免疫共沉淀法测定14-3-3ε蛋白与Cdc25B的相互作用
2019-08-06李瑞林
李瑞林,孟 峻,刘 儒
(内蒙古医科大学第一临床医学院,内蒙古医科大学附属医院检验科,呼和浩特 010050;*通讯作者,E-mail:nmfrank@163.com)
小鼠卵母细胞减数分裂是一特殊的阻滞过程,因卵母细胞发育需要,减数分裂会停滞在生发泡期(germinal vesicle,GV),即G2期末。由生发泡期到生发泡破裂期(germinal vesicle breakdown,GVBD)是卵母细胞第一次减数分裂恢复的过程,相当于有丝分裂G2/M期的过渡[1,2]。关于小鼠卵母细胞减数分裂G2期阻滞的机制研究已成为当前国内外的研究重点。目前一些研究者认为蛋白激酶A(protein kinase A,PKA)/14-3-3ε/细胞分裂周期蛋白25B(cell division cycle 25B,Cdc25B)/成熟促进因子(maturation promoting factor,MPF)通路在卵母细胞减数分裂过程及G2期阻滞中具有调节作用,但关于Cdc25B蛋白和14-3-3ε蛋白是否存在相互作用及其作用位点尚不清楚。而免疫共沉淀利用抗原抗体结合的高特异性,已成为研究蛋白与蛋白相互作用的经典方法[3],本研究采用免疫共沉淀法研究Cdc25B蛋白和14-3-3ε蛋白的相互作用及其具体的作用位点,为小鼠卵母细胞减数分裂G2期阻滞的机制研究提供实验依据。
1 材料与方法
1.1 细胞株及组织
HEK293细胞购自中国科学院细胞库。小鼠肝组织经昆明雄性6-8周小鼠解剖后获得,由内蒙古大学提供。
1.2 主要试剂
克隆载体pGEM-T vector(Promega公司,美国),表达载体pcDNA3.1-ZEO(Invitrogen公司,美国),pEGFP C3载体由复旦大学刘喻博士惠赠,pBSK-CDC25B-WT、pBSK-CDC25B-S321A、pBSK-CDC25B-S321D由内蒙古医科大学微免研究室保存,DH5α感受态菌株(上海泽生生物技术有限公司),质粒DNA纯化试剂盒(Promega,美国),DNA快速纯化/回收试剂盒(上海泽生生物技术有限公司),牛血清白蛋白(BSA,Fraction V,盐城赛宝生物科技有限公司),fuGENG6转染试剂(Roche Molecular Biochemicals,德国),免疫共沉淀试剂盒(上海生工生物有限公司)。
1.3 主要仪器
基因扩增仪(Biometra,德国),凝胶自动成像仪(GDS8000BIO-RAD,美国),紫外分光光度仪(PⅥ UNIcAM SPECTRONIC,英国),CO2培养箱CBll5(WTB-binder,德国)。
1.4 pcDNA3.1-ZEO-HA-14-3-3ε表达载体的构建
用Trizol提取小鼠肝组织(30 mg)RNA,设计14-3-3epsilon引物(NM-009536.4):5′-GACCGTGCCTGCAGGTTGC-3′和5′-CTTGCCAGTGTGGCCGGAGA-3′,预计产物长度545 bp,RT-PCR扩增,PCR产物经回收纯化,将回收纯化的HA-14-3-3ε克隆到pGEM-T载体上,将正确的HA-14-3-3ε阳性克隆与pcDNA3.1-ZEO(+)载体用EcoR Ⅰ酶切后连接,构建成pcDNA3.1-ZEO-HA-14-3-3ε表达载体,经凝胶电泳鉴定此重组载体是否插入正确。
1.5 pEGFP-Cdc25B表达载体的构建
将pBSK-CDC25B-WT、pBSK-CDC25B-S321D、pBSK-CDC25B-S321A三种重组质粒和载体pEGFP-C3分别用BamH Ⅰ和XhoⅠ双酶切,反应体系在37 ℃水浴环境中反应3 h。酶切反应后,用1.5%琼脂糖凝胶电泳检测酶切产物,回收CDC25B-WT、CDC25B-S321D、CDC25B-S321A、pEGFP-C3的cDNA片段,通过OD260 nm判断其回收产量,用于载体重组。将回收片段进行连接反应,取2 μl连接产物转化感受态细胞E.coliJM109,按常规转化方法进行,涂板过夜。挑取单菌落在含Kana(+)的培养基上培养12 h,后用质粒提取试剂盒提取质粒,取2 μl质粒用XhoⅠ和BamH双酶切鉴定。选择酶切片段为1 800 bp的阳性克隆保存,并行序列测定,构建成pEGFP-Cdc25B-WT,pEGFP-Cdc25B-Ser321A,pEGFP-Cdc25B-Ser321D表达载体。
1.6 HEK293细胞培养和转染
HEK293细胞培养于DMEM培养液(含100 U/ml青霉素,10% FBS,10 μg/ml链霉素),于37 ℃、5%CO2、98%饱和湿度培养箱。用fuGENG6转染剂转染HEK293细胞,转染过程严格按照fuGENG6转染说明书进行。转染组分为四组,分别为5μg pEGFP-Cdc25B-WT和5 μg pcDNA3.1-ZEO-HA-14-3-3ε(Cdc25B-WT+14-3-3ε组);5 μg pEGFP-Cdc25B-Ser321A和5 μg pcDNA3.1-ZEO-HA-14-3-3ε(Cdc25B-Ser321A+14-3-3ε组);5 μg pEGFP-Cdc25B-Ser321D和5 μg pcDNA3.1-ZEO-HA-14-3-3ε(Cdc25B-Ser321D+14-3-3ε组);5 μg pEGFP-Cdc25B-Vector和5 μg pcDNA3.1-ZEO-HA-14-3-3ε(Cdc25B-vector+14-3-3ε组)。
1.7 免疫共沉淀法测定14-3-3ε蛋白与Cdc25B S321位点的结合
加Protein G-琼脂糖到含过滤片的柱子中,PBS洗涤琼脂糖,低速离心洗四次。HEK293细胞转染48 h后,4 ℃ 800 r/min离心3 min,收集细胞。吸弃上清并用预冷的PBS洗涤细胞两次,完全去除PBS并保留细胞沉淀,用玻璃匀浆器匀浆30-50次,间隔1 min,重复3次。然后12 000 r/min离心5 min,收集上清液作为细胞裂解液。在新的微量离心管中加入细胞裂解物,PMSF和鼠单克隆抗HA抗体,4 ℃孵育1 h至过夜,将细胞裂解液转移到装有洗涤过Protein G-琼脂糖柱中,孵育2 h至过夜。将每个柱子中插入转移到2 ml的新微量离心管中。离心去掉上清,用IP缓冲液洗涤沉淀琼脂糖珠子5-6次,加入上样缓冲液,轻轻混合珠子,密封柱子。在95 ℃加热5 min,用薄纸或滤纸吸干盖子周围的水。将柱子插入到新的微量离心管中,以12 000g离心30 s,获得免疫沉淀物。
用Western blotting分别检测细胞裂解液和免疫沉淀物中Cdc25B和14-3-3ε蛋白的表达,经细胞裂解、提取蛋白、制胶、电泳、转膜、封闭及抗体孵育,通过化学发光及成像获得免疫印迹结果图,如免疫沉淀物中抗HA抗体和抗GFP抗体均存在(2条带均显影,判定为二者结合),表明Cdc25B和14-3-3ε相互结合,此实验中使用鼠单克隆抗HA抗体1 ∶800用于检测外源性表达的14-3-3ε蛋白,抗-GFP抗体1 ∶8 000用于检测外源性表达的Cdc25B蛋白。
2 结果
2.1 重组质粒pcDNA3.1-ZEO-HA-14-3-3ε
使pcDNA3.1-ZEO-HA-14-3-3εcDNA片段亚克隆至pmax-FP-Red-C表达载体中,用EcoR Ⅰ酶切重组质粒,经凝胶电泳鉴定可见4 800 bp的载体带及800 bp的目的带(见图1),说明亚克隆成功。
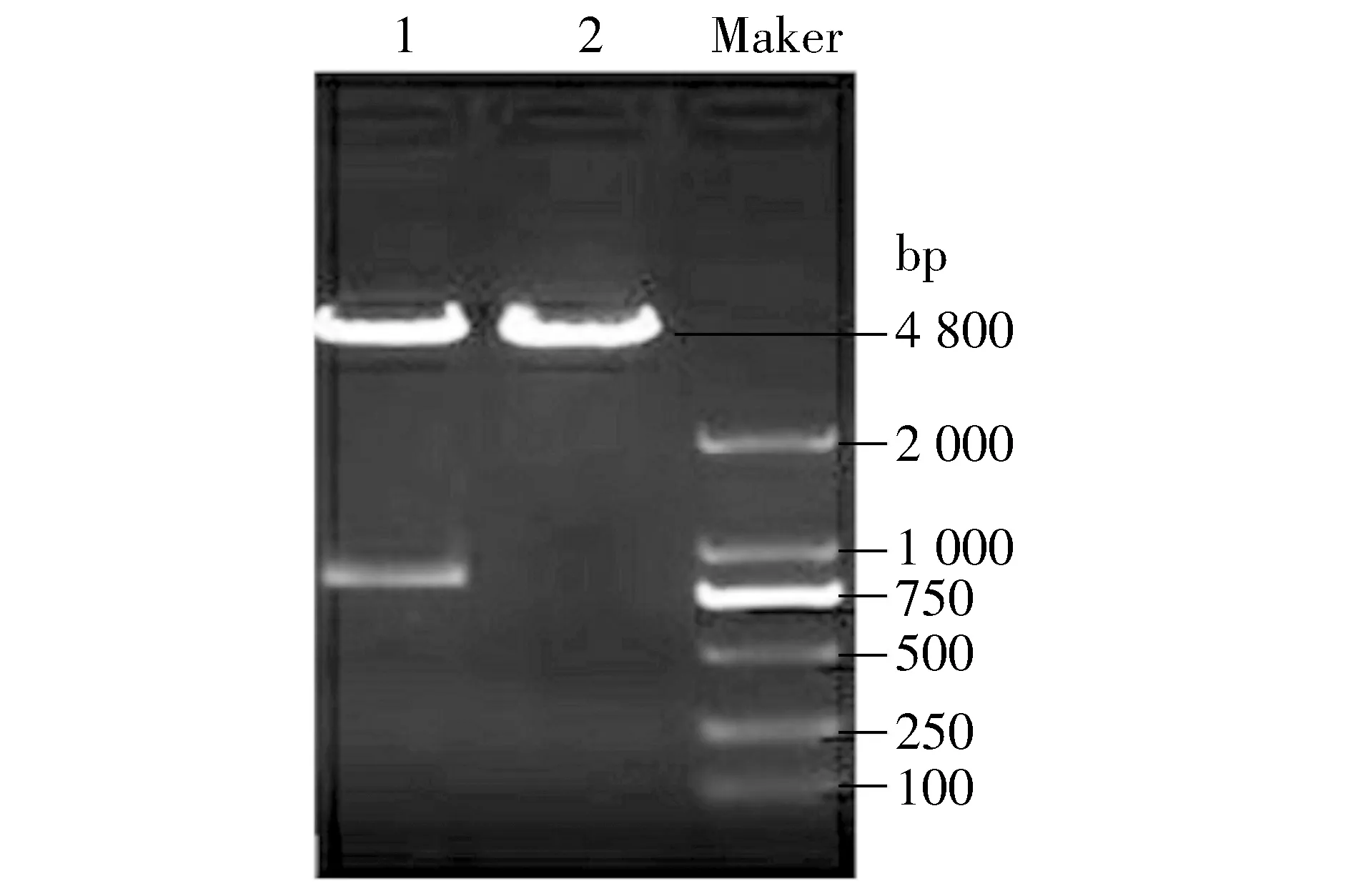
Marker.DL2000 DNA;1.pmax-FP-Red-HA-14-3-3ε;2.pmax-FP-Red-C图1 重组质粒EcoR Ⅰ单酶切鉴定结果Figure 1 Identification of pmax-FP-Red-HA-14-3-3ε plasmid after digested by EcoR Ⅰ
2.2 三种重组质粒的测序及鉴定
双酶切pEGFP-Cdc25B-Ser321A、pEGFP-Cdc25B-WT、pEGFP-Cdc25B-Ser321D三种重组表达载体,经琼脂糖凝胶电泳后可见约5 000 bp的载体条带及1 800 bp的目的条带(见图2),与预期结果一致,经上海泽生测序证明重组体构建成功。
2.3 14-3-3ε和Cdc25B的免疫共沉淀结果
在各组细胞裂解中,除了Cdc25B-Vector组外,Cdc25B和14-3-3ε蛋白均有表达(见图3);而在免疫沉淀物中,Cdc25B-WT和Cdc25B-S321D均与14-3-3ε蛋白有共沉淀(见图3),表明两者结合;而Cdc25BS321A与14-3-3ε无共沉淀(见图3),表明两者未结合。

Marker.DL15000 DNA;1.pEGFP-Cdc25B-WT;2.pEGFP-Cdc25B-Ser321A;3.pEGFP-Cdc25B-Ser321D图2 重组质粒双酶切鉴定结果Figure 2 Double enzyme digestion of recombinant pEGFP-Cdc25B plasmids

图3 免疫共沉淀测定14-3-3ε和Cdc25B的相互作用Figure 3 Determination of the interaction of 14-3-3ε and Cdc25B by co-immunoprecipitation
3 讨论
Cdc25B是一种丝氨酸/苏氨酸双特异性磷酸化酶,是细胞分裂的启动者,在细胞周期中具有重要的调控作用[4,5]。而14-3-3ε是一高度保守的酸性蛋白,广泛表达于真核生物体内,以穿梭胞核与胞质的方式调节与之结合的靶蛋白的功能[6,7]。目前诸多研究表明,此蛋白可调节Cdc25B蛋白的活性和亚细胞定位,两蛋白的结合对细胞分裂进程有重要的调控作用[8]。早期的研究显示,卵母细胞G2/M期的转换需要激活Cdc2/cyclinB复合物(MPF),Wee1B/Myt1激酶能磷酸化Cdc2的Thr14和Tyr15,使MPF失活,卵母细胞停滞于G2期。而Cdc25B能使Cdc2的Thr14和Tyr15位点去磷酸化,激活MPF,启动减数分裂的恢复[9,10]。Yang等[11]的研究发现在爪蟾卵母细胞中,PKA磷酸化Cdc25B的287位丝氨酸(S287),14-3-3ε与磷酸化的Cdc25B S287的结合使得Cdc25B隔离于细胞质不能进入细胞核,不能激活核内的MPF,从而使卵母细胞不能继续分裂,停滞于第一次减数分裂的G2期。
Astuti等[12]研究表明,PKA可磷酸化Cdc25B的321位丝氨酸,引起GV期阻滞,而突变体S321A能完全解除PKA引起的GV期阻滞,而突变体S149不能解除GV期阻滞,因此Cdc25B的321位丝氨酸在卵母细胞减数分裂中有重要作用。Cui等[13]的研究表明,14-3-3ε通过与Cdc25B结合的方式阻滞小鼠受精卵G2/M期的转换。Uchida等[14]研究表明:在HEK293细胞中,14-3-3ε通过与Cdc25B的309位丝氨酸结合的方式控制Cdc25B的细胞内定位。那么小鼠卵母细胞是否也存在14-3-3ε,且与Cdc25B在减数分裂恢复过程中是否有相互作用。
本课题组之前对鼠卵母细胞的研究证明了14-3-3ε的存在,且在GV和GVBD期14-3-3ε蛋白的表达量没有变化[15]。14-3-3ε siRNA使GV到GVBD的发生率为36%,而非小干扰RNA的GV到GVBD的发生率为0,说明14-3-3ε siRNA可促进小鼠卵母细胞的G2/M转换[16]。为证明14-3-3ε和Cdc25B在调控卵母细胞减数分裂中是否存在联系,本课题组之前的研究实验初次证明小鼠卵母细胞的GV期,Cdc25B蛋白和14-3-3ε蛋白共定位于细胞质;在GVBD前,一部分Cdc25B从细胞质进入细胞核;而在GVBD后,Cdc25B和14-3-3ε均匀地分布于整个细胞[17]。此结果表明在GV期Cdc25B和14-3-3ε两蛋白可能具有相互作用,可能是因Cdc25B蛋白和14-3-3ε蛋白的结合使有丝分裂的Cdc25B隔离于胞质而不能入核,不能继续进行减数分裂,使卵母细胞发生GV期阻滞。
为了验证14-3-3ε与Cdc25B确实能结合或有相互作用,本课题组进行了免疫共沉淀实验。此实验证明了14-3-3ε与Cdc25B确实能结合,野生型Cdc25B能和14-3-3ε结合;而当Cdc25B的321位丝氨酸突变为丙氨酸时,Cdc25B与14-3-3ε不再结合;当Cdc25B的321位丝氨酸突变为天冬氨酸时,Cdc25B仍然能结合14-3-3ε,说明S321D具有模拟磷酸化的作用。提示14-3-3ε只能与磷酸化的S321结合,而不与未被磷酸化的S321A结合,Cdc25B的321位丝氨酸是14-3-3ε和Cdc25B结合的特异性位点。14-3-3ε和Cdc25B的321位磷酸化的丝氨酸结合,使Cdc25B隔离于胞浆,卵母细胞停滞于G2期,当14-3-3ε和Cdc25B的结合取消时,Cdc25B又可入核,激活MPF,启动减数分裂。
关于14-3-3ε和Cdc25B的结合取消的分子机制将是以后的研究方向。此次研究初次证明在鼠卵母细胞中,S321是14-3-3ε和Cdc25B结合的特异性位点,PKA磷酸化Cdc25B的321位丝氨酸,磷酸化的321位丝氨酸与14-3-3ε结合引起G2期阻滞,S321是一重要的细胞周期调控靶点。我们可以通过使S321位点的磷酸化和去磷酸化来人为地调控细胞周期进程。通过此位点对细胞周期调控的研究成果可应用于农牧业,提高动物的繁殖数量,满足人类肉食的需求;也可应用于肿瘤的治疗,使肿瘤细胞停滞在G2期,抑制肿瘤的生长;还可用于人类体外辅助生殖技术,缩短受精卵的体外培育时间,尽早植入人体子宫,减少外界因素对受精卵的影响。近年来,不孕不育的患者逐渐增多,人类生殖辅助技术水平虽有很大的进展,然而,这些新技术的实施都需要有受精能力及发育潜力极好的卵母细胞。因此,深入研究卵母细胞成熟调控的机制至关重要。
