制和非气孔限制,判定依据主要是Ci、Gs和Pn的变化方向,当Pn与Ci同时下降时,气孔限制通常会导致Ci下降;非气孔因素会引起Ci升高[39-40]。本试验研究发现,干旱胁迫下烤烟幼苗的SPAD值、Tr、Gs、Ls和WUE均显著降低,Ci显著升高,表明干旱胁迫下非气孔限制因素起主导作用,导致CO2的固定和转化受到抑制,烤烟幼苗的叶绿体内的类囊体膜结构受到破坏,叶绿素含量降低,叶片光合器官受损,叶肉细胞光合活性下降。外源MT和EBR能显著提高干旱胁迫下烤烟幼苗的 SPAD 值、Tr、Gs、Ls和 WUE,降低Ci,且以MT和EBR复配处理最佳,存在显著的协同效应,说明外源MT和EBR能通过抑制叶绿素降解相关基因的表达,促进叶绿素的合成,参与气孔的调节,使与CO2固定相关的酶基因转录上调,增强植物对光能的吸收能力,提高光合性能,从而增强植物对干旱胁迫的适应性[41]。
在自然条件下,叶绿体吸收的光能主要通过光合电子传递、叶绿素荧光和热耗散3种途径来消耗,叶绿素荧光和光合作用在植物各反应中有着十分密切的关系,叶绿素荧光变化能够准确地检测出完整植株在逆境条件下的光合作用真实行为[42-43]。本试验结果表明,干旱胁迫下,烤烟幼苗的Fv/Fm、ΦPSⅡ和qP显著降低,NPQ显著提高,说明烤烟幼苗的PSⅡ反应中心受到损害,电子传递途径受到损伤,传递速率下降,吸收的光能用于光化学反应的比例降低,多余的光能则以荧光和叶黄素循环等形式耗散掉,这与金薇薇等[9]的研究结果基本一致。外源MT、EBR单独或复配均明显提高了干旱胁迫下烤烟幼苗的Fv/Fm、ΦPSⅡ和qP,降低了NPQ,且以MT和EBR复配效果最佳。说明对MT和EBR干旱胁迫下烤烟幼苗的PSⅡ反应中心有一定的保护作用,提高了电子传递速率,并通过增强光合电子传递过程中的热耗散能力来减少过剩激发能的产生,有效减轻PSⅡ反应中心因吸收过多的光能而引起的光抑制和光氧化作用,更好地保护光系统Ⅱ反应中心,提高植物的光合作用,缓解干旱胁迫对其造成的伤害。
4 结论
综上所述,在干旱胁迫下,烤烟幼苗的生长受到严重抑制。喷施MT和EBR均能显著提高烤烟幼苗抗氧化酶活性,降低体内活性氧积累,促进叶绿素的合成,减轻光抑制程度,保护光合机构免受损伤,改善光合性能,增强根系的吸收能力,缓解干旱胁迫对烤烟幼苗生长的抑制作用。与喷施单一外源物质相比,外源MT和EBR复配处理能显著减轻干旱胁迫对烤烟幼苗的损伤,且具有一定的交互作用。MT与EBR互作缓解干旱胁迫是一个复杂的调控机制,还有待进一步在分子、细胞水平上进行深入研究。
