广西弄岗北热带喀斯特季节性雨林林隙特征与空间分布
2019-09-10向悟生李冬王斌李先琨郭屹立文淑均陆树华梁士楚
向悟生 李冬 王斌 李先琨 郭屹立 文淑均 陆树华 梁士楚





摘 要: 林隙是森林中一種普遍存在的小尺度森林干扰形式,对促进森林更新和循环演替具有重要作用。对热带喀斯特森林的林隙干扰目前还了解不多。该文采用Worldview-3高空间分辨率遥感影像结合地面调查,对弄岗喀斯特季节性雨林15 hm2 样地林隙干扰特征及其空间分布规律进行了研究。结果表明:样地内共解译到305个林隙,林隙的密度约为每公顷20个,林隙大小的变化范围为10~371 m2,平均大小为52.3 m2,林隙面积占森林面积的10.6%;林隙在0~50 m尺度整体表现出聚集分布格局;林隙空间分布与大树(DBH≥10 cm)存在显著空间关联,与小树(DBH<10 cm)和枯立木无显著空间关联;不同生态因子对林隙的数量分布和基本结构影响不同,对林隙数量分布影响最显著的包括凹凸度、干旱度指数、丰富度,而坡向、海拔、丰富度对林隙大小有显著影响,海拔、岩石裸露度、地形湿润度指数、干旱度指数、多度等5个生态因子对林隙周长有显著影响。研究结果表明林隙在喀斯特季节性雨林普遍存在,林隙的空间分布与特征存在着地形分异特征,是喀斯特独特的地质地貌和生境条件与喀斯特森林自身生物学特性综合作用的结果。
关键词: 喀斯特季节性雨林, 林隙, 空间分布格局, 空间关联, 生态因子
中图分类号: Q948, P217
文献标识码: A
文章编号: 1000-3142(2019)01-0087-11
Characteristics and spatial distribution of forest gap in a northern tropical karst seasonal rainforest in Nonggang, Guangxi, South China
XIANG Wusheng1,2 , LI Dongxing2 , WANG Bin2 , LI Xiankun2 , GUO Yili2 , WEN Shujun2 , LU Shuhua2 , LIANG Shichu1*
( 1. College of Life Sciences, Guangxi Normal University, Guilin 541004, Guangxi, China; 2. Guangxi Key Laboratory of Plant Conservation and Restoration Ecology in Karst Terrain, Guangxi Institute of Botany, Guangxi Zhuang Autonomous Region and Chinese Academy of Sciences, Guilin 541006, Guangxi, China )
Abstract: Forest gap is the dominant form of disturbance in many forest ecosystems. Gaps play an important role in forest helping to maintain forest regeneration and dynamics. Little is known about the gap pattern and characteristics of karst seasonal rain forests. In this study, high-spatial-resolution Worldview-3 satellite imagery was coupled with field data to assess gap disturbance regime and its pattern in a 15 hm2 northern tropical karst seasonal rain forest dynamic plot in Nonggang National Nature Reserve, Guangxi. Total 305 gaps were identified, ranging in size from 10 to 371 m2 and with mean size 52.3 m2. The canopy gap fraction was 10.6%, and gap density was about 20 per hectare. The spatial pattern of gaps was clustered at scale 0-50 m. Significant association was found between gaps and big trees(DBH≥10 cm), and null associations between gaps and small trees(DBH<10 cm) and between gaps and standing dead trees. The prevailing ecological factors had different influence on gap distribution and gap characteristics. Convexity, altitude above channel(AAH) and tree species richness had significant effect on the gaps distribution. Gap size was significantly influenced by slope, mean elevation and tree species richness. Gap perimeter was mainly affected by mean elevation, rock-bareness rate(RBR), topographic wetness index(TWI), AAH and tree species abundance. These results indicated that gap disturbance was very frequent in karst seasonal rain forests, and there were topographic differentiation in spatial distribution and characteristics of gaps resulted by the combination effects of unique special karst geological conditions, habitats and the biological characteristics of karst forest.
Key words: karst seasonal rain forest, forest gap, spatial pattern, spatial associations, ecological factors
林隙是森林循环中一种典型的干扰过程,是森林生态系统中普遍存在的现象(Runkle,1982)。林隙通过改变森林内的异质性生境,进而对森林群落的更新过程产生重要影响,最终影响到群落的结构和动态过程(Hubbell et al.,1999)。因此研究热带喀斯特森林的林隙特征,进而分析林隙对热带喀斯特森林幼苗更新和森林动态的驱动作用,对热带喀斯特森林生物多样性保护和退化生态系统恢复具有重要科学意义和现实意义。
Brokaw(1982)给出了森林林隙明确的定义。此后,不同森林林隙特征的研究广泛开展。Kathke & Bruelheide(2010)对德国的云杉林研究发现,林隙可占到该森林面积的10%左右,并且随时间而动态变化。Alexander & Mack(2017)对美国阿拉斯加落叶森林的研究结果表明,林隙占森林的面积比例为17%~29%。臧润国和杨彦承(1999)对海南岛热带山地雨林的研究发现,该森林存在丰富的林隙,占森林面积的比例在25%左右。沈泽昊等(2001)对三峡山地常绿阔叶林的研究发现,该森林林隙干扰较频繁,林隙面积可占森林面积的10.09%。闫淑君等(2004)对福建万木林的常绿阔叶林研究发现,林隙可占森林面积的16.66% 。刘少冲等(2013)对小兴安岭阔叶红松混交林林隙特征的研究表明,该森林林隙所占的面积比例为15.71%,大小变化在42.12~372.52 m2之间。吴庆贵等(2013)的研究表明,川西高山冷杉林的林隙密度为14.67·hm-2 ,占森林景观面积的23.05% 。温远光等(2014)的研究结果表明,广西大明山南亚热带山地常绿阔叶林林隙占森林景观面积的52.90%,林隙特征不同于热带山地雨林,也与南亚热带低地的常绿阔叶林存在显著区别。在喀斯特森林的相关研究中,龙翠玲等(2005)对贵州茂兰亚热带喀斯特森林的林隙及其更新特征进行研究,发现林隙干扰在该喀斯特森林中十分频繁和普遍,林隙密度平均每公顷达到20个,比亚热带的山地常绿落叶阔叶混交林和湿性常绿阔叶林的大。综合国内外研究进展可以发现,目前关于林隙的研究多集中于热带雨林、温带森林以及部分亚热带常绿阔叶林,喀斯特森林很少被关注。在热带喀斯特地区,暴雨和大风(台风)等林隙形成的客观条件比中亚热带地区更为频繁,但有关热带喀斯特森林林隙干扰状况、特征等科学问题尚未见有相关研究报道。
从研究手段上看,林隙特征和空间分布的传统研究手段大都是基于地面调查的方法(Schliemann & Bockheim,2011; 胡理乐等,2010)。Schliemann & Bockheim(2011)和Garbarino et al.(2012)研究认为,高分辨率和多光谱遥感影像可以提供地面森林林冠层的细节图像和光谱数据,一般可识别面积10 m2 以上的林冠空隙。我国南方喀斯特山地地形陡峭,小地形复杂,常规方法在较大面积开展林隙的定量研究非常困难(龙翠玲等,2005)。进行较大尺度喀斯特森林林隙的研究,还需要进一步探索相应的可行方法。
广西弄岗国家级自然保护区目前仍保存着我国面积最大、生态系统结构最完整、最典型的北热带喀斯特季节性雨林。喀斯特季节性雨林是喀斯特森林的典型代表,生物多样性十分丰富且含有很多重要的特有种(王斌等,2014)。本研究基于2011年建成的弄岗15 hm2 喀斯特季节性雨林固定监测样地,利用高分辨率和多光谱遥感数据结合地面的精细调查开展研究。主要科学问题包括:(1)喀斯特季节性雨林林隙是否在样地尺度普遍存在;(2)林隙的特征和空间分布规律如何;(3)林隙的空间分布和特征主要受何种生态因子影响。本研究旨在为喀斯特这种极度崎岖地形中森林林隙的研究提供方法参照,为热带喀斯特退化森林生态系统的恢复提供科学依据。
1 研究区概况
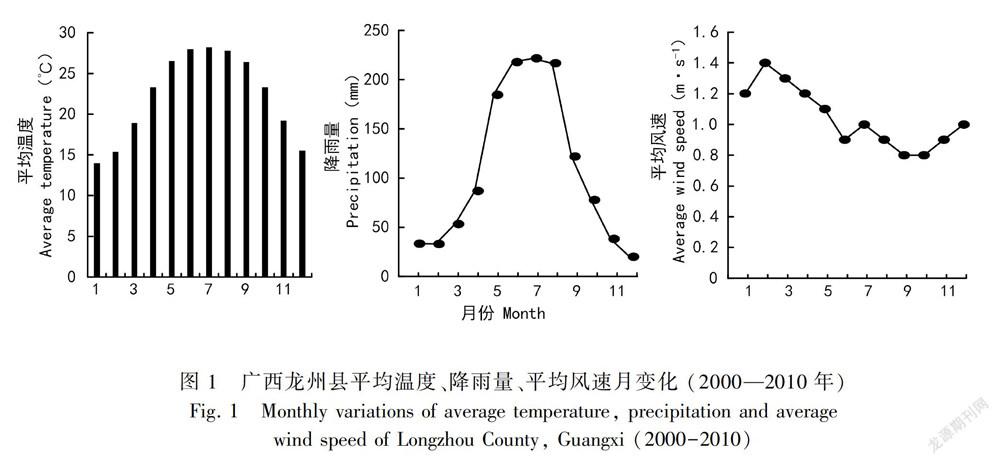
弄岗国家级自然保护区处在宁明县西北部和 龙州县中北部,地理位置为106°42′28″—107°04′54″ E, 22°13′56″—22°39′09″ N,面积约101 km2 。该区地貌类型主要为峰丛谷地型和峰丛洼地型,区内地形十分复杂,山弄密集;土壤主要为石灰土,包括原始石灰土、棕色石灰土、黑色石灰土、水化棕色石灰土等类型。研究区为热带季风气候,春夏秋三季主要受印度洋北部西南季风和赤道季风影响,而冬季则主要受大陆高压东北季风影响。区内月平均降雨分布不均,干湿季节明显,旱季主要在1—4月和11—12月,雨季集中在5—9月,多年平均降雨量在1 150~l 550 mm之间。年均气温22 ℃,年最高气温37~39 ℃,最冷月平均气温在13 ℃以上。由于受季风环流和地形条件影响,平均风速常年较小,一般在1.4~1.5 m·s-1 之间,平均风速较大的为1—4月,平均风速较小的一般为9—10月,风向与谷向变化基本一致(图1)。
依照热带森林科学中心(Center for Tropical Forest Science, CTFS)的标准,弄岗15 hm2 森林动态监测样地(106°57′ E, 22°25′ N)于2011年在弄岗保护区内建成。该样地建成后被纳为CTFS样地网络和中国森林生物多样性监测网络重要节点,是热带区域唯一的喀斯特森林监测样地。该样地南北和东西边长分别为300 m和500 m,海拔高差為180~370 m(向悟生等,2017)。样地包括了峰丛洼地地貌比较典型的生境类型,异质性很高,洼地、山坡、峰顶等生境类型均有分布。丰富的生境类型造就了较高的生物多样性,样地内分布有DBH ≥ 1 cm木本植物223种(不包括藤本植物),分属于56科157属,树种科、属的区系均以热带成分为主,优势科包括大戟科(Euphorbiaceae)、马鞭草科(Verbenaceae)、梧桐科(Moraceae)等(王斌等,2014)。
2 研究方法
2.1 林隙特征获取与空间格局分析
采用2015年研究区的Worldview-3全色和多光谱遥感影像进行林隙的遥感解译。遥感数据的空间分辨率为0.3 m,可以提供比较清晰的林冠信息。获取遥感影像数据后,在ENVI5.3软件中进行大气校正和正射校正,并对全色数据与多光谱数据进行融合增强处理。根据15 hm2 样区四个角的精确地理坐标,切割获取样地遥感数据,通过监督分类的方式提取面积大于10 m2 的林冠空隙信息,详细的方法见参考文献(Garbarino et al.,2012)。根据提取到的15 hm2 样区林隙信息,野外对每个林隙进行核实,并用鱼眼相机在林隙中心1.0 m处垂直向上拍摄半球面影像,室内根据鱼眼照片,在ArcGIS软件支持下对林隙特征进行校正,并计算林隙面积和周长等特征。在ArcGIS软件中,分别建立两个字段X和Y,右键单击选择“计算几何”,在属性中分别选择X Coordinate of Centroid或者Y Coordinate of Centroid用于计算各林隙的中心坐标。
以林隙中心点坐标为基础,采用单变量的双关联函数g(r)分析林隙的空间分布格局。双关联函数g(r)由Ripley’s K 函数推演而来(Wiegand et al.,2004)。其公式如下:
g(r)= dK(r) dr /2πr。
式中,K(r)为以某一林隙中心点为圆心、r为半径的圆内,期望林隙点数与圆内点密度的比值(Wiegand et al.,2004)。当g (r)>1 时,为聚集分布;当 g (r)<1 时,则表示均匀分布;若g (r)=1,则表示随机分布。采用的完全随机零假设作为零模型,进行199 次Monte Carlo随机模拟,将模拟结果中第5个最大、最小模拟值之间的数值范围作为置信区间,显著性水平α的值为 0.05(置信区间为95%)以检验分析结果的显著性。研究分析的尺度范围为0~50 m。为了减小Ⅰ类误差,采用适合度检验(goodness of fit,GOF)分析林隙的空间分布是否显著偏离零模型(Baddeley et al.,2014)。适合度检验将双关联函数中包含的尺度依赖信息压缩到一个统计检验量ui ,ui 为研究尺度范围内实际观测值与零模型模拟值的平方偏差。检验的P值可以根据下式计算:
P ^ =1- s j=1 I(u0>uj) s+1 。
式中,I(u0 >uj )为指示函数,当u0 >uj 时其值取0,否则取1;s为Monte Carlo随机模拟的次数。详细的方法可见参考文献(Loosmore & Ford,2006)。
空间格局的分析和作图采用R 3.4.3软件及其中的“spatstat”软件包完成(R Core Team,2017)。
2.2 林隙与树木的空间关联
为了检测不同树木对林隙形成的作用,将树木分成大树、小树和枯立木,分别检测它们在空间上与林隙的关联情况。基于野外调查获取的冠幅和树高等参数,大树的选取标准为DBH≥10 cm,小树则为DBH<10 cm的树木个体;同时区分大径级的枯立木和所有径级枯立木进行分别分析,大径级枯立木标准与大树一致,即枯立木DBH≥10 cm的个体。采用双变量双关联函数g12 (r)分析林隙与树木之间的空间关联。双变量双关联函数的计算与单变量双关联函数一致,计算时K12 (r)为以某一林隙中心点为圆心、r为半径的圆内,期望树木点数与圆内树木点密度的比值。当g12 (r)>1 时,表示两者存在正关联;当 g12 (r)<1 时,则表示存在负关联;若g12 (r)=1 则表示两者无关联。空间关联的显著性检验和整体尺度的适合度检验和单变量双关联函数一致。
2.3 林隙特征与重要生态因子的关系
将15 hm2 样地划分为375个20 m× 20 m样方,统计中心点落于样方内的林隙数量,并计算林隙面积、周长等基本特征。同时计算每个林隙中心点所在20 m × 20 m样方的重要生态因子,包括地形环境因子的坡度、凹凸度、坡向、海拔、地形湿润度指数(topographic wetness index, TWI)以及干旱度指数(altitude above channel, AAH),相关计算方法见参考文献(郭屹立等,2016);在野外先将15 hm2 样地划分为6 000 个5 m × 5 m的小样方,由二人分别调查样方的岩石裸露度,并取平均值获得每个小样方的岩石裸露率;再通过均值获得每个20 m × 20 m样方的岩石裸露率,得到7个地形相关的生态因子。生物因子主要为每个林隙中心点所在20 m × 20 m样方DBH≥ 1 cm植株的多度、丰富度、平均胸径、最大胸径和胸高断面积等5个生物因子,共计12个生态因子。以20 m× 20 m样方林隙数量、面积、周长分别为因变量,12个生态因子为自变量,采用广义线性模型(GLM)分析林隙空间分布、林隙特征与生态因子的关系。以林隙数量为计数数据,采用Poisson分布的广义线性模型分析林隙分布与生态因子的关系;林隙面积和周长符合正态分布,因此采用Gaussian分布的广义线性模型分析林隙特征与生态因子的关系。采用赤池信息准则(akaike information criterion, AIC)進行模型拟合的适合度分析,AIC值越小模型的拟合优度越高(Burnham & Anderson,2002)。依据AIC指数,使用向后剔除法逐步筛选最优模型,确定对林隙分布和林隙特征有明显影响的生态因子。
广义线性模型拟合在R 3.4.3软件中使用glm函数进行,使用step函数进行模型的逐步筛选(R Core Team,2017)。
3 结果与分析
3.1 林隙特征与空间分布
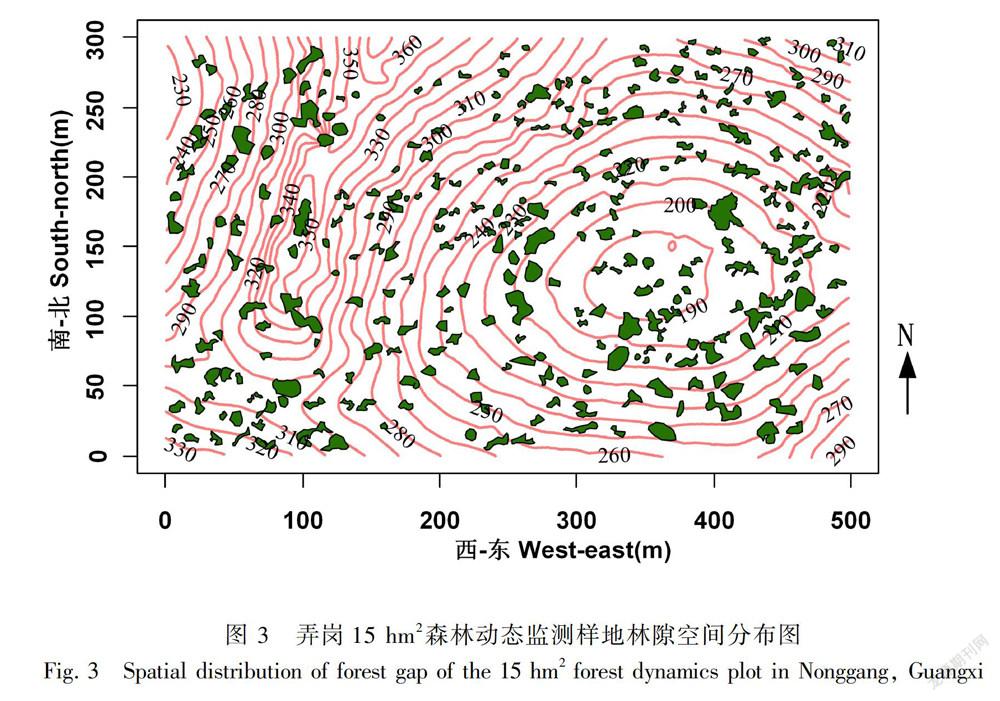
依据高清遥感解译与地面核查校验的结果,Worldview-3遥感影像解译结果具有较高的精度,解译正确率可以在90%以上。15 hm2 的样地中共解译到305个面积大于10 m2的林冠空隙,林隙的密度每公顷约为20个(图2,图3)。305个林隙的总面积为15 956.5 m2,约占样地面积的10.6%。林隙的平均大小為52.3 m2,林隙大小的变化范围为10~371 m2。
从林隙大小分级统计结果看,小型林隙所占的比例较大,10~40 m2的林隙约占总林隙数的54%;40~80 m2的林隙数量较多,占比在30%左右(图2);而面积超过100 m2的林隙数量较少,15 hm2样地中只有28个,占比约为12%。
从林隙空间分布格局分析结果看,林隙的空间分布格局随尺度而变化。在较小尺度时(<6 m),林隙空间分布格局为均匀分布;在6~12 m尺度内,林隙的空间分布格局表现为随机分布;而在13~28 m尺度范围时,林隙的空间格局表现为聚集分布。在尺度大于28 m后,林隙的空间分布格局在随机格局和聚集分布格局间交替变化(图4)。
根据GOF的检验结果来看,林隙空间分布格局在0~50 m尺度整体显著偏离随机零模型(P<0.05),表现出聚集分布的格局。
3.2 林隙分布与树木的空间关联
林隙与所有枯立木的空间关联分析结果显示,在0~50 m尺度范围内两者的双变量双关联函数g12 (r)值均在模拟的上下包迹线范围内,显示出无关联的特征;GOF适合度检验表明,两者空间关联在整体尺度上未显著偏离随机零模型(P>0.05)(图5),即整体上表现为空间无关联。林隙与大径级枯立木也显示出空间无关联特征(P>0.05)(图5)。
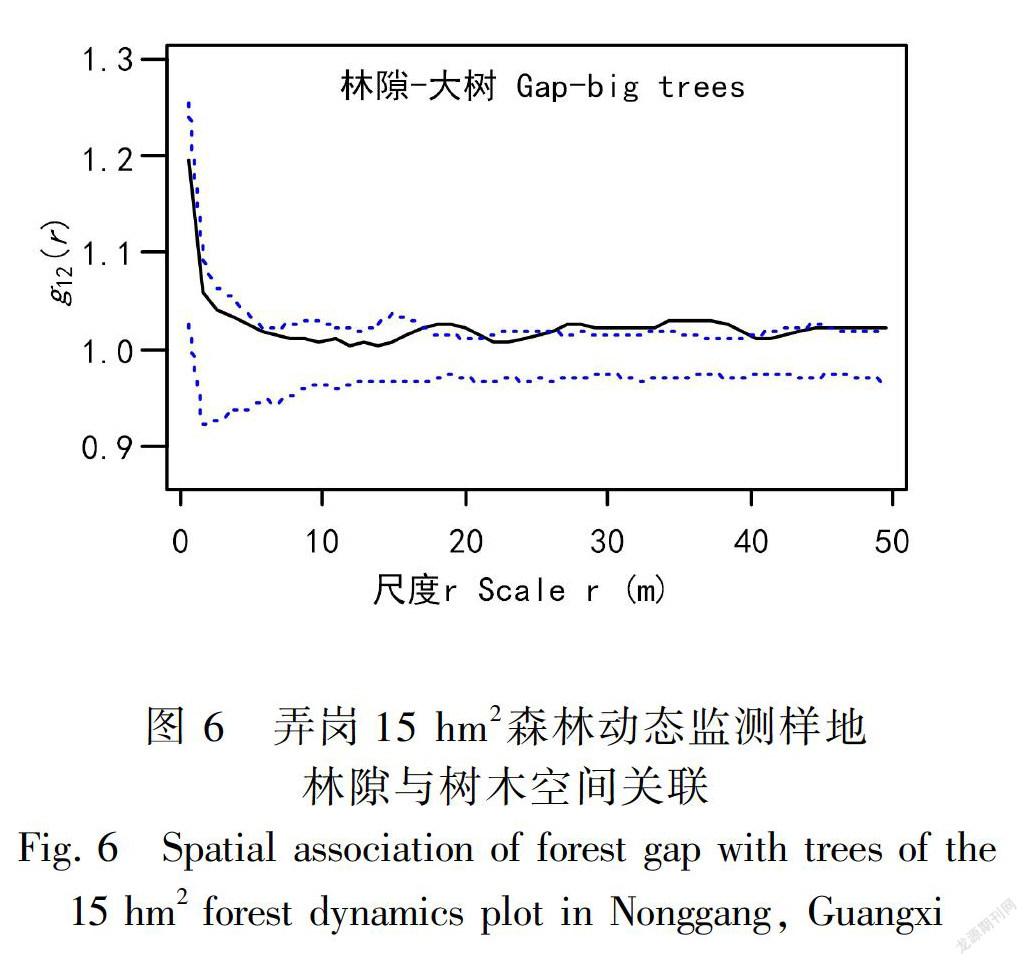
林隙分布与大树的空间关联分析结果显示,在0~18 m尺度范围内,两者的双变量双关联函数g12 (r)值均在模拟的上下包迹线范围内,表现为无关联特征;在19~50 m尺度,显示出正关联和无关联交替变化的特征。GOF检验表明,两者空间关联在整体尺度上显著偏离随机零模型(P<0.001 )(图6),即整体上表现为显著的空间正关联。
林隙与小树的空间关联分析结果显示,在0~50 m尺度范围内两者的双变量双关联函数g12 (r) 值均在模拟的上下包迹线范围内;GOF适合度检验表明,两者空间关联在整体尺度上未显著偏离随机零模型(P>0.05)(图6),整体上表现为空间无关联。
3.3 林隙特征与生态因子的关系
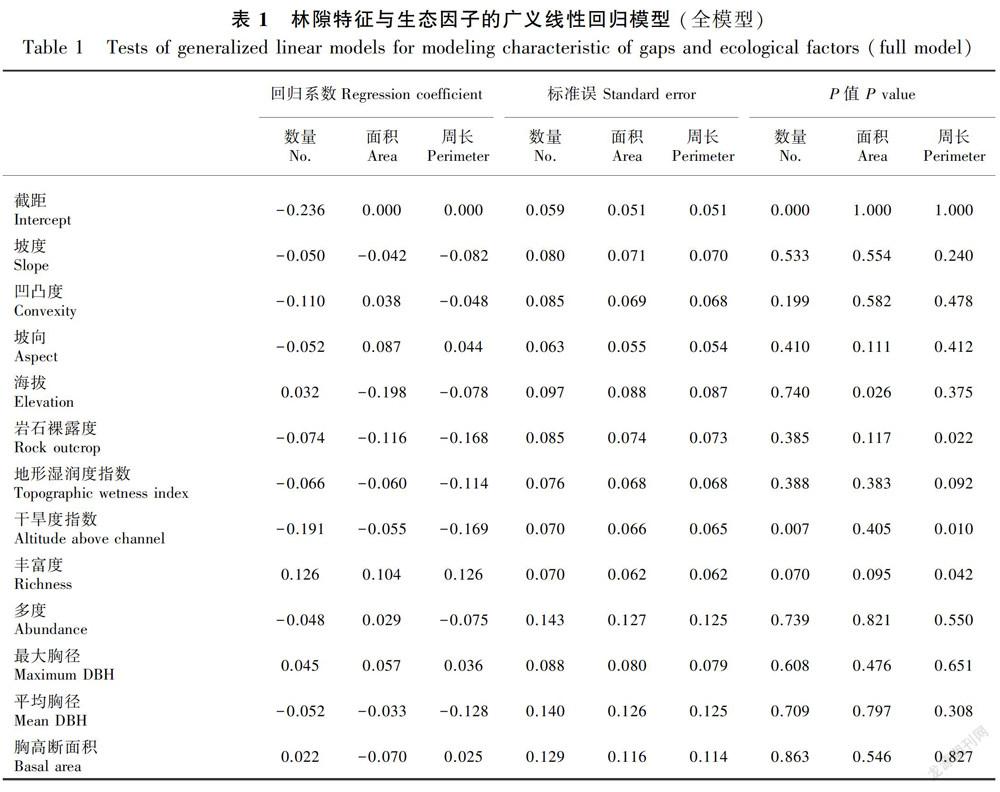
为检验不同生态因子对林隙数量分布、林隙特征的作用,首先进行全模型拟合,然后添加所有12个生态因子进入模型进行初步分析。表1结果表明,凹凸度、干旱度指数、物种丰富度指数与林隙数量有较大的标准化回归系数,林隙数量与所有生态因子全模型的AIC指数为860.7;林隙面积与坡向、海拔、岩石裸露度、干旱度指数的标准化回归系数较大,全模型的AIC值为1 070.4 ;林隙周长与岩石裸露度、地形湿润度指数、干旱度指数、丰富度、平均胸径的标准化回归系数较大,其全模型的AIC值为1 062.6 。
通过使用向后逐步剔除变量选择的方法,筛选林隙数量、林隙特征与生态因子关系的最优模型(表2)。从筛选得到的最优模型来看,对林隙数量分布影响最显著的包括凹凸度、干旱度指数、丰富度三个生态因子,其中凹凸度和干旱度指数对林隙数量有显著的负影响,丰富度对林隙数量有显著正影响,最优模型的AIC值从全模型的860.7降至845.9;林隙面积与坡向、海拔、丰富度有显著关系,海拔对林隙面积大小有负影响,而坡向和丰富度对林隙面积大小为正影响,最优模型的AIC值由全模型的1 070.4 降至1 058.1 ;海拔、岩石裸露度、地形湿润度指数、干旱度指数、多度等5个生态因子对林隙周长有显著影响,其中,海拔、岩石裸露度、地形湿润度指数和干旱度指数为负影响,而多度为正影响,最优模型的AIC值从全模型的1 062.6 降至1 053.7 。
4 讨论
4.1 林隙特征提取与空间分布
从提取的林隙信息分析结果看,热带喀斯特季节性雨林的林隙密度偏低,与处于亚热带的茂兰喀斯特森林非常接近(龙翠玲等,2005),稍高于部分高山针叶林和山地常绿阔叶林(Kian et al.,2017; 沈泽昊等,2001; 吴庆贵等,2013),但低于热带雨林和部分南亚热带山地常绿阔叶林(温远光等,2014; 臧润国和杨彦承,1999);喀斯特季节性雨林林隙面积占森林面积的比例和平均林隙面积与部分常绿阔叶林和高山针叶林等相近(Kian et al.,2017; 沈泽昊等,2001; 吴庆贵等,2013),低于热带雨林、北方针叶林与部分其它常绿阔叶林等类型(刘少冲等,2013; 温远光等,2014; 臧润国和杨彦承,1999),也低于亚热带喀斯特森林(龙翠玲等,2005)。林隙的密度与大小等与森林的发育年龄、地形和气候气象因子密切相关(Kian et al.,2017; Muscolo et al.,2014),也与研究的方法有关(Garbarino et al.,2012; 毛学刚等,2018)。弄岗喀斯特季节性雨林所处地形为典型的峰丛洼地地貌,森林位于环状深切圆洼地中,大风等气象因素对林隙的形成贡献较小;而且喀斯特整体的生境较为恶劣,树木生长缓慢,平均胸径相比其他森林偏小,这些原因应该与弄岗喀斯特季节性雨林林隙密度偏低和林隙面积较小的特征密切相关。
弄岗喀斯特季节性雨林林隙整体上表现为聚集分布的格局。这与一些针叶林的研究结果一致(Garbarino et al.,2012)。空间异质性和人为干扰因素被认为是影响林隙空间分布格局较为重要的因素(Garbarino et al.,2012; Kian et al.,2017)。弄岗15 hm2 样地位于保护区的核心区,人为活动很少,因此,林隙聚集分布格局的形成原因应该和生境异质性密切相关。
本研究采用Worldview-3高空间分辨率遥感影像结合地面调查获取喀斯特季节性雨林林隙信息。研究结果显示,通过高空间分辨率遥感数据获取喀斯特森林林隙特征是可行的。高空间分辨率遥感在其它森林类型林隙信息提取时也被证明是可行的方法(Garbarino et al.,2012; Hobi et al.,2015; 毛学刚等,2018)。虽然目前一些新的研究手段,如无人机激光雷达(LiDAR)等技术可以获得更精細的林隙信息(Vepakomma et al.,2008; 毛学刚等,2017),但成本较高,而且在一些地形复杂的区域,无人机的应用也会受到限制。因此,对喀斯特森林林隙研究,高空间分辨率遥感目前还是一种折中和有效的方法。
4.2 林隙与树木的空间关系
从弄岗喀斯特季节性雨林林隙与树木的空间关联分析结果来看,不同树木对林隙的形成作用是不一样的。大树与林隙在空间上存在显著的正关联,而小树和枯立木与林隙在空间上表现出无关联,说明大树的分布与林隙的形成密切关联。 树木的死亡是林隙形成的原因(Muscolo et al.,2014),从弄岗喀斯特季节性雨林的研究结果来看,树木的枯立死亡并不会导致林隙的形成。这与Senécal et al.(2018)的研究结果一致。树木枯立死亡是一个渐进的过程,在树木逐渐枯死过程中,会增加林下层光资源的可利用度从而促进下层树木的生长,而在其完全死亡时,枯立木原来占据的林冠空隙已经被其它树木填充,因此树木的枯死通常并不会产生林隙。小树一般高度较低,冠幅也较小,通常处于林下层,因此小树对林隙的形成作用并不明显。森林中树木的高度通常与胸径呈正相关关系,并且具有更大的冠幅(Song et al.,2010),在其它森林中,大树的突然死亡(如风灾导致的大树掘根、折干等)已被证明是林隙形成的重要原因(Kian et al.,2017; 龙翠玲等,2005; 温远光等,2014)。喀斯特环境土壤少而岩石裸露率高,树木的根系较浅(龙翠玲等,2005),大树通常较高,在大风、大雨等外力作用下更易倒伏,弄岗喀斯特季节性雨林样地林隙与大树显著的空间正关联应该也与此有关。
4.3 林隙数量分布、基本特征的影响因子
不同生态因子对弄岗喀斯特季节性雨林林隙的数量分布、面积和周长等特征的影响并不相同。凹凸度对林隙数量分布有显著负影响;干旱度指数对林隙数量分布和林隙周长特征均有负影响;海拔对林隙大小和周长均有负影响;坡向对林隙大小有正影响;岩石裸露度和地形湿润度指数对林隙周长有负影响;而丰富度对林隙数量分布和林隙大小均有正影响,而多度对林隙周长为正影响。Goulamoussène et al.(2017)对热带森林的研究认为不同环境因子对林隙大小的作用并不相同,他们发现坡度越小的位置林隙的面积越大。但本研究中未发现坡度对林隙数量分布、大小和周长等特征存在显著的关系,原因可能与不同森林树种分布的影响因素不同有关。
喀斯特地区生境特殊性主要体现在两方面,即水平空间存在高度异质性,同时垂直剖面存在多层次性(郭柯等,2011)。弄岗喀斯特季节性雨林样地为典型喀斯特地形,光照、水分、土壤条件、岩石裸露率等环境因子会从峰顶至洼地发生剧烈变化(王斌等,2014)。地形变化对树种的分布具有两方面的影响,包括直接影响和间接影响。首先,是坡度、坡向、海拔等的直接影响;其次,由于地形因子变化引起光、温、水及土壤等资源再分配的间接影响,从峰顶至洼地随海拔逐渐降低,土壤水分含量迅速增加,土壤厚度逐渐增加而岩石裸露率逐渐减少,同时光照也逐渐减少(王斌等,2014)。地形变化耦合环境因子的急剧变化会影响树种的空间格局(黄甫昭等,2014),最终影响到林隙的分布和特征。受光、温、水、土壤条件和岩石裸露等因素的影响,在弄岗喀斯特季节性雨林样地相对海拔较高位置主要分布耐旱、耐贫瘠、喜光的树种,如清香木(Pistacia weinmannifolia)、黄梨木(Boniodendron minius)、毛叶铁榄(Sinosideroxylon pedunculatum)、山榄叶柿(Diospyros siderophylla)等, 这些树种主要为小乔木和灌木。在样地相对海拔较低地区分布的主要为耐涝、耐荫树种,如中国无忧花(Saraca declinata)、苹婆(Sterculia monosperma)、广西牡荆(Vitex kwangsiensis)、劲直刺桐(Erythrina stricta)、对叶榕(Ficus hispida)等,这些树种以高大乔木和小乔木为主;而在峰顶和洼地之间的中海拔地带,主要分布的是高大乔木树种为主,如蚬木(Excentrodendron hsienmu)、闭花木(Cleistanthus sumatranus)、网脉核果木(Drypetes perreticulata)等种类(黄甫昭等,2014)。总体来看,峰顶很少有DBH≥20cm的大树分布,而在中低海拔部位DBH≥20cm的大树分布较多(王斌等,2014)。从树木空间分布的规律可以推测大林隙主要在低海拔部位、而小林隙应该在高海拔部位。从广义线性模型分析的结果也可以比较清楚的发现这一规律,即海拔越低的位置林隙的大小和周长越大,岩石裸露度越低的位置林隙周长越大。同时,从模型拟合的结果也发现,物种多样性和多度等生物因子也对林隙的分布和特征有显著的影响,这应该主要与树种之间的相互作用有关。因此,弄岗喀斯特季节性雨林林隙的空间分布与特征存在着地形分异特征,是喀斯特独特的地质地貌和生境条件与喀斯特森林自身生物学特性综合作用的结果,是植物与环境协同进化的体现。
参考文献:
AL EXANDER HD, MACK MC, 2017. Gap regeneration within mature deciduous forests of Interior Alaska: Implications for future forest change [J]. For Ecol Manag, 396(2017):35-43.
BADDELEY A, DIGGLE PJ, HARDEGEN A, et al., 2014. On tests of spatial pattern based on simulation envelopes [J]. Ecol Monogr, 84(3): 477-489.
BROKAW NVL, 1982. The definition of treefall gap and its effect on measures of forest dynamics [J]. Biotropica, 14(2): 158-160.
BURNHAM KP,ANDERSON DR, 2002. Model selection and inference: A practical information-theoretic approach [M]. New York: Springer: 149-199.
GARBARINO M, BORGOGNO MONDINO E, LINGUAE, et al., 2012. Gap disturbances and regeneration patterns in a Bosnian old-growth forest: A multispectral remote sensing and ground-based approach [J]. Ann For Sci, 69(5): 617-625.
GOULAMOUSSNE Y, BEDEAU C, DESCROIX L, et al., 2017. Environmental control of natural gap size distribution in tropical forests [J]. Biogeosciences, 14(2): 1-18.
GUO K, LIU CC, DONG M, 2011. Ecological adaptation of plants and control of rocky-desertification on karst region of Southwest China [J]. Chin J Plant Ecol, 35(10): 991-999. [郭柯, 刘长成, 董鸣, 2011. 我国西南喀斯特植物生态适应性与石漠化治理 [J]. 植物生态学报, 35(10): 991-999.]
GUO YL, WANG B, XIANG WS, et al., 2016. Responses of spatial pattern of woody plants’ basal area to topographic factors in a tropical karst seasonal rainforest in Nonggang, Guangxi, southern China [J]. Biodivers Sci, 24(1): 30-39. [郭屹立, 王斌, 向悟生, 等, 2016. 喀斯特季节性雨林木本植物胸高断面积分布格局及其对地形因子的响应 [J]. 生物多样性, 24(1): 30-39.]
HOBI ML, GINZLER C, COMMARMOT B, et al., 2015. Gap pattern of the largest primeval beech forest of Europe revealed by remote sensing [J]. Ecosphere, 6(5): t15-t76.
HU LL, LI JS, WU XP, et al., 2010. Rewiews on methods of measuring geometric chracteristics of forest gaps involving gap size, gap shape, and the height of canopy trees surrounding the gap [J]. Acta Ecol Sin, 30(7): 1911-1919. [胡理乐, 李俊生, 吴晓莆, 等, 2010. 林窗幾何特征的测定方法 [J]. 生态学报, 30(7): 1911-1919.]
HUANG FZ, WANG B, DING T, et al., 2014. Numerical classification of associations in a northern tropical karst seasonal rain forest and the relationships of these associations with environmental factors [J]. Biodivers Sci, 22(2): 157-166. [黄甫昭, 王斌, 丁涛, 等, 2014. 弄岗北热带喀斯特季节性雨林群丛数量分类及与环境的关系 [J]. 生物多样性, 22(2): 157-166.]
HUBBELL SP, FOSTER RB,O'BRIEN ST, et al., 1999. Light gap disturbances, recruitment limitation, and tree diversity in a neotropical forest [J]. Science, 283(5401): 554-557.
KATHKE S, BRUELHEID EH, 2010. Gap dynamics in a near-natural spruce forest at Mt. Brocken, Germany [J]. For Ecol Manag, 259(3): 624-632.
KIAN S, TABARI M, ESMAILZADEHO, et al., 2017. Gap characteristics and disturbance regime in an intact hyrcanian oriental beech forest, iran [J]. Austr J For Sci, 134(4):323-346.
LIU SC, HUA WJ, DUANWB, et al., 2013. Gap characteristics in the mixed broad-leaved Korean pine forest in Xiaoxingan Mountains [J]. Acta Ecol Sin, 33(17): 5234-5244. [刘少冲, 王敬华, 段文标, 等, 2013. 小兴安岭阔叶红松混交林林隙特征 [J]. 生态学报, 33(17): 5234-5244.]
LONG CL, YU SX, WEI LM, et al., 2005. Disturbance regimes and the characteristics of gaps in Maolan Karst Forest, Guizhou Province [J]. Sci Silv Sin, 41(4): 13-19. [龙翠玲, 余世孝, 魏鲁明, 等, 2005. 茂兰喀斯特森林干扰状况与林隙特征 [J]. 林业科学, 41(4): 13-19.]
LOOSMORE NB, FORD ED, 2006. Statistical inference using the g or k point pattern spatial statistics [J]. Ecology, 87(8): 1925-1931.
MAO XG, DU ZH, LIU JQ, et al., 2018. Object-oriented segmentation and classification of forest gap based on QuickBird remote sensing image [J]. Chin J Appl Ecol, 29(1): 1-10. [毛學刚, 杜子涵, 刘家倩, 等, 2018. 基于面向对象的QuickBird遥感影像林隙分割与分类 [J]. 应用生态学报, 29(1): 1-10.]
MAO XG, HOU JY, BAI XF, et al., 2017. Multiscale forest gap segmentation and object-oriented classification based on DOM and LiDAR [J]. T Chin Soc Agric Mach, 48(9): 152- 159. [毛学刚, 侯吉宇, 白雪峰, 等, 2017. 基于DOM及LiDAR的多尺度分割与面向对象林隙分类 [J]. 农业机械学报, 48(9): 152-159.]
MUSCOLO A, BAGNATO S, SIDARI M, et al., 2014. A review of the roles of forest canopy gaps [J]. J For Res, 25(4):725- 736.
R CORE TEAM D. R: A language and environment for statistical computing [EB/OL]. URL https://www.R-project.org/.
RUNKLE JR, 1982. Patterns of disturbance in some Old-Growth mesic forests of eastern north america [J]. Ecology, 63(5):1533-1546.
SCHLIEMANN SA, BOCKHEIM JG, 2011. Methods for studying treefall gaps: A review [J]. For Ecol Manag, 261(7): 1143-1151.
SENCAL J, DOYON F, MESSIERC, 2018. Tree death not resulting in gap creation: An investigation of canopy dynamics of northern temperate deciduous forests [J]. Remote Sens-Basel, 10(1): 121.
SHEN ZH, LI DX, WANG GF, 2001. Studies on the gap disturbance of the mountain mixed forests of evergreen and deciduous broad-leaved species at mt. Dalaoling in the three gorges Ⅰ. The essential characteristics of the gaps [J]. Acta Phytoecol Sin, 25(3): 276-282. [沈泽昊, 李道兴, 王功芳, 2001. 三峡大老岭山地常绿落叶阔叶混交林林隙干扰研究I林隙基本特征 [J]. 植物生态学报, 25(3): 276-282.]
SONG CH,DICKINSON MB, SU LH, et al., 2010. Estimating average tree crown size using spatial information from Ikonos and QuickBird images: Across-sensor and across-site comparisons [J]. Remote Sens Environ, 114(5): 1099-1107.
VEPAKOMMA U, ST-ONGE B, KNEESHAWD, 2008. Spatially explicit characterization of boreal forest gap dynamics using multi-temporal lidar data [J]. Remote Sens Environ, 112(5): 2326-2340.
WANG B, HUANG YS, LI XK, et al., 2014. Species composition and spatial distribution of the 15 ha northern tropical karst seasonal rain forest dynamic plot in Nonggang of Guangxi, Southern China [J]. Biodivers Sci, 22(2): 141-156. [王斌, 黄俞淞, 李先琨, 等, 2014. 弄岗北热带喀斯特季节性雨林15 ha监测样地的树种组成与空间分布 [J]. 生物多样性, 22(2): 141-156.]
WANG B, XIANG WS, DING T, et al., 2014. Spatial distribution of standing dead trees abundance and its impact factors in the karst seasonal rain forest, Nonggang, southern China [J]. Chin Sci Bull, 59(35): 3479-3490. [王斌, 向悟生, 丁涛, 等, 2014. 弄岗喀斯特季节性雨林枯立木多度的空间分布及影响因子 [J]. 科学通报, 59(35): 3479-3490.]
WEN YG, LIN JY, ZHU HG, et al., 2014. Gaps and their natural disturbance regimes in the south subtropical montane evergreen broadleaved forest [J]. Guangxi Sci, 21(5): 447-453. [溫远光, 林建勇, 朱宏光, 等, 2014. 南亚热带山地常绿阔叶林林隙及其自然干扰特征研究 [J]. 广西科学, 21(5): 447-453.]
WIEGAND T, MOLONEY KA, 2004. Rings, circles, and null-models for point pattern analysis in ecology [J]. Oikos, 104(2): 209-229.
WU QG, WU FZ, YANG WQ, et al., 2013. Characteristics of gaps and disturbance regimes of the alpine fir forest in Western Sichuan [J]. Chin J Appl Environ Biol, 19(6): 922-928. [吴庆贵, 吴福忠, 杨万勤, 等, 2013. 川西高山森林林隙特征及干扰状况 [J]. 应用与环境生物学报, 19(6): 922-928.]
XIANG WS, LI XK, WANG B, 2017. A study of mapping forest monitoring plot in rugged topography of South China [J]. Guangxi Sci, 24(3): 219-224. [向悟生, 李先琨, 王斌, 2017. 中国南方复杂地形森林样地的测量标定方法 [J]. 广西科学, 24(3): 219-224.]
YAN SJ, HONG W, WU CZ, et al., 2004. Gaps and their natural disturbance characteristics in mid-subtropical evergreen broad-leaved forest in Wanmulin [J]. Chin J Appl Ecol, 15(7): 1126-1130. [闫淑君, 洪伟, 吴承祯, 等, 2004. 中亚热带常绿阔叶林林隙及其自然干扰特征的研究 [J]. 应用生态学报, 15(7): 1126-1130.]
ZANG RG, YANG YC, 1999. Gaps and their natural disturbance regimes in the tropical montane rain forest of Hainan Island [J]. Sci Silv Sin, 35(1): 1-8. [臧润国, 杨彦承, 1999. 海南岛热带山地雨林林隙及其自然干扰特征 [J]. 林业科学, 35(1): 1-8.]
