冷鲜牛肉贮藏中菌群结构及优势菌致腐性的分析
2019-10-08顾春涛毕伟伟朱军莉
顾春涛,毕伟伟,朱军莉*
(浙江工商大学食品与生物工程学院,浙江省食品安全重点实验室,浙江 杭州 310018)
牛肉是中国仅次于猪肉的第二大消耗肉品,具有较好的行业前景。冷鲜肉能良好地保持肉品风味和营养价值,逐渐成为肉类消费的主流。冷鲜牛肉品质与其微生物种类及数量密切相关,在冷链贮运中由于嗜冷微生物繁殖代谢不断积累醛、酮、胺、有机酸等物质[1],导致牛肉腐败变质。牛肉微生物菌群在贮藏过程的增长,导致牛肉品质下降,与牛肉的货架期密切相关[2]。在肉品复杂的微生物菌群中,只有一种或几种特定腐败菌在腐败进程中起着主导作用。
生鲜食品贮藏过程中菌群落鉴定主要依赖培养,它是实验室鉴定微生物最常用的方法,能简便地计算出样品中活菌的数量,并且是检测某些特定腐败菌的标准方法[3]。然而,培养方法可能很难检测出数量较少的特定微生物种属[4],并且由于培养基成分不能满足所有微生物的营养要求,及化学物质敏感度或者选择性不理想,会导致错估微生物的菌群落结构[5]。随着分子生物学分析技术快速发展,高通量测序技术具有较好定量、节约时间、较精准地检测大多数菌种等优点,逐步应用于食品微生物菌群结构分析[6]。采用高通量测序技术分析发现真空包装猪肉中微球菌、肠杆菌、乳酸菌和肉杆菌是主要腐败菌[7]。而高通量测序也存在一定问题,如聚合酶链式反应(polymerase chain reaction,PCR)扩增可能导致错误的相对丰度及产生嵌合序列[8]。因此,在食品复杂体系中的微生物菌群研究中需将培养依赖和培养非依赖的技术相结合。
目前冷鲜肉类的研究主要集中在贮藏品质变化、腐败菌分离鉴定和新保鲜技术研发[9-10],而冷链中牛肉微生物菌群结构及优势菌致腐能力的系统研究仍较少。鉴于此,本研究以牛肉为研究对象,采用依赖培养的16S rDNA测序结合非培养的高通量测序分析牛肉在0 ℃贮藏的微生物菌群变化,鉴定冷鲜牛肉中的优势腐败菌,评价其致腐性。研究为较全面了解冷鲜牛肉中微生物动态变化,分析主要特定腐败菌,以期为靶向抑制腐败菌生长,为高品质牛肉冷鲜技术的研发提供参考。
1 材料与方法
1.1 材料与试剂
牛肉由高沙农贸市场提供,黄牛屠宰后,4 ℃充分冷却排酸72 h,中心温度降至0~4 ℃后取腰大肌(里脊)部位,冰运至实验室,在无菌环境中分割成50 g左右小块,置于塑料托盘中,覆盖聚乙烯膜于(0±1)℃贮藏。本实验微生物分析牛肉样品均为同一批次产品,且牛肉汁制作所用牛肉也为同一批次产品。
平板计数琼脂(plate count agar,PCA)、假单胞菌CFC选择性培养基、热死环丝菌STAA分离培养基、结晶紫中性红胆盐琼脂(violet red bile agar,VRBA)和乳酸菌MRS(De Man, Rogosa and Sharpe)分离培养基、煌绿乳糖胆盐肉汤肉汤(brilliant green lactose bile broth,BGLB) 青岛海博生物技术有限公司;高通量测序细菌基因组DNA提取试剂盒、PCR产物回收试剂盒 美国Omega公司;Quant-iT PicoGreen dsDNA分析试剂盒美国Thermo Fisher Scientific公司;细菌基因组DNA提取试剂盒 日本BioFlux公司;PCR全套试剂 日本TaKaRa公司;引物由生工生物工程(上海)股份有限公司合成;NaCl、干酪素、Na2CO3、H3BO3、甲基红、溴甲酚绿(均为分析纯)、HCl(0.1 mol/L) 上海阿拉丁公司。
1.2 仪器与设备
BXM-75VE型高压灭菌锅、SPX-150B-Z型生化培养箱上海博讯公司;DYY-2c型电泳仪 北京六一仪器厂;QuantiFluor荧光定量系统 美国Promega公司;MiSeq测序仪 美国Illumina公司;3-18K型离心机 德国Sigma公司;UV-1800型紫外分光光度计 日本Shimadzu公司;FE20型实验室pH计 瑞士梅特勒-托利多公司;
KDN-103F型凯氏定氮仪 上海华烨公司。
1.3 方法
1.3.1 牛肉感官评分
参考郑加旭等[11]的感官评分方法,以可接受度作为感官评价标准,牛肉在0 ℃冷藏,每3 d定期取样品,由6 名经过训练的评价员评估其感官评分。0~3 分为好;3~6 分为中;6~9 分为差。
1.3.2 牛肉细菌总数测定
牛肉样品在0 ℃冷藏,每3 d定期取样,参考GB 4789.2—2016《菌落总数测定》[12]测定菌落总数,假单胞菌数、热死环丝菌数、乳酸菌数与菌落总数测定方法相似,分别用CFC琼脂、STAA琼脂、MRS琼脂和PCA计数,其中PCA和CFC琼脂于30 ℃培养48 h,STAA琼脂于22 ℃培养48 h。肠杆菌数参考GB 4789.3—2016《大肠菌群计数》[13]测定,用VRBA琼脂计数。
1.3.3 细菌分离和16S rDNA鉴定
将冷鲜牛肉在贮藏初期0 d和末期2 1 d样品培养后,挑取P C A上外观形态不同的菌落1 3 株和1 5 株,纯化后,挑取单菌落,进行革兰氏染色,并提取基因组D N A,用通用引物[14]1 6 S F(5’-AGAGTTTGATCCTGGCTCAG-3’)和16S R(5’-AAGGAGGTGATCCAGCCGCA-3’)扩增16S rDNA片段,测序及拼接序列后,在NCBI网站(https://www.ncbi.nlm.nih.gov/)的BLAST程序进行核酸序列的同源性比对。
1.3.4 高通量测序
取采集的牛肉样品各5 g,加入45 mL无菌生理盐水,4 ℃条件下摇床中振荡2 h,4 ℃、200×g离心2 min,取上清液12 000×g再次离心2 min,弃上清液,收集沉淀。
DNA提取:应用OMEGA公司的DNA提取试剂盒提取上述沉淀中的细菌基因组DNA,用紫外分光光度计检测DNA浓度和纯度。
PCR扩增及高通量测序:采用通用引物[15]515F(5’-GTGCCAGCMGCCGCGGTAA-3’)和806R(5’-GGACTACHVGGGTWTCTAAT-3’)PCR扩增细菌16S rDNA。反应条件均为98 ℃预变性30 s;98 ℃变性10 s,54 ℃退火30 s,72 ℃延伸45 s,35 个循环;72 ℃延伸10 min。对纯化后的PCR产物采用Quant-iT PicoGreen dsDNA试剂盒在QuantiFluor荧光定量系统上对基因文库进行定量;使用MiSeq测序仪进行双端测序。
数据分析处理,将测序获得的双端数据拼接成长的tag,并将序列上建库引入的barcode和引物序列去除。此外,按照荧光信号强度修剪质量值低的序列。利用Vsearch(https://github.com/torognes/vsearch)对序列进行聚类,通过操作分类单元(operational taxonomic unit,OTU)的Alpha多样性分析得到样品中OTU数目(丰富度)和群落结构的稳定性(均匀度)[16]。通过OTU稀释曲线分析,评估测序量是否达标。并与RDP数据库和NT-16S数据库进行相似性比对,选取丰度大于0.5%的物种分类,将其余的物种设置为others,以BIOM格式OTU表统计OTU丰度[17]。
1.3.5 无菌牛肉汁的制备和接种
无菌牛肉汁提取参考刘永吉[18]的方法,略作修改。具体如下:将100 g新鲜牛肉切碎后,用200 mL无菌水拍打成匀浆液,4 ℃、5 000×g离心15 min,将上清液过0.22 μm无菌滤膜后备用。将贮藏末期的15 株分离株过夜培养物,用生理盐水稀释至106~107CFU/mL,以1%接种到无菌牛肉汁,使初始浓度保持在104~105CFU/mL。牛肉汁在0 ℃冷藏,在第6、15天取样进行感官、pH值和挥发性盐基氮(total volatile basic nitrogen,TVB-N)值检测,以接种无菌水的样品作空白对照。
1.3.6 牛肉汁中感官和蛋白酶测定
取牛肉汁样品5 mL,感官评价参考1.3.1节方法。并取牛肉汁1.5 mL在4 ℃、10 000×g离心10 min,上清液0.22 μm无菌滤膜后,参照SB/T 10317—1999《蛋白酶活力测定法》[19]中的福林-酚方法测定蛋白酶活力。
1.3.7 牛肉汁中pH值和TVB-N值测定
牛肉汁混匀,离心,上清液0.22 μm无菌滤膜后,参照pH计说明书和GB 5009.228—2016《食品中挥发性盐基氮的测定》[20]半微量定氮法测定pH值和TVB-N值。
1.4 数据处理与统计分析
牛肉冷藏中微生物鉴定实验和牛肉汁评价腐败分离株致腐性均设计3 个重复,采用Microsoft Excel 2010和Origin 9.0软件(OriginLab公司)进行数据处理和作图,并利用SPSS 19.0软件(IBM公司)的AVOVA进行方差分析,P<0.05,差异显著,结果为。
2 结果与分析
2.1 冷鲜牛肉感官评分腐败菌总数变化
如图1所示,牛肉在冷藏前9 d感官品质保持较好,第12天肉品新鲜度下降,第15天出现异味,第18天样品有酸败味,感官品质不可接受。冷鲜牛肉初始(0 d)菌落总数为4.06(lg(CFU/g)),表明样品品质良好。牛肉在0 ℃贮藏初期微生物缓慢增加,第3天增长快速,至第9天超过6(lg(CFU/g)),而第15天达到8.73(lg(CFU/g)),之后达到稳定。牛肉在冷藏中CFC琼脂上假单胞菌数、STAA琼脂上热死环丝菌数、VRBA琼脂上肠杆菌数和MRS琼脂上乳酸菌数不断上升,其增长趋势与菌落总数相似,其中假单胞菌数和热死环丝菌数增长最快,肠杆菌数和乳酸菌数的增长较慢。牛肉贮藏末期假单胞菌数最高,热死环丝菌次之,而肠杆菌数和乳酸菌数低于假单胞菌约2(lg(CFU/g))(P<0.05)。结合感官评价和菌落总数,0 ℃冷藏牛肉的货架期约为15 d。
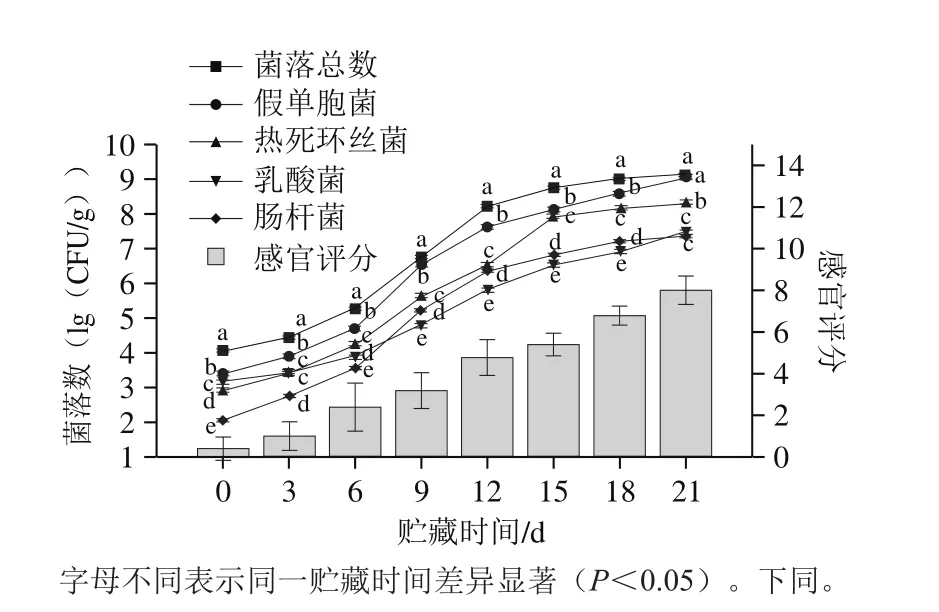
图1 牛肉0 ℃贮藏时感官评分和菌落数的变化Fig. 1 Changes in sensory score and microbial load of beef stored at 0 ℃
2.2 冷鲜牛肉菌相的16S rDNA鉴定
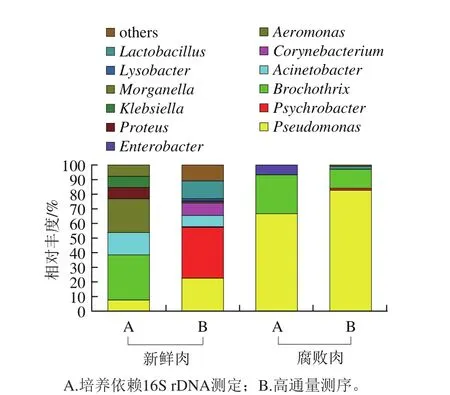
图2 冷鲜牛肉新鲜肉和腐败肉中细菌群落结构组成的差异Fig. 2 Bacterial community structures of all samples at the genus level
从新鲜牛肉和腐败牛肉样品分别挑选具有特异性形态的菌落13 株和15 株,经纯化及初步鉴定后发现新鲜肉和腐败肉分别有9 株和11 株为革兰氏阴性菌,表明牛肉中腐败微生物菌群主要以革兰氏阴性菌为主。将28 株菌以16S rDNA 通用引物PCR扩增,测序后得到的序列采用GenBank数据库中的BLAST进行同源性比对显示,新鲜肉分离株中有4 株与热死环丝菌有较高的同源性,约占30.77%;3 株与气单胞菌属(Aeromonasspp.)有较高的同源性,占23.08%,有2 株与不动杆菌属(Acinetobacterspp.)有较高的同源性,约占15.38%,还有4 株分别与假单胞菌属(Pseudomonasspp.)、奇异变形杆菌(Proteus mirabilis)、克雷伯氏菌属(Klebsiellaspp.)和摩根氏菌属(Morganellaspp.)同源性较高,各占7.69%(图2)。而腐败肉有10 株与假单胞菌有较高的同源性,约占66.67%,4 株与热死环丝菌同源性较高,约占26.67%,还有1 株与蜂房哈夫尼菌(Hafnia alvei)同源性较高,占6.67%。培养及生理生化特征结果显示好氧革兰阴性菌假单胞菌和阳性菌热死环丝菌是冷鲜牛肉有氧贮藏下的优势腐败菌[21],Pennacchia等[22]对冷鲜牛肉可培养微生物群的高通量测序,发现贮藏20 d后假单胞菌等为优势腐败菌,还有某些嗜冷肠杆菌,如格氏沙雷氏菌(Serratia grimesii)等。
2.3 冷鲜牛肉微生物高通量检测分析
采用高通量测序技术进一步分析冷鲜牛肉初始和腐败样品菌群分布。结果发现104 个属的细菌和部分未分类的细菌被挖掘,其中新鲜牛肉中有89 种,腐败牛肉中有43 种,其中两个样品中共有OTU数为27。选取其中相对丰度大于0.5%的7 个物种分类,绘制样品丰度堆叠柱状图。如图2所示,冷鲜牛肉贮藏前期的优势菌为嗜冷杆菌属(Psychrobacterspp.,34.78%)、假单胞菌属(22.64%)、乳杆菌属(Lactobacillusspp.,12.21%)、棒状杆菌属(Corynebacteriumspp.,8.44%)、不动杆菌属(7.76%)。而货架末期的优势菌为假单胞菌属(82.78%)、环丝菌属(13.11%)、乳杆菌属(1.90%)、嗜冷杆菌属(1.27%)。Ercolini等[23]使用高通量测序技术分析发现,牛肉低温有氧和高氧贮藏下假单胞菌是主要腐败菌。Ercolini等[24]运用该技术分析显示牛肉在低温普通和气调包装初期微生物主要为假单胞菌和热死环丝菌,而贮藏后期普通包装假单胞菌优势生长,气调包装乳酸菌占优势。
上述结果发现,培养依赖16S rDNA结合不依赖培养高通量测序分析菌群种类基本相似,冷鲜牛肉初始菌相复杂,主要由热死环丝菌、嗜冷菌、假单胞菌、不动杆菌、气单胞菌等多种微生物构成。随着冷藏期的延长牛肉中腐败菌相趋于简单,其中假单胞菌和热死环丝菌为优势腐败菌,占主导地位。然而两种方法在鉴定部分微生物种属中仍存在一定偏差,特别是新鲜肉样。通过培养依赖16S rDNA方法在新鲜牛肉未鉴定到嗜冷杆菌、乳杆菌、棒状杆菌和溶杆菌,与PCA营养成分和培养条件的限制,导致这些菌不能生长有关。而培养依赖方法中发现的变形杆菌、克雷伯氏菌和摩根氏菌未能在高通量测序中找到,可能因其相对丰度过低,未被扩增。
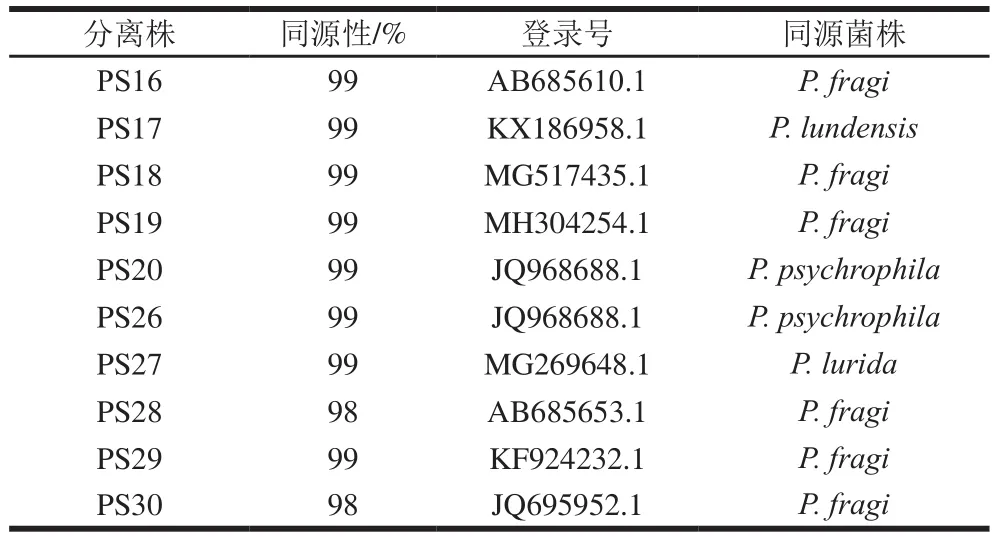
表1 腐败肉假单胞菌分离株16S rDNA序列同源性分析Table 1 Homology analysis of 16S rDNA genes of Pseudomonas isolates
研究发现腐败牛肉中假单胞菌是优势腐败菌,如表1所示。同源性发现致腐菌假单胞菌中PS28、PS16、PS30、PS18、PS29、PS19六株与莓实假单胞菌(P. fragi)有较高同源性,PS26和PS20与嗜冷假单胞菌(P. psychrophila)有较高同源性,另外PS17与隆德假单胞菌(P. lundensis)、PS27与拉丁假单胞菌(P. lurida)有较高同源性。Casaburi等[25]相似地报道莓实假单胞菌是肉类中最常见假单胞菌,其次是隆德假单胞菌和荧光假单胞菌。Ercolini等[26]发现假单胞菌是有氧保藏冷鲜牛肉中的优势腐败菌,所有莓实假单胞菌均能产生酯类挥发性物质,对肉类腐败起到重要作用。可见,莓实假单胞菌是冷鲜牛肉的优势腐败菌。
2.4 优势腐败菌在牛肉汁中致腐性评价
2.4.1 细菌生长
无菌牛肉汁可以提供牛肉中可作为能量和营养来源的物质,为一种近似牛肉的营养介质。细菌生长是细菌腐败能力显现的重要方面,在无菌牛肉汁中评价其生长特性,可以更客观地模拟腐败菌在牛肉中生长繁殖。牛肉汁空白组在贮藏期内无细菌生长。如图3所示,在货架期前期(6 d),接种热死环丝菌组、假单胞菌组和蜂房哈夫尼菌组分别达到7.23~7.33、6.80~7.10(lg(CFU/mL))和6.81(lg(CFU/mL))。在货架期末期(15 d),接种牛肉汁的菌液浓度热死环丝菌组达到8.43~8.50(lg(CFU/mL)),假单胞菌组为7.58~7.95(lg(CFU/mL)),而蜂房哈夫尼菌的菌液浓度达到8.46(lg(CFU/mL))。在3 种优势腐败菌中热死环丝菌和哈夫尼细菌菌液浓度较高,假单胞菌菌液浓度较低。
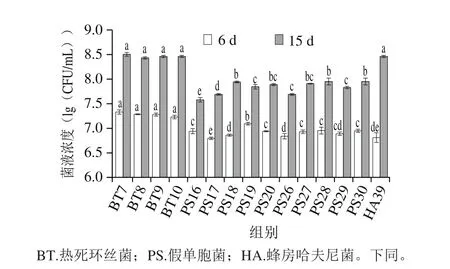
图3 热死环丝菌、假单胞菌和哈夫尼菌在0 ℃牛肉汁中的菌体生长Fig. 3 Growth rates of Brochothrix, Pseudomonas and Hafnia inoculated in sterilized beef juice stored at 0 ℃
2.4.2 感官评定
感官评定是评价肉制品品质的重要指标,因微生物生长繁殖过程中利用肉类蛋白质等营养物质,逐步积累代谢降解产物,导致牛肉汁出现浑浊、散发酸臭味和硫化氢味等腐败变质现象。如图4所示,牛肉汁空白组感官评价良好。而接菌组感官品质均呈下降趋势,较空白组感官变化明显。冷藏6 d接种热死环丝菌组、假单胞菌组和蜂房哈夫尼菌组的感官评分分别达到1.33~2.17、2.67~6.33和4.00。15 d接种牛肉汁的感官评分热死环丝菌组达到3.17~4.50;假单胞菌组为6.00~8.33,不可接受;而蜂房哈夫尼菌的感官评分达到8.33,也不可接受。在3 种优势腐败菌中假单胞菌和哈夫尼细菌感官变化较快,其中莓实假单胞菌PS28最高,其次为PS30、PS26和PS27。假单胞菌和肠杆菌科细菌代谢利用氨基酸,释放NH3等刺激性气体,导致感官快速劣变[27]。而热死环丝菌感官变化较小,可能与该菌在有氧条件下利用葡萄糖生成乙偶姻及乙酸,产生甜的气味[28]有关。

图4 热死环丝菌、假单胞菌和哈夫尼菌在0 ℃牛肉汁中的感官评分Fig. 4 Sensory scores of sterilized beef juice inoculated with Brochothrix, Pseudomonas and Hafnia during storage at 0 ℃
2.4.3 蛋白酶活性
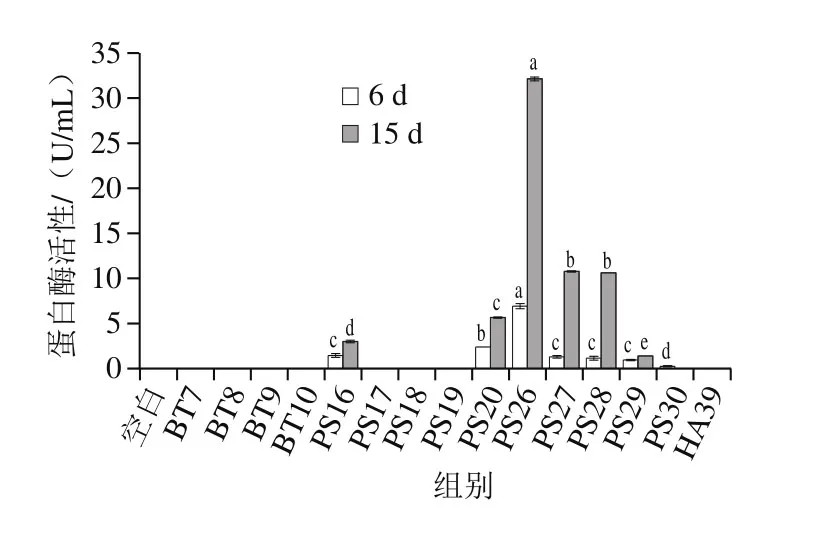
图5 热死环丝菌、假单胞菌和哈夫尼菌在0 ℃牛肉汁中的蛋白酶活性变化Fig. 5 Changes in protease activity of Brochothrix, Pseudomonas and Hafnia inoculated in sterilized beef juice during storage at 0 ℃
食品腐败菌能够分泌蛋白酶,分解动物性产品中的蛋白质,促进微生物大量增殖,导致肉类腐败。如图5所示,热死环丝菌、假单胞菌和蜂房哈夫尼菌优势腐败菌蛋白酶活性在第6天变化较小,仅发现假单胞菌PS26、PS20、PS16、PS27、PS28、PS29和PS30表现蛋白酶活性(P<0.05),除PS30外在冷藏至第15天显著增加(P<0.05),分别达到32.13、5.66、3.00、10.77、10.62、1.40 U/mL,其中莓实假单胞菌PS26最高,其次为PS27和PS28。假单胞菌的蛋白酶活性具有菌株依赖[29],而热死环丝菌和蜂房哈夫尼菌未有蛋白酶活性。
2.4.4 pH值

图6 热死环丝菌、假单胞菌和哈夫尼菌在0 ℃牛肉汁中的pH值变化Fig. 6 Changes in pH value of sterilized beef juice inoculated with Brochothrix, Pseudomonas and Hafnia during storage at 0 ℃
如图6所示,牛肉汁空白组初期pH值为6.10,冷藏15 d无变化(P>0.05)。热死环丝菌组冷藏6 d pH值下降至5.09~5.41(P<0.05),假单胞菌组pH值上升至6.08~6.85(P<0.05),蜂房哈夫尼菌组pH值达到6.23。贮藏至第15天,热死环丝菌组中除BT7下降外其余无明显降低(P>0.05),假单胞菌组中PS16、PS17、PS19、PS26、PS27菌pH值显著上升(P<0.05),增至6.28~6.82,其余组无显著变化(P>0.05);接种蜂房哈夫尼菌牛肉汁pH值为6.27。研究发现接种热死环丝菌BT7和BT10的pH值最低,而接种假单胞菌PS28的pH值最高,达到7.18。冷藏牛肉汁的pH值变化与微生物代谢产物的积累有关,研究表明热死环丝菌在有氧条件下以缬氨酸和亮氨酸为底物产生挥发性酸,导致牛肉汁pH值降低[28],而假单胞菌和肠杆菌科细菌代谢利用蛋白质,释放氨等碱性物质,导致肉类pH值上升[30]。
2.4.5 TVB-N值

图7 热死环丝菌、假单胞菌和哈夫尼菌在0 ℃牛肉汁中的TVB-N值变化Fig. 7 Changes in TVB-N value of sterilized beef juice inoculated with Brochothrix, Pseudomonas and Hafnia during storage at 0 ℃
牛肉冷藏期间TVB-N的累积是由于在微生物和酶的作用下,引起蛋白质的脱氨、脱羧作用,降解产生氨及胺类等挥发性的碱性含氮物质,是评价肉质新鲜度最重要的指标。从图7可知,无菌牛肉汁初期TVB-N值为5.25 mg/100 mL。接种热死环丝菌组、假单胞菌组和蜂房哈夫尼菌组在冷藏至第6天时,TVB-N值分别为6.30~7.00、5.60~10.85 mg/100 mL和7.00 mg/100 mL。冷藏15 d热死环丝菌组中TVB-N值上升至8.23~8.58 mg/100 mL,假单胞菌组中除PS16和PS30外,其余接种牛肉汁TVB-N值上升至8.23~15.23 mg/100 mL(P<0.05),其中莓实假单胞菌PS28菌株上升最快(P<0.05);而接种蜂房哈夫尼菌牛肉汁的TVB-N值达到8.23 mg/100 mL。
腐败末期3 种优势腐败菌均能在冷藏条件下缓慢生长,且表现不同的致腐性,假单胞菌和蜂房哈夫尼菌感官评分、pH值和TVB-N值高于热死环丝菌,提示这两种致腐菌能形成氨类代谢产物。并且假单胞菌表现较强的蛋白酶活性。从鸡肉中分离出致腐性沙雷氏菌、假单胞菌和杀鲑气单胞菌,也显示假单胞菌组TVB-N值、pH值均高于其他分离菌,且异味和黏液分泌明显[31]。而接种的热死环丝菌对真空包装牛肉感官影响较小[32]。Gribble等[33]对接种热死环丝菌和肠杆菌的真空包装羊肉的颜色、气味以及产气能力评价显示肠杆菌致腐性更强,更早形成腐败。在多株牛肉源的假单胞菌分离株中莓实假单胞菌PS28致腐性最强,相似地报道莓实假单胞菌形成酯、酮、醇、硫化合物等多种不良的风味物质[25]。
3 结 论
研究采用培养依赖16S rDNA鉴定结合高通量测序技术较详实地分析冷鲜牛肉冷藏中微生物群落结构的动态变化,发现冷鲜牛肉贮藏初期细菌表现多样性,而腐败终点菌群种类明显下降,以假单胞菌和热死环丝菌为优势腐败菌。腐败末期优势腐败菌表现不同的致腐性,其中假单胞菌和蜂房哈夫尼菌致腐性强于热死环丝菌,假单胞菌还表现较强的蛋白酶活性,其中莓实假单胞菌PS28致腐性最强。可见,冷鲜牛肉中的特定腐败菌是莓实假单胞菌。
