化石
2019-11-08
寒武纪大爆发前夕动物的重要演化事件
身体两侧对称和分节现象的产生是动物演化史上极为重要的事件。分节的出现意味着身体的结构有了分区,功能上也就有了分工。但是,这类体型复杂的动物何时出现?自达尔文以来,一直是古生物学家和演化生物学家关注的焦点。"寒武纪大爆发"中出现了大量的以三叶虫为代表的具分节的两侧对称动物,因此,寒武纪常被称为"三叶虫时代"。长期以来,大家推测在寒武纪之前的埃迪卡拉纪(中国称为"震旦纪",6.3 亿-5.4 亿年前),它们应该有更加古老的祖先,但是一直没有找到可靠的化石证据。近期,研究者在中国三峡地区大约5.5 亿年前的地层中发现了一类特殊保存的新的动物化石,该动物的遗体和它最后行进的拖痕(印迹)同时保留在一块岩石上。这一新的化石证据表明在埃迪卡拉纪已出现了具有运动能力、并且身体分节的两侧对称后生动物。化石发现于湖北三峡地区埃迪卡拉系灯影组的石板滩生物群中。自2013 年开始,科研者用了近5 年的时间对新发现的化石进行详细的研究,并将其命名为夷陵虫(Yilingia)。

夷陵虫身体长条形,两侧对称,呈三叶形,具有明显的身体分节,也具有了前后和背腹的区别。更加奇特的是,部分标本中动物的实体与它的遗迹同时保存在一起,再现了一条行进中的虫子的"最后时刻"。这是一类全新的动物化石,在地质历史时期和现代都没有发现形态相同的动物,研究者以它的发现地点湖北省宜昌市夷陵区给它取了一个新名字" 夷陵虫",推测可能是环节动物或节肢动物。传统的埃迪卡拉生物群曾被认为是身体没有真正分节、缺乏运动能力、走向演化盲端的生物类群。虽然在埃迪卡拉纪也发现了许多生物运动产生的遗迹化石,但是,造迹生物是什么一直是一个谜。夷陵虫是目前在寒武纪之前发现的唯一的身体分节,具有运动能力,并可以形成连续的遗迹的两侧对称动物。夷陵虫的发现为两侧对称动物身体分节在埃迪卡拉纪的出现提供了直接的化石证据,也为探索该时期众多遗迹化石的造迹者提供了重要证据。该发现显示,能运动的底栖动物在埃迪卡拉纪已经出现,并对海底沉积物进行了改造。表明"寒武纪大爆发"时期以底栖动物为主体的生态系统在这一时期已经开始建立,并逐渐取代了前寒武纪统治地球数十亿年的微生物席基底,对地球表面系统造成了深远的环境和生态影响。
(Nature 2019,https://doi.org/10.038/s41586-019-1522-7)
揭密蝎子蜘蛛等螯肢类动物起源

螯肢动物是包括蝎子、蜘蛛、蜱螨和鲎等在内的超过11.5 万个物种的庞大节肢动物类群,对于寒武纪大爆发期间该类群动物是否已经起源一直不是很清楚。近期,研究者在寒武纪中期布尔吉斯页岩动物群中发现了一种莫里森虫(Mollisonia plenovenatrix)新物种,代表了目前已知最古老的螯肢类节肢动物。这项研究将包括螯肢动物的起源时间提前到了5 亿年前。尽管只有拇指大小,但莫里森虫很可能是凶猛的捕食动物。它长有一双卵形大眼睛、具长步行肢以及分别执行感觉、抓握、粉碎和咀嚼功能的多对附肢的"多功能"头部。但最重要的是,莫里森虫在其口部前方发育有一对小型的钳状附肢,称为"螯肢"。这对典型的附肢使得蝎子和蜘蛛等节肢动物能够击杀、夹持和切碎猎物,并以此命名这类节肢动物为"螯肢动物"。莫里森虫标本展现了确凿的"螯肢"特征。在这项发现之前,尽管一部分寒武纪节肢动物具有明显的似螯肢类特征,但螯肢类最典型的"螯肢"尚未在寒武纪化石中得到证实。此外,莫里森虫身体后部的附肢上具有三片鳃一样的构造,这和螯肢类身体后部成对的特化呼吸器官" 书鳃"或"书肺"相似。这进一步表明莫里森虫尽管是原始的螯肢类,但和现代的螯肢类在形态上已相当接近。
莫里森虫具有相当发育的步行足,因而被认为是生活在海底的捕食动物。由于莫里森虫和现代螯肢类形态相近,说明螯肢类节肢动物可能在寒武纪快速出现并迅速占据了其他节肢动物涉足较少的海底生态位。而螯肢类的起源很可能发生在寒武纪的更早期,即动物的"寒武纪大爆发"真正发生的时候。莫里森虫最早在一个世纪前由美国著名古生物学家查理斯·沃科特发现于加拿大的布尔吉斯页岩。然而,之前这类动物只有外骨骼被发现。此次的新研究采用了布尔吉斯页岩新化石点"大理石谷"的化石标本,保存了比沃科特发现的化石更精美的信息,首次揭示了莫里森虫的附肢等其他软躯体,进而揭示了莫里森虫的亲缘关系。这项发现进一步强调了寒武纪化石库的科学重要性。加拿大布尔吉斯页岩和中国澄江生物群等寒武纪化石库的核心重要性在于保存了" 寒武纪大爆发"中快速多样化的动物身体造型和最早的海洋动物群落。这些著名化石充分展现了动物的软躯体形态特征,这包括附肢、眼睛、消化道甚至 神 经 组 织。
(Nature 2019,DOI 10. 038/s 41586-019-1525-4.)
几丁虫更可能是一类独立的生物而非卵
几丁虫是早、中古生代(4.85~3.59 亿年前)海相地层中广泛存在的、具有机质壳壁的微体化石。由于其演化速率快,分布广,形态构造简单易识别,其在年代地层及生物地层对比中发挥着重要作用。然而,自1931 年被正式建名报道以来,关于其生物学属性确难有定论。1963年,伴随着几枚茧状保存标本的发现,几丁虫被越来越多的学者认为是某类后生动物的卵或卵囊,并逐渐作为主流学说被广泛接受。近期,研究者对美国俄亥俄州上奥陶统2 块微体样品中保存精美的一新种——维奥拉篱几丁虫(Hercochitina violana)——的形态大小进行了无差别度量。研究结果显示该种的壳体长度相差3 倍有余,且壳表纹饰及体室下部的收缩随着壳体变长愈发显著,其形态差异似乎更接近纵向生长而非种内差异。这一现象与现下主流的卵或卵囊学说相去甚远。为了进一步探索几丁虫形态差异的幅度及其生物学属性,研究者收集整理了前人发表的57 属(目前已知报道的所有属)378 种几丁虫的最大最小壳长比以及来自现代水生6 门10 纲45 种无脊椎动物卵的大小差异值进行了类比研究。结果显示,当度量标本的数量足够时,几丁虫壳体的大小差异明显大于现代水生无脊椎动物卵的大小差异,说明其为后生动物的卵的可能性极低,更可能为一类独立的微体生物。值得指出的是,在对几丁虫的度量数据进行局部加权回归回归分析时,目前唯一报道有茧状标本的链几丁虫属的大小差异值相对较小,从形态差异的角度并不能完全排除其为某后生生物卵的可能性。
(Proc.R.Soc.B286:20191270.)
甲虫独特的雌雄异型现象

昆虫纲是自然界中生物多样性最为丰富的类群,而甲虫(鞘翅目)是昆虫中多样性最高的一个庞杂类群,包括超过170 个现生科。可能由于化石保存缺陷或是标本较少,甲虫化石的分类系统学研究几乎都没有涉及雌雄个体之间的形态差异,因而对甲虫在有性生殖的长期演化过程中,其形态和结构的演化知之甚少。近期,研究者通过对缅甸琥珀中大量的甲虫化石进行系统研究,发现了6 枚保存精美的锯谷盗化石。它们雌雄个体罕见地保存了特化的触角、上颚等结构的形态差异,揭示了扁甲类甲虫雌雄异型的早期演化,并对理解现代的朽木树皮下生活的甲虫之间的趋同进化具有重要意义。锯谷盗科是扁甲总科的一个广布小科,包属,约500 个现生种。锯谷盗化石较为罕见,主要报道于始新世的波罗的海琥珀和法国瓦兹琥珀。研究者从缅甸琥珀(距今约1 亿年)中发现6枚保存精美的锯谷盗化石,包括3 个雄性和3个雌性个体,它们均属于已报道的长角原锯谷盗。根据扁平的体型、超长的触角和前胸侧脊等特征,原锯谷盗属能够很容易地与其他现生和化石锯谷盗区分开来,并与现生的Uleiota 属最为接近。通过多个雌雄个体的对比分析发现,长角原锯谷盗的雄性个体的触角明显长于雌性个体,雄性触角的第一个小节(柄节)也显著长于雌性个体,而且雄性个体发育极其特殊的上颚角,雌性则不发育该构造。雄性的上颚角在个体之间也存在一定的长度差异。更为有趣的是,研究发现朽木树皮下生活的某些亲缘关系较远的类群之间存在极其类似的雌雄异型现象。该发现代表一例典型的趋同现象,也是白垩纪中期缅甸琥珀中发现的首例甲虫雌雄异型化石记录,证明了森林生态系统中某些重要生态位在白垩纪中期已经被昆虫所利用,突显了早期与现代生态系统组成的相似性。
(Palaeoentomology2019,2:289-296)
副鸟类头骨可动性的演化
鸟类是唯一留存下的恐龙后裔。它们不仅跨过了约6500 万年前的白垩纪大灭绝,而且成功演化成了如今多样性最高的脊椎动物之一。鸟类的成功演化与其种种有关飞行的结构相关,也与其特殊的头骨构造密不可分。从非鸟兽脚类恐龙到真正的鸟类,头骨发生了一系列变化,如面部缩短、脑颅扩大、眼眶周围骨骼退化,以及骨骼纤细化和彼此间关节退化消失。尤其特殊的是,现生鸟类主要是新腭类具有高度可动的头骨,体现在其上颌能够相对于脑颅进行灵活地移动,而非鸟恐龙的头骨则不具备这样的可动性。
自19 世纪鸟类的头骨可动性被发现以来,研究者便对其相关的机制、演化、分异和功能大加关注。鸟类头骨可动性可能具有以下功能:扩大口裂的范围,加快喙部闭合打开的速度,提高头骨吸收冲击力的能力,提高取食的准确性。这一特性主要通过相对灵活的颧弓、方骨和腭区骨骼共同实现:方骨向前后方向移动时将推力传递给可动的颧弓和腭区骨骼,从而实现部分上颌相对于脑颅的上抬。腭区的翼骨-腭骨-犁骨系统可以从头骨后部向前部传递推力,因此在头骨可动性的实现中具有重要作用。由于早期鸟类极少保存有腭区骨骼,关于鸟类头骨可动性的起源及早期演化此前均知之甚少。
近日,研究者通过对会鸟(Sapeornis)和中国猎龙(Sinovenator)的头骨进行了高精度CT 扫描,并对腭区重要组成部分——犁骨进行了三维重建。同时研究者对现生鸟类的腭区进行了全面的形态学研究,并抽取现生鸟类各大支系代表的头骨进行了高精度CT 扫描和犁骨的三维重建,加上两件早期副鸟类新的犁骨模型共同进行了三维几何形态分析。结果显示现生鸟类中的古腭类沿袭了非鸟恐龙及早期鸟类的原始犁骨形态,而新腭类则进行了更多的改造。古腭类的头骨可动能力极其低,多样性程度也极低。由于犁骨的形态对于腭区骨骼的灵活性具有巨大的影响,以会鸟为代表的基干鸟类和古腭类的相似性显示早期鸟类可能头骨可动性同样很低,这一点也为新发现的会鸟外翼骨所证实。根据目前仅有的证据,研究者初步推测高超的头骨可动性可能伴随着古腭类和新腭类的分化,仅在新腭类中演化出现。这一特性可能赋予了新腭类取食策略上的优势和可塑性,使得它们在和古腭类同时面对大灭绝后的空白生态位时,最终脱颖而出。
(PNAS 2019,www.pnas.org/cgi/doi/10.1073/pnas.1907754116)
基干阔鼻猴脑的特征与类人猿的脑演化
人类的脑,无论是绝对容量还是相对容量,都比其他灵长类更大,沟回结构也更复杂。传统的观点认为,人类增大的大脑,是在演化的过程中逐渐积累的结果,总体上延续了灵长类相对脑容量逐渐增加、沟回结构逐渐变复杂的趋势。
在类人猿漫长的系统演化历史中,人类类型的脑从何时开始区别于其他类人猿、以何种速度增大、以及从何时开始可以称之为人脑,在此研究之前并没有明确的答案。近日,研究者以生活于南美洲的基干阔鼻猴类卡拉斯科智利猴(Chilecebus carrascoensis)化石为" 校正点",重建了阔鼻猴类和狭鼻猴类类人猿祖先类型的脑的特征,分析了类人猿脑的增大过程,进而试图回答上述关于人脑演化的问题。
动物的脑属于软组织,在动物死亡之后,很短的时间之内就会被微生物分解,不可能成为化石。但是,哺乳动物的脑是明显膨大的器官,在生长发育的过程中会在头骨的内壁上留下脑的印痕。类人猿因为往往比具有同等体重的其它哺乳动物有更大的脑,所以脑的增大更为明显,在头骨内壁上留下的印痕也就更清楚,常常可以保存大的血管和主要沟回的印痕,在整体形态上与包裹于硬脑膜的脑非常相似。
研究者利用高精度CT 扫描的方法,数字化地三维重建了卡拉斯科智利猴的头骨内壁的脑的印痕,发现它具有相对平扁的额叶,但是具有发育良好的枕叶,嗅球相对较小,并存在有多对沟回,整体形态上甚至与现生的阔鼻猴狨猴相当。智利猴化石发现于距智利首都圣地亚哥东南约100 公里的地方,埋藏于坚硬的火山碎屑岩之中。是目前已知保存最完整的基干阔鼻猴类类人猿,距今约2000 万年。

阔鼻猴是类人猿冠类群的两大分支之一,早在距今4000 万年以前,阔鼻猴类就与包括人类在内的类人猿的另外一个大支系,也就是狭鼻猴类,开始分道扬镳了。阔鼻猴类从此独立演化,平行于人类和狭鼻猴类发展出了很多脑部的进步特征,一些阔鼻猴类脑的相对大小也明显增大,与属于狭鼻猴的猿类相当。某些阔鼻猴,比如卷尾猴,会使用简单的工具,有人认为卷尾猴的智力水平甚至超过黑猩猩。
毫无疑问,阔鼻猴类是研究包括人类在内的狭鼻猴类脑演化的自然参照系,而卡拉斯科智利猴是这个参照系中目前已知最为关键的一个参照点。结果显示,阔鼻猴和狭鼻猴类的祖先具有相对大小相似但结构不同的脑,在4000 多万年的演化历史中,两个类群中都有脑容量增大的趋势,除人类支系以外,两者的增大比例非常相近,并且趋同地演化出了嗅球缩小、沟回增加等宏观特征。人类支系自700 万年前与大型猿类分开演化以来,脑容量的增加完全不同于其他类人猿的增加比例。相对于自身体重,并去除演化积累的因素,人类的相对脑容量是猿类的3 倍以上。可以说,自700 万年前人猿分开以后,人类支系最显著的特征就是脑按照不同于其它类人猿的演化模式而高速发展。
(Science Advances 2019,5:eaav7913)
破解印度熊和郊熊关系之谜

在中新世-上新世的大型熊类中,最出名的是印度熊和郊熊。二者个体巨大,分布广泛,发现历史悠久,是中新世-上新世的代表物种。虽然二者在很多特征上十分接近,但是以往并没有证据能够直接联系起二者。因此对于二者的演化关系也一直争议颇多,有的学者认为二者非常接近,是直系的亲属,有的学者认为二者没有任何亲缘关系,并认为印度熊属于大熊猫亚科,而郊熊属于半熊亚科。近期,研究者描述了一件来自美国亚利桑那州的郊熊化石,并建立了一新种,即亨氏郊熊。该种个体明显小于北美同时代的兄弟,也小于大部分旧大陆的郊熊,为已知个体最小的郊熊之一。更为重要的是,这件标本的下裂齿体现出了印度熊和郊熊过渡的特征,为解决二者关系提供了绝佳的契机。通过详细回顾非洲郊熊下裂齿的变异模式,研究者提出郊熊很可能起源于印度熊,二者同属于大熊猫亚科。但其中具体的演化关系,这一过程最早发生的时代和地点还有待进一步讨论。
(Journal of Mammalian Evolution 2019,https://doi.org/10.1007/s10914-019-09480-x)
"谜一样的恐龙"的早期演化

阿尔瓦雷斯龙类是一类高度特化的小型兽脚类恐龙,具有高度特化的前肢和修长的后肢,化石记录分布于中-上侏罗统至上白垩统,时间跨度近一亿年。阿根廷科学家约瑟夫波拿巴于1991 年报道了此类群的第一种恐龙——卡氏阿尔瓦雷斯龙。然而,此类群恐龙的后续研究波折不断,部分晚白垩世的属种被错误地鉴定为鸟类,这个类群在虚骨龙类的位置也迟迟不能确定。因此,这类恐龙也被称为"谜一样的恐龙"。随着更多化石的发现和兽脚类恐龙系统学研究进步,学界目前公认阿尔瓦雷斯龙类是一类相对原始的虚骨龙类。然而,受限于稀缺的早期演化记录和稀少的化石数量,阿尔瓦雷斯类仍然"充满谜团"。幸运的是,来自中国西部中晚侏罗世到早白垩世的化石记录,包括灵巧简手龙、赵氏敖闰龙、乌拉特半爪龙、彭氏西域爪龙等,逐步填补了阿尔瓦雷斯龙类演化历史中的大段空白。继这一系列重要发现之后,研究者于近日又详细描述并命名了侏罗纪阿尔瓦雷斯龙类一新属种——意外石树沟爪龙(Shishugounykus inexpectus)。这类恐龙的属名用以纪念此化石所产层位,也是世界上研究中-晚侏罗世恐龙动物群最为重要的组——石树沟组。种名"inexpectus"寓意这种恐龙是在石树沟组系列科考发掘当中的"意外之喜"。
意外石树沟爪龙是继灵巧简手龙和赵氏敖闰龙之后,中国新疆准噶尔盆地石树沟组地层中发现的第三种阿尔瓦雷斯龙类恐龙,也与它们一起构成了世界范围内已知最早的阿尔瓦雷斯龙类化石分布。这三种阿尔瓦雷斯龙类在形态和体型上都差异明显,表明阿尔瓦雷斯龙类恐龙在目前已知最早的化石记录就已具有较高多样性。阿尔瓦雷斯龙类高度特化的前肢,是它们最具代表性的特征。意外石树沟爪龙保存完好的前肢化石,为我们了解阿尔瓦雷斯龙类前肢演化打开了一扇新的窗口。研究发现,意外石树沟爪龙的前肢不仅具有与其他阿尔瓦雷斯龙类相近的特点,也具有与典型虚骨龙类前肢类似的特征。结合其他两类侏罗纪阿尔瓦雷斯龙类的前肢特征,这类恐龙前肢的早期演化可能呈"镶嵌式"演化。
(Scientific Reports 2019,9:11727)
南亚的印度熊类
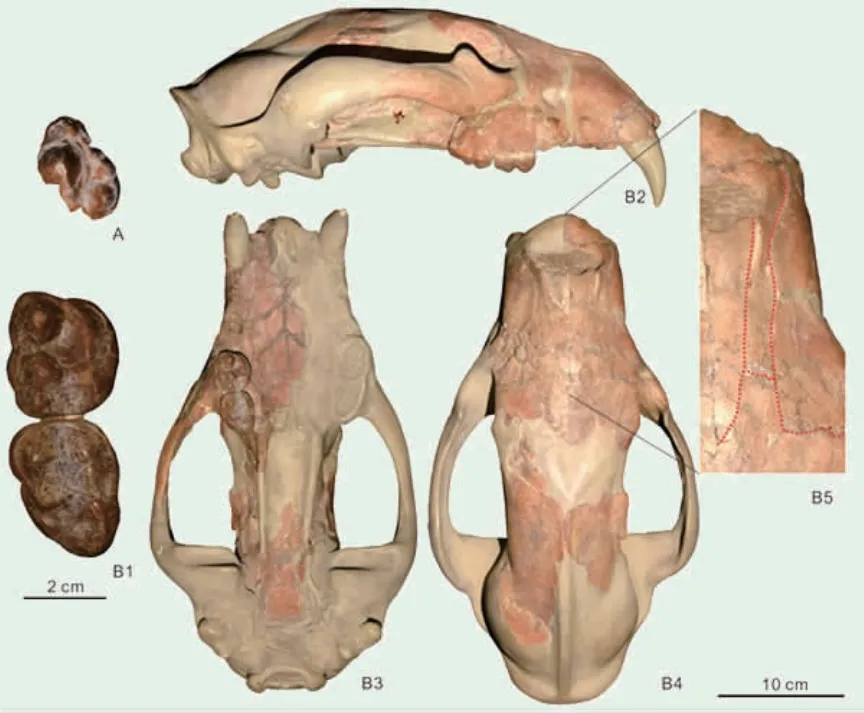
位于青藏高原南麓产地西瓦里克是亚洲最经典的化石产区之一,自从19 世纪上叶就有大量研究开展,至今仍然是国际古生物的研究热点区域。近期,研究者详细报道了来自耶鲁大学皮博迪博物馆馆藏的南亚印度熊及其他犬形亚目的化石。该批化石采集于1933 年,但多数都没有经过详细描述或报道。这批化石中保存最好的是一件印度熊头骨化石。印度熊是熊科大熊猫亚科成员,出现并繁盛于晚中新世(1130万-530 万年前)。和现代佛系的大熊猫不同,印度熊并非以竹子为主食,而和现代棕熊一样是杂食动物,并且拥有捕食大型猎物的能力。印度熊最早即发现在南亚(包括2 种,即盐岭印度熊和旁遮普印度熊),故而得名印度熊。然而南亚最初发现的印度熊化石保存较为残破,因此学术界对于南亚印度熊的性质,存在诸多争议。通过对新的头骨材料的研究,研究者发现印度熊模式种,盐岭印度熊和欧洲的阿提卡印度熊差别非常小,其中少量的差别,如体型稍大,M2具有后附尖,更可能代表同一个物种的地理变异,因此学者将盐岭印度熊归入阿提卡印度熊。南亚的另外一种印度熊,旁遮普印度熊则具有明显的特殊性,与任何印度熊都不同,反而接近郊熊。综合对比认为旁遮普印度熊最接近北美的舒氏"郊熊",二者的特征都介于典型的印度熊和郊熊之间,可能属于一个新属。综合来看,印度熊的模式种应该修订为阿提卡印度熊,而旁遮普印度熊不能归入印度熊。
(Historical Biology 2019, DOI: 10.1080/08912963.2019.1648449)
古神翼龙类最新研究成果
中国翼龙属于古神翼龙科,是一类没有牙齿的翼龙,其典型特征是鼻眶前孔大、前上颌骨脊组成的头饰从头骨的前端一直延续到头骨的后端。古神翼龙科成员化石相继在巴西、摩洛哥、中国和西班牙地区被发现。中国的古神翼龙类成员中一些属种的有效性还存在争议。2002年研究命名的董氏中国翼龙是中国最早发现的古神翼龙类。在其之后发现的" 谷氏中国翼龙"被认为是董氏中国翼龙的年轻个体。此外,在我国热河生物群还有一个一直存疑的"华夏翼龙",这类翼龙和中国翼龙的主要区别在于前上颌脊形状上的细微差别,大部分翼龙研究者认为它们脊冠之间的微小差异是由种间差别或性别差异引起的,而非属间差别,即"华夏翼龙"是中国翼龙的晚出同物异名。在2017 年出版的《中国古脊椎动物志》第二卷第四册翼龙类中初步总结和修订了中国发现的古神翼龙类,将"季氏华夏翼龙"作为董氏中国翼龙的晚出异名;具冠" 华夏翼龙" 和本溪" 华夏翼龙"放入中国翼龙属,保留原种名。
近期,研究者详细描述了一件保存完整的返祖中国翼龙化石新材料,这件标本产自于辽宁凌源四合当下白垩统。返祖中国翼龙最早由我国学者于2016 年命名为返祖"华夏翼龙"。研究者通过对新材料详细的对比研究,修订了返祖中国翼龙的系统学位置,认为返祖"华夏翼龙"应修订为返祖中国翼龙,归入中国翼龙,并补充修订了中国翼龙的头骨和头后骨骼特征。这一完整的返祖中国翼龙新材料的发现,同时还补充了中国翼龙属的骨骼特征, 增加了对其演化历史和多样性的认识,对探索中国古神翼龙属种内部的各种演化关系有非常重要的意义,也为今后研究古神翼龙科的起源奠定了坚实的基础。这一标本还罕见的保存了胃容物,进一步的研究将对这一翼龙类群的食性有较深入的了解。
(Anais da Academia Brasileira de Ciências 2019,91:e20180756)

小盗龙的食性

近期,研究者在一件白垩纪早期热河生物群的驰龙科新标本——赵氏小盗龙(Microraptor zhaoianus)的胃中,发现了一个保存近乎完整的、尚未被消化的蜥蜴个体。这是迄今报道的第四件胃中保存食物的小盗龙标本。经观察分析,研究者认为这件蜥蜴标本代表一个新属种,将其命名为王氏因陀罗蜥(Indrasaurus wangi)。属名" 因陀罗" 来自古印度的吠陀传说,古神"因陀罗"在一场著名的战役中,被龙(这里暗指小盗龙)完整地吞入肚中;种名"王氏"则命名自中国古动物馆馆长、中科院古脊椎所研究员王原,旨在向他多年来在古两栖爬行动物研究和古生物学科普工作中所做出的杰出贡献表达敬意。系统发育分析结果显示,王氏因陀罗蜥与白垩纪的蜥蜴有较近的亲缘关系,而与现代蜥蜴的亲缘关系较远。值得提出的是,这种蜥蜴的牙齿不同于白垩纪热河生物群内的其他蜥蜴,从而增加了热河生物群蜥蜴类的生物多样性,也暗示了这种蜥蜴的食性可能较其他蜥蜴有所不同。结合之前的研究可知,小盗龙是一种肉食性恐龙,食谱包含鸟,鱼,哺乳动物和蜥蜴,指示小盗龙是一种机会主义捕猎者。体内的蜥蜴被其从头到尾、整体吞入体内,这种进食方式与现在的食肉鸟类捕食蜥蜴的方式相似。即使生活在侏罗纪时期的伤齿龙类——近鸟龙已经具备了与现代鸟类相似的消化特征(将难以消化的骨骼、毛发、昆虫硬壳等,以食团的形式吐出),但是小盗龙明显不具备这种消化方式。这表明在从恐龙到鸟的演化过程中,相近的类群存在平行演化,一些性状可能独立地演化了很多次。
(Current Biology 2019,29:P2423-2429.E2)
杂志排行

生物进化的其它文章
- 审美与进化
- 正盘山怀旧
- ——毕生追逐的梦">" 寻根溯源"
——毕生追逐的梦 - 河狸,从古至今的传奇
- 行为
- 美国卡耐基自然博物馆新生代哺乳类展示掠影