两种放牧模式下高寒草甸群落植被构成及稳定性特征
2019-09-24施颖胡廷花高红娟罗巧玉于应文
施颖,胡廷花,高红娟,罗巧玉,于应文*
(1.兰州大学草地农业生态系统国家重点实验室,兰州大学农业农村部草牧业创新重点实验室,兰州大学草地农业科技学院,甘肃 兰州 730020;2.青海师范大学生命科学学院,青海 西宁810000)
高寒草甸作为青藏高原典型植被类型,约占该区域草地面积的1/2[1],在青藏高原生物多样性保护、生态系统维持、畜牧业发展等方面发挥重要作用[2]。近年来,受全球气候变暖和人类活动影响以及不合理草地利用[3],导致该区域草地退化严重。其中,不合理放牧是导致草地退化的主要人为因素之一[4]。通常,青藏高原高寒草甸主要分为冬春放牧的冷季牧场、夏季放牧的夏季牧场及牧户周边全年放牧的四季牧场[5-7];夏季牧场由于面积广阔,放牧时期短,且植物处于生长季,而利于草地群落物种多样性和均匀度的维持[8];冷季牧场因牧户家畜数量和管理措施及水平不同,其放牧强度和草地质量差异较大,四季牧场因多为公共放牧地而过牧,从而青藏高原高寒草甸生态系统管理的关键是草地退化主要发生的冷季牧场和四季牧场。研究表明,合理的放牧管理,不仅利于草地群落结构改善,较高草地生产力及群落物种多样性的维持[9],还利于草地养分循环及土壤结构改变,进而促进草地的进展演替,提高草地的稳定性。因此,研究冷季牧场和四季牧场的草地稳定性,探讨适宜的草地放牧模式,对于青藏高原高寒草甸的健康管理和生产力维持具有重要意义。以往研究也表明,连续放牧使土壤养分流失严重[7],休牧能提高草地土壤养分含量及根系生产力,恢复草原基况[10-11];围封可维持草原较高生产力,春季或生长季放牧比冷季放牧对草地生产力影响大[8,12]。虽然这些研究从放牧强度、封育、施肥、补播及混合措施[13-16]等角度,系统分析了青藏高原高寒草甸的植物群落学[17-18]和土壤理化特征[7,19-21],为草地稳定性的深入研究提供了理论基础,但相对缺乏季节性放牧下高寒草甸稳定性的探讨,也缺乏青藏高原最为普遍的特定放牧强度下冷季放牧和四季放牧高寒草甸稳定性的比较研究。本研究通过对青藏高原东北缘冷季放牧和四季放牧模式下,高寒草甸群落植被构成和土壤养分特征及群落稳定性分析,明晰高寒草甸的群落稳定性特征,探究适宜的草地管理模式,为高寒草甸的合理放牧管理提供实践依据。
1 材料与方法
1.1 研究区概况
研究区位于青藏高原东北缘的甘肃省天祝县抓喜秀龙乡境内的甘肃农业大学天祝高山草原试验站附近的高寒草甸上,地理坐标为E 102°40′-102°47′,N 37°11′-37°14′,海拔2960 m。年均温-0.1 ℃,1月气温最低为-18.3 ℃,7月气温最高为12.7 ℃,>0 ℃积温1380 ℃;多年平均降水量416 mm,无绝对无霜期,仅分冷热两季,土壤类型为亚高山草甸土。草地植物种主要有垂穗披碱草(Elymusnutans)、矮嵩草(Kobresiahumilis)、线叶嵩草(Kobresiacapillifolia)、洽草(Koeleriacristata)、针茅(Stipakrylovii)、珠芽蓼(Polygonumviviparum)、阴山扁蓿豆(Melilotoidsruthenicavar.inschanica)和球花蒿(Artemisiasmithii)等。
1.2 样地设置
2016年5月,在研究区草地利用最为普遍的冷季放牧(cold season grazing,CSG)和四季放牧(warm and cold season grazing,WCSG)牧场,分别设置面积约0.3~0.5 hm2的样地各3块,作为3次样地重复。其中,CSG牧场的草地植被构成、放牧史及草地利用率在1999年之前与WCSG一致。但自1999年,CSG牧场改为冷季牧场后,CSG牧场仅在冷季,即每年11月-翌年5月放牧,其余时间均处于围栏休牧状态,而WCSG 牧场仍在暖季(6-10月)和冷季(11月-翌年5月)进行连续放牧。试验之前和试验期间,CSG牧场和WCSG牧场的放牧家畜种类(牦牛和臧羊)一样。1999年之后,CSG牧场的冷季草地利用率约为85%~95%,WCSG牧场的暖季和冷季利用率分别约为55%~65%和85%~95%;1999-2016年期间,CSG牧场实际在冷季进行高强度放牧,使CSG牧场在翌年4月末的草地牧草现存量基本与WCSG牧场一样,进而试验期间,CSG牧场和WCSG牧场的全年放牧率相近[22]。
1.3 测定和分析方法
1.3.1植物群落特征 2016年7月下旬至8月中旬,在各设置重复样地内,随机设置0.5 m×0.5 m的样方10个,每个牧场共设置30个样方,调查各样地内所有植物种的高度、盖度、地上生物量及群落盖度。地上生物量测定时,先按绿色物质和死物质分开,再将绿色物质按不同植物种分开,然后置入烘箱65 ℃下烘干称重。基于样方植物种高度、盖度、生物量及物种数数据,计算植物种重要值、功能群生物量构成、草地Raunkiaer频度系数及演替度。
植物种重要值(importance value,IV):按公式IV=(RC+RH)/2计算。式中,RH和RC分别为植物种的相对高度(relative height,RH=样方内某植物种的高度/样方内所有植物种的高度之和)和相对盖度(relative coverage,RC=样方内某植物种的盖度/样方内所有植物种的盖度之和)。
功能群生物量构成:将样方分种生物量按经济类群分为禾草、莎草、豆科和其他4类;按生活型分为地面芽(含地上芽)植物(hemicryptophyte,HP)、地下芽(geophyte,GP)植物和一年生(含越年生)植物(therophyte,TP)3类[22]。
草地Raunkiaer频度系数(R):按公式R=(n/N)×100%计算,式中,n为某一个种在全部取样中出现的次数;N为全部取样数。Raunkiaer把频度系数划分为5个等级,A(级)为1%~20%,B(级)为21%~40%,C(级)为41%~60%,D(级)为61%~80%,E(级)为81%~100%。

1.3.2土样采集分析 在测定完植被特征的各样方内,同期用直径3.5 cm土钻采集0~10 cm和10~20 cm土样,将同类放牧草地上相同重复样地的10钻同层土样合为1个混合样,共采集12个混合土样,分别测定有机碳(organic carbon,C)(重铬酸钾-浓硫酸油浴法)、全氮(total nitrogen,N)(凯氏定氮法)、全磷(total phosphorus,P)(钼锑抗比色法)及全钾(total potassium,K)含量(火焰光度法)[24]。
1.4 数据分析
用Excel进行数据处理及制图,SPSS 16.0软件的One-Way ANOVA对两种放牧模式下草地群落特征(高度、盖度、生物量及物种数)、功能群生物量构成及土壤养分进行差异显著性分析,并将数据表示为均值(mean)±标准误(standard error of mean,SEM)。
2 结果与分析
2.1 植物种重要值
草地主要植物种重要值显示(表1), CSG草地优势和亚优势种以冷地早熟禾、 垂穗披碱草、 无芒雀麦和阴山扁蓿豆, 其重要值分别为0.295、0.155、0.150和0.136; 矮嵩草、线叶嵩草、高山紫菀、大籽蒿、甘肃棘豆、多裂委陵菜、车前、蒲公英、甘肃马先蒿为主要伴生种。WCSG草地以矮嵩草和垂穗披碱草为优势种,其重要值分别为0.183和0.151;线叶嵩草、阴山扁蓿豆和早熟禾为亚优势种,其重要值分别为0.111、0.101和0.100;紫花针茅、球花蒿、蒲公英、高山紫菀、二裂委陵菜为主要伴生种。说明,CSG增加少数杂类草的环境适应性,WCSG草地由矮嵩草/垂穗披碱草群落向禾草/杂类草群落转化。

表1 两种放牧模式下高寒草甸主要植物种重要值Table 1 The importance values (IV) of main species under two grazing modes
CSG:冷季放牧Cold season grazing;WCSG:四季放牧Warm and cold season grazing.下同The same below.
2.2 群落特征
草层高度、盖度和总地上生物量均为CSG草地极显著高于WCSG草地(P<0.001),且前者分别约为后者的6.5、1.1和2.8倍;群落植物物种数为CSG草地显著低于WCSG草地(P<0.05),后者是前者1.3倍(表2)。说明CSG管理利于草地植物生长,降低草地群落物种数。
2.3 功能群生物量构成
草地群落植物种经济类群生物量构成中,CSG草地中禾草比例占主体(86.5%),WCSG草地莎草比例占主体(59.1%),此与其重要值构成结果一致;且两种放牧模式下豆科和其他科植物的生物量比例相近(P>0.05)(图1)。植物种生活型生物量构成为,CSG草地中地面芽比例显著高于WCSG草地(P<0.001),前者为后者的1.4倍;但前者的地下芽植物和一年生植物比例均低于后者(P<0.01或P<0.05),且前者分别为后者的18.2%和6.8%(图1)。说明,CSG管理利于草地群落中禾草和地面芽植物的生长,WCSG管理有利于莎草、地下芽及适口性差的一年生植物的生长。

表2 两种放牧模式下高寒草甸植物群落特征Table 2 Plant community characteristics under two grazing modes
注:*, ***分别表示P<0.05和P<0.001水平差异显著。
Note: *, *** mean significant differences at 0.05, 0.001 level, respectively.

图1 两种放牧模式下高寒草甸群落功能群生物量构成Fig.1 Biomass composition under two grazing modes HP:地面芽Hemicryptophyte; GP:地下芽Geophyte; TP:一年生Therophyte. ns:*,**, ***分别表示 P>0.05水平差异不显著,P<0.05,P<0.01 和P<0.001水平差异显著。ns:*, **, *** mean not significant differences at 0.05, and significant differences at 0.05, 0.01 and 0.001 level, respectively.
2.4 物种Raunkiaer频度系数和演替度
物种Raunkiaer标准频度直方图显示,CSG草地的各级植物种频度构成呈典型反“J”型,即A级(64%)>B级(16%)>C级=D级(4%)
2.5 土壤养分
放牧模式对草地土壤C、N、C/N、C/P、C/K、N/P和N/K含量有显著影响(P<0.05、P<0.01或P<0.001),土壤深度对其C、N、C/K含量影响显著(P<0.05),放牧模式和土壤深度互作对其C、C/N、C/K含量影响显著(P<0.05);放牧模式、土壤深度及其互作对其P、K、P/K含量均无显著影响(P>0.05)。说明,CSG管理利于草地土壤有机C和全N养分恢复和维持。
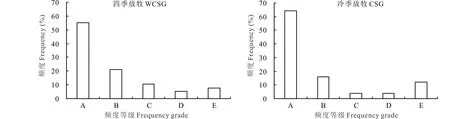
图2 两种放牧模式下高寒草甸物种Raunkiaer的标准频度直方图Fig.2 Standard frequency histogram under two grazing modes A: 1%~20%; B: 21%~40%; C: 41%~60%; D: 61%~80%; E: 81%~100%.
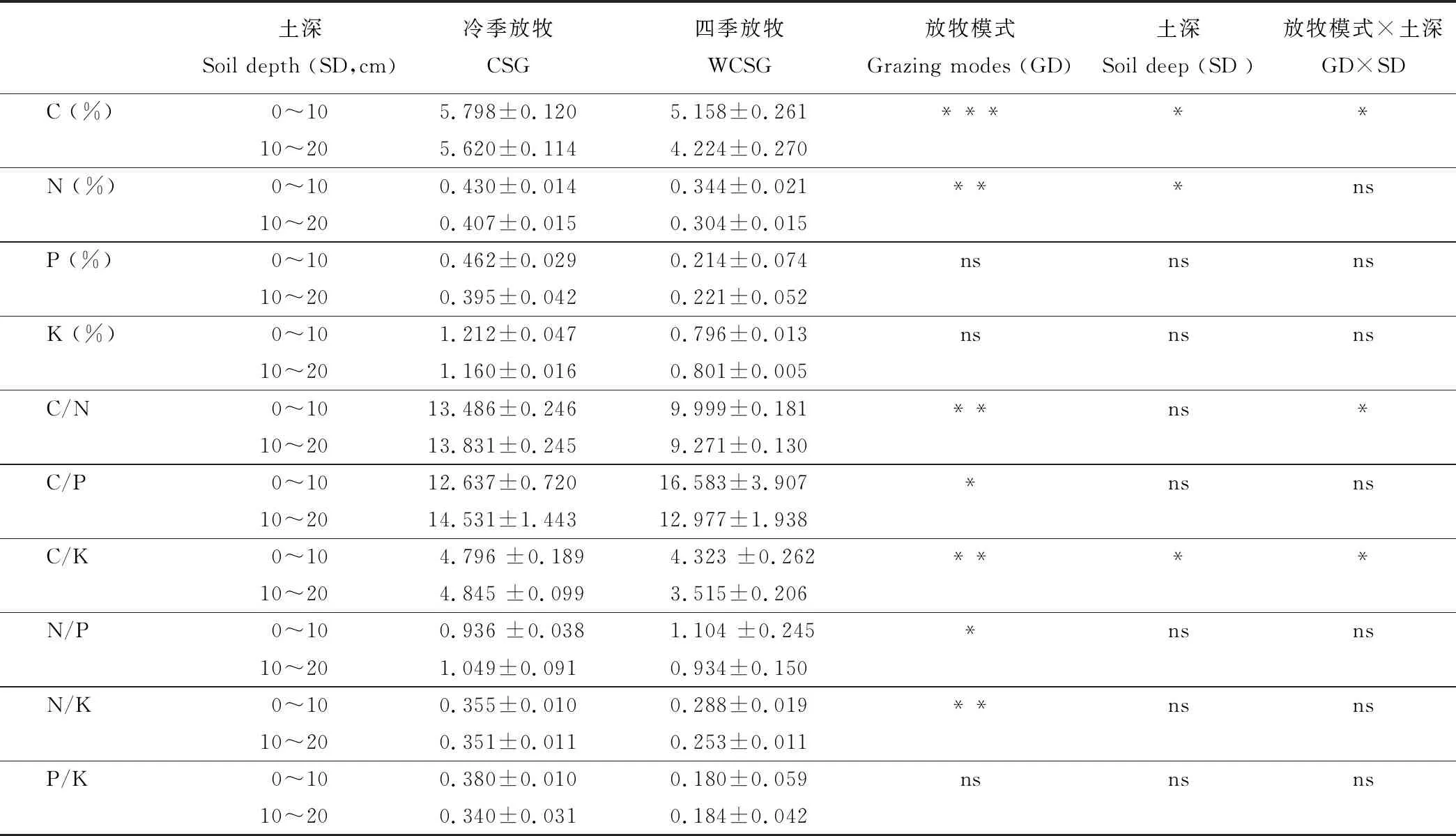
土深Soil depth (SD,cm)冷季放牧CSG四季放牧WCSG放牧模式Grazing modes (GD)土深 Soil deep (SD )放牧模式×土深 GD×SDC (%)0~105.798±0.1205.158±0.261*****10~205.620±0.1144.224±0.270N (%)0~100.430±0.0140.344±0.021***ns10~200.407±0.0150.304±0.015P (%)0~100.462±0.0290.214±0.074nsnsns10~200.395±0.0420.221±0.052K (%)0~101.212±0.0470.796±0.013nsnsns10~201.160±0.0160.801±0.005C/N0~1013.486±0.2469.999±0.181**ns*10~2013.831±0.2459.271±0.130C/P0~1012.637±0.72016.583±3.907 *nsns10~2014.531±1.44312.977±1.938 C/K0~104.796 ±0.1894.323 ±0.262****10~204.845 ±0.0993.515±0.206 N/P0~100.936 ±0.0381.104 ±0.245*nsns10~201.049±0.091 0.934±0.150 N/K0~100.355±0.010 0.288±0.019 **nsns10~200.351±0.011 0.253±0.011 P/K0~100.380±0.010 0.180±0.059 nsnsns10~200.340±0.031 0.184±0.042
注:ns, *, **, ***分别表示P>0.05水平差异不显著,P<0.05,P<0.01 和P<0.001水平差异显著。
Note: ns, *, **, *** mean not significant differences at 0.05, and significant differences at 0.05, 0.01 and 0.001 level, respectively.
3 讨论与结论
植物群落特征变化主要体现在物种组成及优势种和亚优势种更替方面[25-26]。本研究冷季牧场禾草占绝对优势,四季牧场莎草占优势。其主要原因为冷季牧场因生长季不放牧,使植株高、根系较深的禾草更具光和养分竞争优势[18],从而抑制草丛中矮生型杂类草和莎草的生长;且由于禾草的营养价值和适口性较高,家畜喜食,使四季牧场中处于上层的禾草因家畜的频繁采食,而抑制了禾草的补偿性生长[27],进而降低禾草的生长优势及其与莎草的竞争,由此利于低矮莎草生长,增加草地中莎草比例。同时,冷季牧场中高大禾草的生长和竞争优势,也抑制草层下部低矮不耐阴植物种生长,使其逐渐从群落中消失[28],由此降低冷季牧场的植物物种数;此与孙涛等[29]和刘玉等[8]报道的结果一致,亦与李佳琪等[18]所得的“去除物种”而为适应干扰的先锋物种释放生态的结果类似。本研究四季放牧草地以矮嵩草、垂穗披碱草、线叶嵩草、阴山扁蓿豆为主要植物种,无绝对优势种,导致其群落物种间竞争大,也证实草地植物种间竞争主要发生在不同科属种之间[30]。此外,本研究冷季放牧草地中莎草比例很低,这不仅与生长季较高禾草的生长优势抑制了草层中较矮嵩草植物的生长有关,也与生物量测定时,草地中莎草科主要植物嵩草属因生长低矮而仅收获一部分有关。
草地植物群落特征(特别是植物物种种间关系和功能群特征)及植物物种多样性是草地稳定性和演替度的主要评价指标[31]。物种功能群构成集中反映植物对生境的适应性[27]。本研究冷季放牧促进草地地面芽植物生长,抑制地下芽和一年生植物生长,其原因一方面是,冷季牧场因家畜采食大量地上枯落物[8],为返青期植物的生长提供充足光照和空间资源而利于植物的再生和幼苗形成,且生长季无家畜啃食和践踏,这利于地面芽和地上芽植物的生长;另一方面,由于植物根系的生长相对地上具有明显滞后特性[32],而生长季因禾草强大须根系,使其与对干扰极敏感的地下芽植物形成强烈竞争[18,33],进而抑制地下芽生长。本研究四季放牧草地的物种多样性较高,其主要原因是家畜的持续采食,降低草地中优势种的优势度和生态位宽度,为毒杂草和适口性差的一年生植物的生长创造条件,进而使草地物种多样性增加。这与魏斌等[30]的研究中封育使地面芽植物物种数增加的结果相同,使一年生植物增加和地下芽植物差异不明显的结果存在差异,可能与草地退化程度和封育时期不同等有关。
土壤养分恢复是评价生态系统恢复的一个重要因素[34],本研究冷季放牧利于草地土壤C和N养分的恢复和保持。其原因是四季牧场全年处于放牧状态,草地地上生物量和凋落物量持续处于较低状态,进而通过植物残体等输入土壤的养分量较少;冷季放牧草地因牧草生长季家畜未放牧而利于牧草的充分生长和群落结构优化,也使凋落物等进入草地土壤的养分量增加[20],进而改善草地土壤环境所致。本研究两种放牧模式的草地土壤全P含量无差异,这与曹建军等[35]的研究结果一致,其原因是草地土壤全P含量自身较稳定[35]。本研究四季放牧草地因家畜连续放牧所沉积排泄物量较多,草地内土壤养分的空间异质性较大,从而土壤全P含量的标准误很大,由此使两种放牧模式草地的土壤全P含量差异消除。
群落稳定性取决于物种对生境变化的敏感性和种间关系对环境干扰的放大效应[18]。本研究基于草地植物物种的Raunkiaer标准频度及植被构成和土壤养分特征结果,即冷季牧场群落呈典型反“J”型,其群落趋于相对成熟且稳定阶段,且冷季牧场具较高的禾草组分、较低的莎草组分及较高的土壤C、N含量;得知,冷季放牧促进高寒草甸由莎草/杂类草群落向禾草/杂类草群落演替,提高草地稳定性的结论。可见,适宜放牧制度是草地植物生长季放牧的关键,也是影响草地稳定性的主导因子。本研究中得出的冷季放牧群落多样性虽没有四季放牧高,却处于相对稳定阶段,这与王长庭等[36]得出的一个生态系统中多样性与生态系统稳定性有正相关趋势,但在种群水平上的稳定性不一定适应的结果相符;亦与魏斌等[30]得出演替度高的草地,其物种多样性反而低的结论一致。同时,由于群落演替与发展变化需要空间梯度和时间梯度,虽然本研究冷季放牧草地的演替度值具有更大趋势,但其与四季放牧草地无差异。这是由于演替是一个长期渐变过程,可能因两种放牧模式之间的差异效应经7年的处理还没充分展现出来,也可能由于高寒草甸是一个惰性稳定群落,其演替变化极缓慢,进而使两种放牧模式的演替差异较小所致。此外,本研究是在均一生境中开展的较长时间尺度的控制试验,而草地放牧演替和稳定性机制实际是一个非均质生境和长时间尺度下的累积效应。因此,将本研究结果中的草地稳定性结论扩展到长时间尺度和大空间尺度上仍存在很大挑战,还需长期深入系统研究进一步证实。
