拉关木小苗在白骨壤林内及林缘生长实验研究
2019-09-21周亮王秀丽林娴慧刘可鑫卢昌义
周亮, 王秀丽, 林娴慧, 刘可鑫, 卢昌义,*
拉关木小苗在白骨壤林内及林缘生长实验研究
周亮1, 2, 王秀丽1, 林娴慧1, 刘可鑫1, 卢昌义1, 2,*
1. 河口生态安全与环境健康福建省高校重点实验室(厦门大学嘉庚学院), 漳州 363105 2. 厦门大学环境与生态学院, 厦门 361102
探讨引进的红树植物拉关木()是否对本土红树植物白骨壤林()造成入侵风险。通过野外种植实验, 将1年生拉关木苗种植在成熟白骨壤林的林内和林缘, 每月观察记录并测量其生长指标。结果表明: 白骨壤林内和林缘的1年生拉关木苗的月均苗高增量变化趋势相似, 均表现为夏季高, 冬季低; 月均基径增量表现为林内低于林缘, 且随着种植时间的延长而降低。白骨壤林内和林缘的拉关木苗野外种植13个月后的苗高增量仅为14.1 cm和13.5 cm, 基径增量仅为1.0 mm和1.5 mm, 叶片数增量均为负值, 分别为-0.4片和-0.7片, 可见外来种拉关木苗在厦门海沧湾的白骨壤林内及林缘生长速度缓慢, 不具有速生快长特性, 因此, 外来红树植物拉关木只有在其适宜生长的环境才能表现出速生的特性。种植13个月后, 白骨壤林内和林缘拉关木苗的存活率仅为4.7%和5.3%。表明由于光照强度、低温等环境因子的限制, 引进种拉关木小苗很难通过自然扩散在厦门本土红树植物白骨壤林内自然更新生长。
拉关木苗; 白骨壤林; 光照强度; 生长指标; 外来种
0 前言
拉关木〔〕(L.) Gaertn. F., 属于使君子科(Combretaceae), 是真红树植物的一种, 广泛分布于南美、西印度群岛、百慕大群岛、西非以及佛罗里达沿岸[1]。拉关木属于速生常绿乔木, 其高度通常小于15 m, 胸径小于30 cm, 在特定环境下树高会达到25 m, 胸径达到70 cm[2]。由于经济发展, 沿海地区过度开发, 导致毗邻的红树林区域面积缩小, 中国现有的天然红树林面积目前仅为15000 hm2[3]。为提高我国红树林的种类多样性和河口海岸带红树林防护工程质量, 中国林业科学研究院热带林业研究所的研究人员于1999年从墨西哥拉巴斯市首次引入拉关木, 种植于海南东寨港红树林保护区的引种园内[4]。近年来, 在广东、福建等地也陆续引进该种造林。由于拉关木的速生快长特性, 其是否会对本土红树植物造成入侵值得关注。影响红树林下层植被分布的因素主要是: 光照强度、潮汐、种子大小及植物本身的生长特性, 其中, 光照强度的影响受到学者们普遍关注[5]。陈玉锋等[6]研究表明, 光照是限制植物红毛五加()光合速率的主要影响因素。众多的研究者对红树林的深入研究均表明: 光照强度对红树林的生长、发育、繁殖、种群变化和群落演替均有着重要的影响[7-12]。生态入侵的表现往往体现在对光照条件“生态位”的竞争方面。作为外来引进种, 拉关木与乡土红树植物生态位的关系值得关注。目前尚未见拉关木苗在本土红树植物白骨壤林内及林缘的生长情况的研究报道。
本文选取光照强度这个生态限制因子, 分析拉关木通过树冠间隙(Canopy gap)与本地已成林红树植物白骨壤林的竞争可能性和动态过程。钟才荣等[13]对红树植物拉关木的引种效果调查研究表明, 6个月生的拉关木苗在海南东寨港的苗高比广东电白高出6 cm, 而基径则比广东电白的小0.2 cm。表明苗高和基径可以反映不同省份地区拉关木苗的生长量情况。杨虎彪和刘国道[14]对不同光照强度对幼龄期鹧鸪茶生长的影响的研究结果表明, 不同光照强度对鹧鸪茶叶片数及株高均有影响, 光照强度73%的鹧鸪茶叶片数明显高于光照强度28%处理。因此, 本研究通过1年生拉关木苗在乡土红树植物白骨壤林的林内及林缘的苗高、基径、叶片数的测定, 对1年生拉关木苗的生长过程和状态进行定性、定量分析, 旨在了解拉关木在低光强环境中的生态特性及其在乡土林内生境中的生态入侵风险评估, 为今后合理引进该树种以及红树林生态系统的保护管理提供参考。
1 材料与方法
1.1 试验地自然概况
研究区域位处福建省厦门市海沧区海沧湾沿岸的红树林区(118°03′E, 24°29′N)。该区属南亚热带海洋性季风气候, 年平均气温21.3 ℃, 1月份平均气温13.4 ℃, 7月份平均气温29.4 ℃, 极端最低气温4.3 ℃, 极端最高气温36.6 ℃, 年日照时间1971 h, 年降水量1084.5 mm[15]。试验样地为成熟的天然白骨壤林, 郁闭度约65.0%, 高1.8—2.0 m, 基径5.0—7.0 cm。试验样地白骨壤林生长区为中潮带。
1.2 试验材料
2014年6月从广东省湛江市徐闻县红树林育苗苗圃选购1年生拉关木苗, 将拉关木苗放厦门大学漳州校区生化楼A栋1楼的廊道空间, 温度、光照均较稳定, 将拉关木苗摆在育苗盆内, 定期给小苗浇水, 定植培养1个月。
1.3 试验设计
所选的试验样地厦门市海沧区海沧湾沿岸的白骨壤林情况如图1所示, 根据现场试验样地实际情况的考察, 该片白骨壤林附近光滩的底质和白骨壤林内、林缘的差别大, 白骨壤林试验样地附近光滩的底质主要为细石头和砂质, 而白骨壤林内主要为黑色底泥。由于本试验主要研究外来红树植物拉关木是否会对本土红树植物白骨壤林有入侵的风险, 考虑的主要环境因素是光因子, 而离白骨壤林样地较远的光滩也会因底质、高程、海浪等环境因子的不同, 不适合作为对照样地。因此, 本研究根据试验样地的现场情况设置了白骨壤林的林内及林缘各设3个重复的平行样地, 共6个实验样地, 选取高度一致、生长良好的1年生拉关木苗300株, 每个样地种植50株。试验样地林内、林缘的底质、温度、滩涂高程等相近。实验前对每株实验苗悬挂标志牌, 标注各实验苗的编号。同时对实验苗各生长指标进行测量, 测量其基径、株高、叶片数, 作为日后实验数据对比值。种植后, 以30 d为监测周期, 于每月底测量各样地拉关木苗生长指标(苗高、基径、叶片数)测定, 并统计各样地拉关木苗的成活率。
1.4 野外光照强度监测
采用台湾泰士TES—1332数位式照度计(单位), 分别测定各样地林外、林缘、林内光照强度, 以种植的拉关木苗顶端的高度为光强测量高度, 选择晴朗天气, 于退潮期, 对每个样地以每小时为频度进行光照强度测定, 取其平均值作为该样地的光照强度。
1.5 数据处理
用Excel进行统计分析和作图, 采用SPSS 17.0软件对数据进行单因素方差分析, p<0.05表示差异显著。

Figure 1 The test plots offorest in Haicang Bay, Xiamen City
2 结果与分析
2.1 白骨壤林各样地的光照强度情况
2015年2月, 选择晴朗天气, 样地处于退潮期, 采用台湾泰士TES—1332数位式照度计(单位Lux), 分别测定各样地林外、林缘、林内的光照强度,测定结果如图2所示, 样地白骨壤林缘的光照强度为林内的2.7倍。
2.2 白骨壤林内及林缘拉关木苗生长动态分析
由图3可知, 白骨壤林内、林缘1年生拉关木苗种植初期(即2014年7月底), 其月均苗高增量较低, 仅为1.13 cm和0.99 cm; 野外种植2个月后(即2014年8月底), 白骨壤林内和林缘1年生拉关木苗的月均苗高增量分别达3.48 cm和2.75 cm。林内和林缘的月均苗高增量变化趋势相似, 均表现为夏季高, 冬季低。由于2014年12月至2015年3月, 极端低温的胁迫, 导致2015年2月至4月, 拉关木小苗的苗高增量减少, 次年3月, 林内和林缘的月均苗高增量达最小值, 仅为0.22 cm和0.28 cm。2015年5月后, 随着气温的升高, 月均苗高增量有所增加。
由图4可知, 月均基径增量随种植时间的变化趋势与月均苗高增量相似, 均随着种植时间的延长呈先增加后减少的变化趋势。野外白骨壤林样地种植第1个月, 由于野外环境的适应, 林内和林缘的拉关木小苗的月均基径增量分别仅为0.13 mm和0.19 mm; 林内和林缘的月均基径增量在2014年8月份达最大值, 分别达0.28 mm和0.50 mm; 到2014年11月(即野外种植第5个月)后, 林内和林缘样地的拉关木苗的月均基径增量均少于0.1 mm。
由图5可知, 野外种植第1个月, 白骨壤林内和林缘拉关木苗的月均叶片数增量分别为–0.9片和–0.4片,刚种植一个月内出现落叶现象主要原因是: 1年生拉关木苗从室内定植培养到野外环境, 由于刚种植的苗根系有轻微受损, 且野外由于昼夜温差大、海浪等环境因子的干扰, 需要一段时间的定植恢复期。但在2014年8月份(即野外种植第2个月), 拉关木苗的月均叶片数增量有所增加, 林内和林缘的月均叶片数增量分别为0.6片和4.0片。在2014年9月份至2015年4月份期间, 白骨壤林内和林缘的1年生拉关木苗的月均叶片数增量均为负值, 出现落叶现象; 2015年5月份, 林内和林缘样地的拉关木苗的月均叶片数增量均有所增加, 分别达1.6片和2.4片。但在2015年6月份, 白骨壤林缘的1年生拉关木苗的月均叶片数增量减少, 到2015年7月份时出现叶片凋落现象; 而2015年6—7月份, 林内的1年生拉关木苗的叶片再次出现凋落现象, 海沧白骨壤林内及林缘1年生拉关木苗叶片数随时间变化出现波动情况, 可能与样地外围商业船舶、渔船的出入, 导致对白骨壤林样地造成较大的海浪冲击有关。
Figure 2 Light intensity offorests in Haicang Bay in February 2015
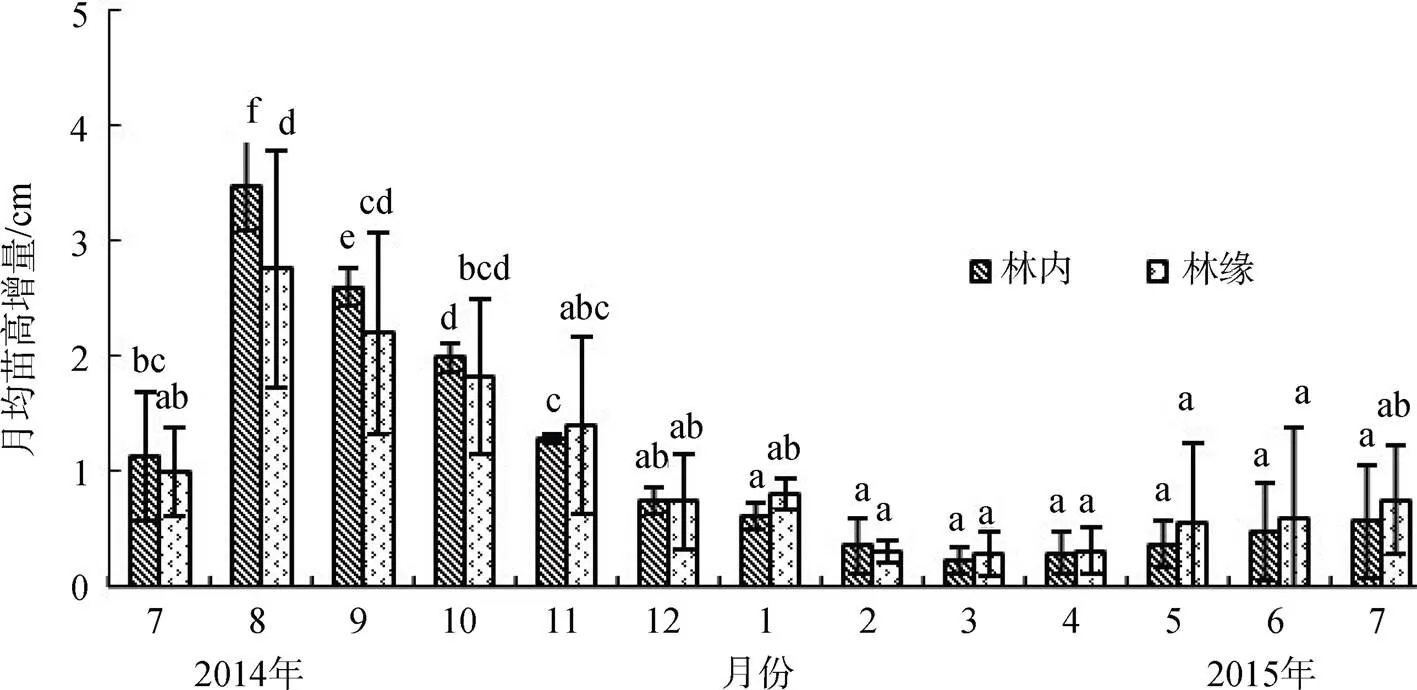
图3 不同光照强度下白骨壤林的拉关木小苗月均苗高增量
Figure 3 Monthly average height increment ofseedlings under different light intensity inforests

图4 不同光照强度下白骨壤林的拉关木小苗月均基径增量
Figure 4 Monthly average base diameter increment ofseedlings under different light intensity inforests
由表1, 表2可见, 在2014年7月至2015年6月期间, 海沧白骨壤林内的1年生拉关木小苗的存活率均高于林缘。由于3个试验样地海沧白骨壤林内的郁闭度仅为60.1%—65.3% (如表1所示), 因此白骨壤林内的1年生拉关木苗能存活较长时间, 但到2015年7月, 白骨壤林内和林缘的拉关木苗的存活率仅为4.7%和5.3%。
3 讨论
光照是植物生长必不可少的要素, 光照强度的强弱直接影响到植物的光合作用, 最终影响植物的生长。本研究结果表明, 野外种植6个月的时间内, 成熟白骨壤林内的1年生拉关木苗的月均苗高增量普遍高于林缘, 2014年8月份, 白骨壤林内1年生拉关木苗的月均苗高增量比林缘高26.5%, 出现这种情况的原因可能是因试验样地的白骨壤林的郁闭度为60.1%—65.3%, 光线均能透进白骨壤林内, 弱光促进了1年生拉关木苗的苗高的增长。这与弱光对无瓣海桑幼苗的茎高有促进作用[16]的结果相似。有研究表明, 弱光对多数植物幼苗和秋茄()胎生幼苗的初期生长有促进作用的研究结果相似[8, 17]。这可能是低光照导致红树林植物的光合速率下降, 植物生长激素分泌增加, 从而致使植物个体的茎高增加[5]。白骨壤林缘和林内样地的1年生拉关木苗的月均苗高增量在种植第2个月时均达到最大值, 分别达3.48 cm和2.75 cm。种植第3个月后, 林内和林缘的1年生拉关木苗的月均苗高增量、月均基径增量、及月均叶片数增量均呈下降趋势, 表明长时间的弱光条件不利于拉关木幼苗的生长; 这与刁俊明等[18]研究表明: 无瓣海桑在荫蔽初期(2—4个月), 弱光环境对幼苗的茎高的增加有促进作用, 随着荫蔽时间的延长则产生抑制作用, 不利于幼苗的生长的结论一致。钟才荣等[13](2011)研究表明, 拉关木在海南东寨港保护区6个月龄的苗高为86 cm, 在广东电白红树林苗圃6个月龄的苗高为74 cm。而本研究中, 白骨壤林内和林缘的拉关木苗野外种植13个月的苗高增量仅为14.1 cm和13.5 cm, 基径增量仅为1.0 mm和1.5 mm, 可见外来种拉关木苗在厦门海沧湾的白骨壤林内和林缘的生长速度极慢, 不具有速生快长特性, 因此, 外来红树植物拉关木只有在其适宜生长的环境下才能表现出速生快长的特性。

图5 不同光照强度下白骨壤林的拉关木小苗月均叶片数增量
Figure 5 Monthly average number of leaves increment ofseedlings under different light intensity inforests

表1 不同光照强度下白骨壤林的拉关木小苗2014年的存活率

表2 不同光照强度下白骨壤林的拉关木小苗2015年的存活率
冬季至次年的初春季, 白骨壤林内和林缘的1年生拉关木苗的月均苗高增量、月均基径增量均非常低, 在次年3月份1年生拉关木苗的月均苗高增量达最小值, 分别为0.22 cm和0.28 cm, 叶片出现凋落, 这可能是因为低温导致1年生拉关木小苗的生长缓慢甚至出现落叶现象。因本土红树植物白骨壤是自然分布最北的抗低温广布种红树[19], 表明不耐低温的引进种拉关木小苗难于在福建较北的地方占据本土红树植物白骨壤林的生态位。
本研究结果表明, 1年生拉关木苗在白骨壤林内和林缘种植一个月后, 存活率为86.0%和74.0%, 随着种植时间的延长, 其存活率不断减少, 且林内1年生拉关木苗的存活率普遍低于林缘, 这可能是因为试验样地白骨壤林为海沧湾的入海口处, 该试验片区的海浪冲击大, 白骨壤林缘的1年生拉关木苗由于海浪冲击大, 而白骨壤林内海浪冲击小, 导致拉关木苗种植1年时间里, 林内的存活率均高于林缘。种植6个月后, 林内和林缘拉关木苗的存活率为45.3%和32.0%, 到次年7月份林内和林缘的1年生拉关木苗的存活率仅为4.7%和5.3%。因此, 由于光照强度、低温等环境因子的限制, 引进种拉关木小苗很难通过自然扩散在厦门本土红树植物白骨壤林内自然更新生长。
[1] NELLIS D. Seashore plants of South Florida and the Caribbean[M]. Sarasota: Pineapple Press, 1994: 164.
[2] JIMENEZ J, MARTINEZ R, ENCARNOCION L. Massive tree mortality in a Puerto Rican mangroves forests[J]. Caribbian Journal of Science, 1985, 21:75–78.
[3] PENG Y, ZHOU Y, Chen G Z. The restoration of mangrove wetland: a review[J]. Acta Ecologica Sinica, 2008, 28: 786–797.
[4] 廖宝文, 郑松发, 陈玉军, 等. 海南东寨港几种国外红树植物引种初报[J]. 中南林学院学报, 2006, 26(3): 63–67.
[5] 刁俊明, 彭逸生, 郑明轩, 等. 红树植物对低光照水平的生长及生理生态响应研究进展[J]. 嘉应学院学报, 2009, 27(3): 69–74.
[6] 陈玉锋, 黄旭峰, 古锐, 等. 不同光照强度下红毛五加光合及生理特性研究[J]. 中国中药杂志, 2018, 43(5): 926–933.
[7] SMITH T. Effects of seed predators and light level on the distribution of(Forsk) Vierh in topical, tidal forests[J]. Estuarine, Coastal and Shelf Science, 1987, 25(1): 43–51.
[8] TAMAI S, IANPA P. Establishment and growth of mangrove seedlings in mangrove forests of southen Thaliland[J]. Ecological Research, 1988, 3(3): 227–238.
[9] ALLEN J, KRAUSS K, HAUFF R. Factors limiting the intertidal distribution of the mangrove species[J]. Oecologia, 2003, 135(1): 110–121.
[10] PROFFITT C, DEVLIN D. Long-term growth and succession in restored and natural mangrove forests in southwestem Florida[J]. Wetlands Ecology and Management, 2005, 13(5): 531–551.
[11] IMAI N, TAKYU M, NAKAMURA Y, et al. Gap formation and regeneration of tropical mangrove forests in Ranong Thailand[J]. Plant Ecology, 2006, 186(1): 37–46.
[12] SANTIAGO L, LAU T, MELCHER P, et al. Morphological and physiological responses of Hawaiian hibiscus tiliaceus populations to light and salinity[J]. International Journal of Plant Sciences, 2000, 161(1): 99–106.
[13] 钟才荣, 李诗川, 杨宇晨, 等. 红树植物拉关木的引种效果调查研究[J]. 福建林业科技, 2011, 38(3): 96–99.
[14] 杨虎彪, 刘国道. 不同光照强度对幼龄期鹧鸪茶生长的影响[J]. 热带作物学报, 2017(11): 2056–2059.
[15] 厦门市统计局, 国家统计局厦门调查队. 厦门经济特区年鉴—2015 [M]. 北京: 中国统计出版社, 2015.
[16] 刁俊明, 陈桂珠. 光强对无瓣海桑幼苗的生长和光合特性的影响[J]. 林业科学研究, 2008, 21(4): 496–492.
[17] PROFFITT C E, DEVLIN D J. Long-term growth and succession in restored and natural mangrove forests in southwestern Florida[J]. Wetlands Ecology and Management, 2005, 13(1): 531–551.
[18] 刁俊明, 曾宪录, 陈桂珠. 无瓣海桑幼苗对不同遮光度的生理生态响应[J]. 生态学杂志, 2010, 29(7): 1289– 1294.
[19] 林鹏. 红树林[M]. 北京: 海洋出版社, 1984.
Growth ofseedlings inside and at the edge offorests
ZHOU Liang1,2, WANG Xiuli1, LIN Xianhui1, LIU Kexin1, LU Changyi1,2,*
1. Key Laboratory of Estuarine Ecological Security and Environmental Health (Xiamen University Tan Kah Kee College), Fujian Province University, Zhangzhou 363105, China 2. College of the Environment & Ecology, Xiamen University, Xiamen 361102, China
In order to assess the invasion risk of fast-growing introduced mangrove species, field experiments of planting 1-year-oldseedlings were used to study the effects of environmental factors of light intensity on their growth inside and at the edge of native mangrove speciesforest. Results showed that the average monthly shoot height of 1-year-oldseedlings inside and at the edge of theforests was similar, both high in summer and low in winter; the average monthly shoot diameter of 1- year-oldseedlings inside theforests was lower than that inforest margins, and the average monthly shoot diameter increment ofseedlings decreased with the prolongation of planting time. After planting for 13 months, the seedling height ofinside and at the edge of native mangrove speciesforest was only 14.1 and 13.5 cm, respectively; and the base diameter increment was only 1.0 and 1.5 mm, respectively. The average monthly number leaves of 1-year-oldseedlings inside and at the edge of theforest decreased to -0.4 and -0.7, respectively. The results indicated that the growth rate of exotic mangrove specieswas slowly inside and at the edge offorestsin Haitang Bay, Xiamen city; and it did not show fast-growing characteristics. Therefore, the exotic mangrove speciescan grow fast only in its suitable environment. The survival rate ofseedlings inside theforests was only 4.7%, and that inforest margin was 5.3%. Due to the limitation of light intensity, low temperature, it is very difficult forseedlings to naturally renew and grow under natural conditions in the natural indigenousmangrove forest in Xiamen.
seedling;forests; light intensity; growth indexes; exotic spe
10.14108/j.cnki.1008-8873.2019.04.001
Q948.1
A
1008-8873(2019)04-001-06
2018-09-11;
2018-11-05
国家重点研发计划“典型脆弱生态修复与保护研究”重点专项(2017YFC0506103); 国家自然科学基金(41376115), 福建省中青年教师教育科研项目(JA15618), 福建省大学生创新创业训练计划项目(201913469024)
周亮(1981—), 男, 福建厦门人, 讲师, 主要从事红树林湿地生态及环境遥感方面的研究, E-mail: zhou812@xujc.com
卢昌义, 教授, 博士生导师, 主要从事红树林湿地海岸生态恢复工程方面研究, E-mail: lucy@xmu.edu.cn
周亮, 王秀丽, 林娴慧, 等. 拉关木小苗在白骨壤林内及林缘生长实验研究[J]. 生态科学, 2019, 38(4): 1-6.
ZHOU Liang, WANG Xiuli, LIN Xianhui, et al. Growth ofseedlings inside and at the edge offorests[J]. Ecological Science, 2019, 38(4): 1-6.
