条斑紫菜(Pyropia yezoensis)绿斑病病原菌的分离鉴定*
2019-08-05牟宗娟杨慧超茅云翔阎永伟莫照兰
李 杰 牟宗娟, 杨慧超,3 茅云翔 阎永伟 莫照兰,3
条斑紫菜()绿斑病病原菌的分离鉴定*
李 杰1牟宗娟1,2杨慧超1,3茅云翔2阎永伟1莫照兰1,3①
(1. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室农业农村部海水养殖病害防治重点实验室 中国水产科学研究院黄海水产研究所 青岛 266071;2. 中国海洋大学海洋生命学院 青岛 266003; 3. 上海海洋大学水产与生命学院 上海 201306)
绿斑病(Green spot disease)是一种常见的海区栽培条斑紫菜()病害,在整个紫菜栽培期间都可能发生,以每年11~12月份最为严重,主要出现在幼叶期和成叶期。首先,在叶状体上出现红色或淡红色小斑,而后逐渐转变为绿色,病斑继续扩展,在叶状体表面形成若干孔洞,后期几乎整个藻体变绿。本研究对日照地区患绿斑病的条斑紫菜进行病原菌分离纯化,得到5株优势菌(编号为Y1~Y5),人工回感实验结果显示,Y1可以引起健康条斑紫菜发生绿斑病。对Y1进行了生理生化检测、16S rRNA、和基因序列分析,确定病原菌为海洋假交替单胞菌()。对绿斑病的发病进程进行了观察,并检测了培养温度、海水比重和养殖密度等环境因子对绿斑病发生的影响,结果显示,高温和高密度养殖会加速绿斑病病情的发展,海水比重为1.022时,绿斑病发病较严重。本研究确定了一株引起条斑紫菜绿斑病的病原菌,并分析了部分理化因子对感染的影响,为条斑紫菜绿斑病的防控提供了基础数据。
条斑紫菜;叶状体;绿斑病;海洋假交替单胞菌
条斑紫菜()是大型潮间带藻类,自然分布较广,自辽宁至浙江,适合紫菜生长的岩礁海域均可发现其踪迹(曾呈奎等, 1985)。条斑紫菜的栽培主要集中在中国、日本和韩国,其味鲜美,含有丰富的必需氨基酸、维生素和矿物质,是中国两大紫菜栽培品种之一。
随着条斑紫菜栽培规模的扩大,密集栽培技术的发展,其病害问题表现越来越突出,严重影响紫菜的产量和商品质量。据不完全统计,条斑紫菜育苗和栽培过程中现已发现病害16种,已报道和发现的包括绿斑病、赤腐病、拟油壶菌病、丝状细菌症、癌肿病、缩曲症、黄斑病和色圈病等多种紫菜病害(Nakao, 1972; Fujita, 1973; Lin, 1984; 陈秋萍等, 1991; 马家海, 1992、1996; 马家海等, 1999; 闫咏等, 2002)。紫菜的病害是由多方面原因引起的,致病机理非常复杂,微生物是不可忽视的重要因素之一。本研究针对山东省日照地区条斑紫菜叶状体绿斑病,开展了病原分离鉴定、致病性检测、发病进程及环境因子影响等一系列研究,旨在探明绿斑病的病因和病理,并为该病的防治提供参考数据。
1 材料与方法
1.1 实验用条斑紫菜
患绿斑病条斑紫菜于2010年11月采自山东省日照岚山海区条斑紫菜养殖场,水温为18℃;健康条斑紫菜采自山东省青岛汇泉湾以及团岛,养殖于PES海藻培养基中(马家海, 1996)。
1.2 细菌分离
选择症状明显的条斑紫菜叶状体,无菌海水漂洗数次,剪取病斑藻段、匀浆,用无菌海水梯度稀释匀浆液,涂布2216E海水培养基于28℃培养5 d,挑取菌落形态一致的优势菌,采用15%的甘油保种、保存于–80℃冰箱备用。
1.3 人工感染
挑选颜色正常的健康条斑紫菜叶状体,除去藻体表面杂质,置于0.7% KI溶液中浸泡15 min(方文雅, 2010),用灭菌海水冲洗后用于回感实验。
将分离得到的疑似病原菌在2216E液体培养基中培养过夜,离心收集菌体,用灭菌PES-海水冲洗3次后重悬。吸取一定量菌悬液加到灭菌PES-海水中,制备细菌终浓度为108和107CFU/ml的PES-海水,以灭菌PES-海水作为阴性对照。每个浓度感染5片紫菜叶状体,培养温度为15℃,光照周期L︰D=12︰12 (h),光强为62.5 μmol/m2·s (5000 lx)。每个感染组和对照组设置3个平行,观察紫菜叶状体的病理发展过程,实验结束时,选择具有病症的紫菜进行细菌分离鉴定,以确证病原菌。
1.4 病原菌鉴定
采用革兰氏染色法对细菌进行染色,镜检;采用Vitek 2-GN和API-ID32E生理生化检测试剂盒(生物梅里埃公司, 法国),参照产品使用说明书对病原菌进行生理生化特征测定;利用16S rRNA基因引物27F (5¢-AGAGTTTGATCCTGGCTCAG-3¢)和1492R (5¢- CTACGGCTACCTTGTTACGA-3¢),PCR扩增Y1 16S rRNA基因序列;根据NCBI中20余种假交替单胞菌()的和基因序列,通过BioEdit软件对比序列分析,选择保守区设计2对简并引物F (5¢-GTGTATYTGTCGGTTTGGC-3¢)、R(5¢-TNARYTCTTTWGAYARHGCC-3¢)和F (5¢-CCDYTRRTRCARGTDTCDGGYGC-3¢)、R (5¢-AYARDCGCATHGGCATSACNAC-3¢),PCR扩增Y1和基因片段,片段长度分别为1301和1124 bp,PCR反应条件:95℃预变性5 min, 95℃变性30 s,55℃退火30 s,72℃延伸90 s,35个循环,72℃延伸7 min。将扩增得到的PCR产物送至上海桑尼生物科技有限公司进行测序,获得的序列信息在GenBank中用Blast(http://blast.ncbi.nlm. nih.gov/)进行同源性比较,利用MEGA 5.0软件,采用邻位相连法(Neighbor-Joining)构建系统进化树(Bootstrap=1000)。
1.5 环境因子对感染的影响
培养温度分别设置为8℃、15℃、18℃和22℃,检测温度对绿斑病发病的影响;条斑紫菜密度分别设置为 0.05 g/100 ml、0.1 g/100 ml和0.2 g/100 ml,检测紫菜养殖密度对绿斑病发病的影响;用无菌超纯水调节海水比重为1.019、1.022和1.025(相对应的海水盐度分别为24.5、29.1和32.7),检测海水比重对绿斑病发病的影响。每个实验组设置3个平行,除变化的环境因子,其他培养条件:温度为18℃、密度为0.1 g/100 ml,海水比重为1.022。实验组分别加入终浓度为108CFU/ml的病原菌,对照组在同样条件下不加入病原菌培养。观察叶片的变化,并计算感染 1周后病烂部分面积所占整个叶片面积的百分比和培养体系中细菌数量的变化。
2 结果与分析
2.1 绿斑病及细菌分离
条斑紫菜绿斑病发病部位多位于叶状体的中部和边缘,初期发病紫菜叶片出现红色小点,扩大后逐渐转变成灰绿色,在叶状体表面形成若干孔洞,叶片腐烂变绿(图1A)。在2216E培养基上共分离得到5株优势菌,分别编号为Y1、Y2、Y3、Y4和Y5。
2.2 人工感染
菌株Y1感染紫菜可以产生明显绿斑病症状 (图1B),Y2、Y3、Y4和Y5各个浓度梯度感染的条斑紫菜均未观察到发病。Y1菌株108CFU/ml感染组紫菜在培养2 d后出现病症,107CFU/ml感染组在10 d出现病症。显微镜观察可见,在病斑形成位置,最初单个紫菜细胞发生明显变化,原生质浓缩,颜色由正常的紫褐色变为紫红色,细胞形状变成不规则状(图2A),进一步周围的细胞渐渐失去细胞正常结构(图2B),后期以这些病变细胞为中心,呈放射状蔓延(图2C),边缘的细胞颜色渐渐变绿并最终形成肉眼可见的病斑(图2D)。条斑紫菜叶状体病变部分与健康部分区别明显,且病变部分细胞由于颜色不同可以清晰地分为多个不同状态的细胞层(图2E、图2F)。

图1 海区自然发病条斑紫菜(A)和人工感染发病条斑紫菜(B)
2.3 病原菌的鉴定
革兰氏染色结果显示,Y1属于革兰氏阴性短杆菌。Vitek 2 GN鉴定结果显示,其丙氨酸-苯丙氨酸-脯氨酸芳胺酶(APPA)、β-半乳糖苷酶(β-GAL)、脂酶(LIP)、酪氨酸芳胺酶(TyrA)、α-半乳糖苷酶(α-GAL)、磷酸酶(PHOS)以及ELLM反应为阳性,其他均为阴性。经ID-32E鉴定,α-葡萄糖(α-GLU)、α-麦芽糖苷酶(α-MAL)和L-天门冬素芳胺酶(ASPA)反应为阳性,其他反应为阴性。ATB系统分析结果显示,Y1为假交替单胞菌()。
对Y1的16S rRNA、和基因进行扩增测序和GenBank比对,并构建系统发育进化树,结果显示,Y1的基因序列与海洋假交替单胞菌()相似度均为99%~100%,且系统进化树与海洋假交替单胞菌聚为一支(图3),表明Y1与海洋假交替单胞菌进化关系最为相近,确定引起此次条斑紫菜绿斑病的病原为海洋假交替单胞菌。
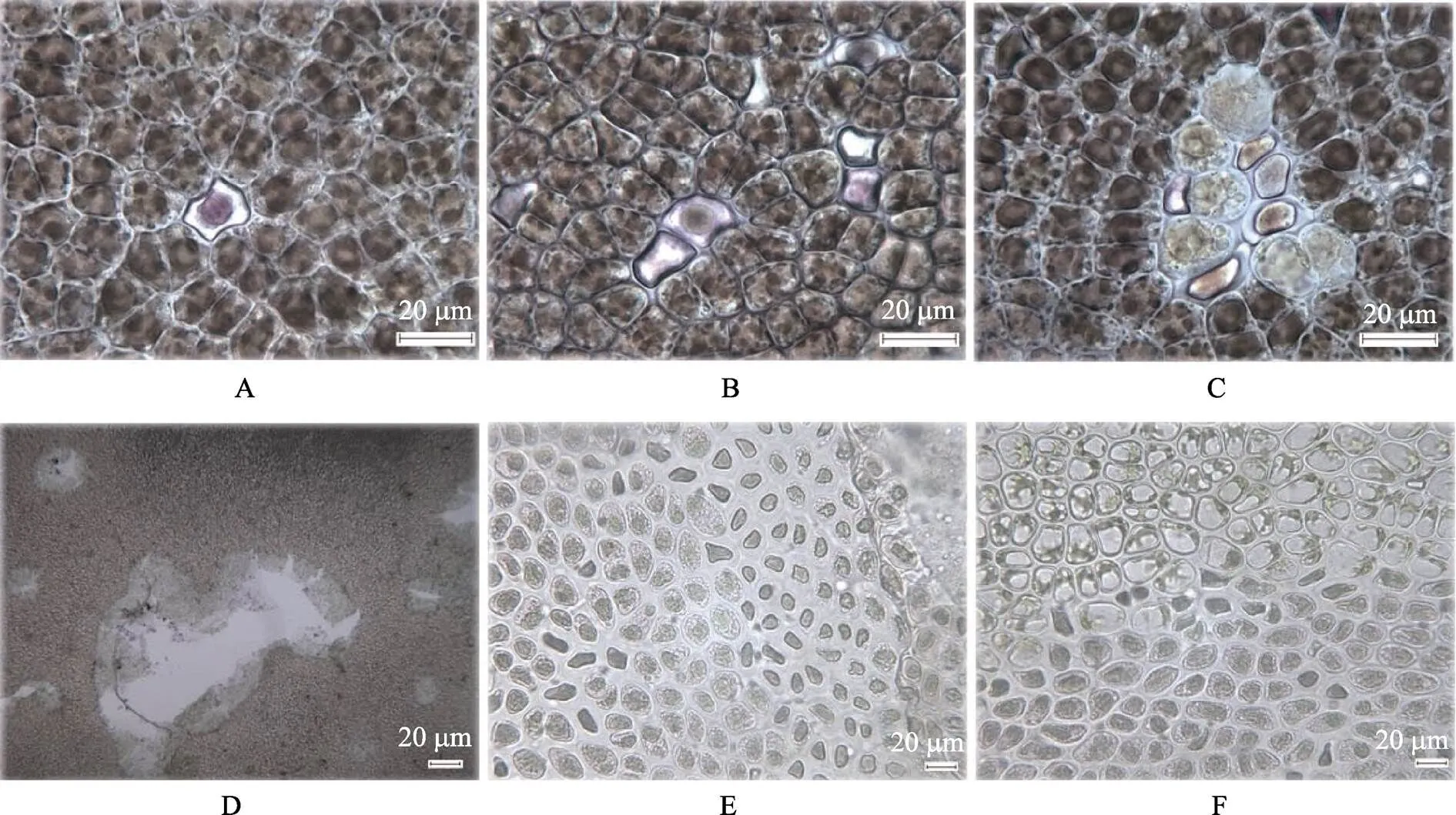
图2 人工感染条斑紫菜
A:单个紫菜细胞浓缩变色;B:多个紫菜细胞浓缩变色;C:初期形成的绿斑;D:叶片破碎形成孔洞;E和F:绿斑和正常细胞交界处细胞质颗粒降解的细胞层
A: Concentration and discoloration ofsingle cell; B: Concentration and discoloration ofcells; C: Early stage of green spot disease; D: Holes on laver; E and F: Cytoplasmic granules degradation at the junction of diseased and normal cells
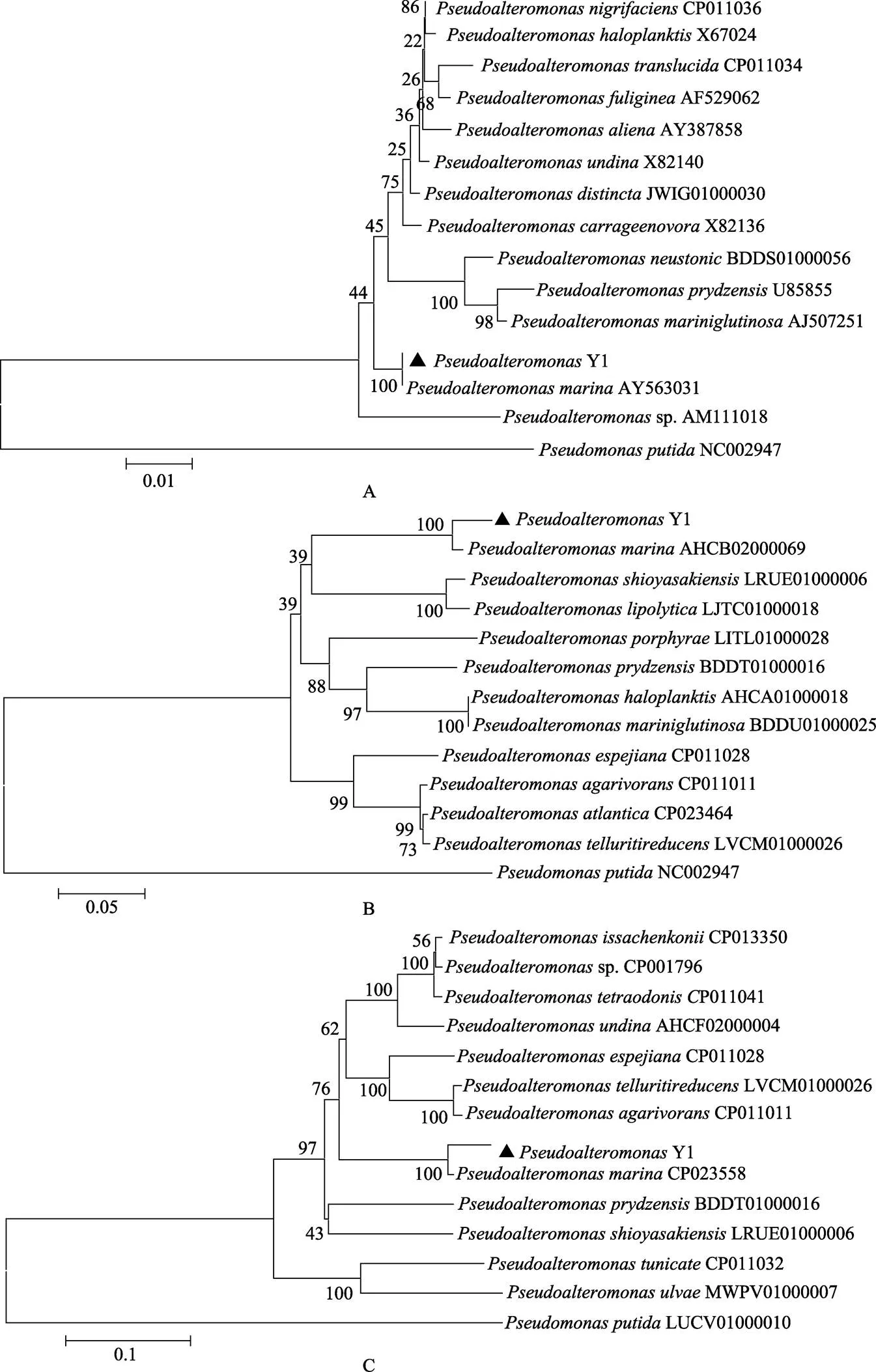
图3 Y1系统进化发育分析
A: 基于16S rRNA基因序列构建的系统发育进化树; B: 基于序列构建的系统发育进化树; C: 基于序列构建的系统发育进化树
A: Phylogenetic tree constructed for isolates based on 16S rRNA gene sequences; B: Phylogenetic tree constructed for isolates based onsequences; C: Phylogenetic tree constructed for isolates based onsequences
2.4 环境因子对感染的影响
设置4个不同的感染温度,检测了温度对条斑紫菜绿斑病发病的影响。结果显示,在感染温度为18℃时,条斑紫菜发病最快,感染36 h后,可观察到红色斑点;感染温度为15℃和22℃时,感染48 h后出现症状;感染温度为8℃时发病较慢,感染72 h后出现1~2个很小的斑点。除22℃高温组,各组的对照组在整个实验过程中未观察到明显病烂症状。感染培养7 d后,除8℃组仅有几个斑点外,其余各实验组叶状体均可见多个病烂区,并伴有组织脱落。各实验组叶状体病烂部分面积所占总面积的比例如图4A所示,当感染温度为22℃时,叶状体病烂最严重,几乎看不到正常的叶片部分。
设置3个不同的养殖密度,检测了密度对条斑紫菜绿斑病发病的影响。低密度条件下病烂部分最少,随着养殖密度的增大,病烂程度有所加剧。感染72 h后,0.2 g/100 ml组出现多个明显的病斑,0.1 g/100 ml组仅出现1~2个病斑,0.05 g/100 ml组没有出现病斑。感染7 d后,各组都出现严重病烂。经计算,各实验组叶状体病烂部分面积所占总面积的比例如图4B所示,0.05 g/100 ml实验组病烂面积最小,约45%的叶片出现病烂;0.1 g/100 ml组与0.2 g/100 ml组病烂面积较大,超过60%的叶片出现病烂。
设置3个不同比重的海水,检测海水比重对条斑紫菜绿斑病发病的影响。正常海水比重组叶状体(1.022)在感染2 d后出现红色斑点,7 d后明显病烂;低海水比重组(1.019)和高海水比重组(1.025)分别在感染4 d和5 d之后,出现红色斑点,且发生病烂的面积较小。7 d后,各实验组叶状体病烂部分面积所占总面积的比例如图4C所示,1.019低海水比重组和1.025高海水比重组海水都能一定程度上延缓病情的发生进展。

图4 环境因子对条斑紫菜绿斑病发病的影响
3 讨论
绿斑病是紫菜栽培过程中的常见疾病,在整个紫菜栽培期都可能发生,但以每年11、12月份最为严重(丁怀宇, 2008)。绿斑病多发生于栽培密度较大的海区,尤其是在高温后极易发生,病变的部位不定,在条斑紫菜叶状体的梢部、中部、基部、中央和边缘部位均可出现病斑。绿斑病一旦发生,传染的速度极快,病斑可在短时间内连成一片,并迅速变绿,在适宜条件下,藻体1周即可全部溃烂,严重时,网帘上紫菜完全脱落。绿斑病会使紫菜表面粗糙、无光泽,影响其商品价值。2013~2014年,韩国舒川郡紫菜养殖场绿斑病爆发造成110万美元的损失,相当于总销售额的10.7%(Kim, 2014)。
1968年绿斑病在日本首次发现,其病原较为广泛,从患病紫菜中可以分离出微球菌(sp.)、假单胞菌(sp.)以及弧菌(sp.)(Nakao, 1972),中国已报道柠檬假交替单胞菌()引起的条斑紫菜绿斑病(闫咏等, 2002),本实验室在前期研究中发现弧菌也可以引起坛紫菜()绿斑病(韩晓娟等, 2015)。目前报道的绿斑病病原菌大部分具有较强的胞外酶活性,这些胞外酶导致的宿主损伤可能是引起紫菜叶片绿斑的重要原因。近年来研究发现,叶绿体病毒(Chloroplast virus)也可以引起条斑紫菜绿斑病的发生(Kim, 2016)。
绿斑病只是一种表观特征,多种因素造成的藻体破坏、藻红素溶出,均可呈现绿斑病症状。海水温度异常升高、降雨或有机质污染引起藻体代谢失常都可能引起紫菜绿斑病(Gachon, 2010),但环境胁迫条件下导致的附生微生物菌群失控是引起紫菜病烂的主要因素(Egan, 2016)。通过环境因子实验结果,发现高温、高密度养殖是引起紫菜绿斑病发生的主要因子,而海水比重的变化在一定程度上可以减缓病烂速度,这也与海区栽培情况相印证。每年11、12月份,海区紫菜进入快速生长期,此时网帘上紫菜密度较大,为致病菌提供了丰富的营养和良好的环境。在正常养殖条件下,条件致病菌数量一般维持在一定范围内,不具备致病力。当出现升温或降雨等环境变化,条件致病菌的致病性可能增强,造成紫菜绿斑病的发生。合理控制养殖密度,及时疏苗,密切注意水温等气象环境的变化,充足的干出以改变紫菜表面海水比重等方法可以有效抑制绿斑病的发生。
本研究通过人工回接感染实验、细菌生理生化和基因序列分析,发现海洋假交替单胞菌可以引起条斑紫菜的绿斑病,养殖密度、温度和海水比重等环境因子会影响绿斑病的发病进程,其具体感染和致病机制有待进一步深入研究。
Chen QP, Jiang ZX. A preliminary analysis of the causes of extensive rotting from diseases ofin Tong Tou prefecture. Journal of Zhejiang College of Fisheries, 1991, 10(2): 144–148 [陈秋萍, 江最新. 洞头县坛紫菜大面积病烂原因分析. 浙江水产学院学报, 1991, 10(2): 144–148]
Ding HY. Study on development process of green-spot disease in. Journal of Anhui Agricultural Sciences, 2008, 36(11): 4626–4628 [丁怀宇. 条斑紫菜绿斑病发病进程研究. 安徽农业科学, 2008, 36(11): 4626–4628]
Egan S, Gardiner M. Microbial dysbiosis: Rethinking disease in marine ecosystems. Frontiers in Microbiology, 2016, 7(505): 991
Fang WY. Preliminary study of the platform to study the interaction ofspp. and their phycospheres. Masterˊs Thesis of Ningbo University, 2010 [方文雅. 紫菜与藻际微生物互作研究平台的初步研究. 宁波大学硕士研究生学位论文, 2010]
Fujita Y. Maceration of laver frond by enzymes of bacteria causing green spot rotting-like deterioration. Bulletin of the Japanese Society of Scientific Fisheries, 1973, 39(8): 911–915
Gachon CM, Sime-Ngando T, Strittmatter M,Algal diseases: Spotlight on a black box. Trends in Plant Science, 2010, 15(11): 633–640
Han XJ, Mao YX, Li J,. Identification and pathogenicity of a pathogen causing green rot disease in.Journal of Fisheries of China, 2015, 39(11): 1721–1729 [韩晓娟, 茅云翔, 李杰,等一株引起坛紫菜绿斑病病原的分离鉴定及致病性研究. 水产学报, 2015, 39(11): 1721–1729]
Kim GH, Klochkova TA, Lee DJChloroplast virus causes green-spot disease in cultivatedof Korea. Algal Research, 2016, 17: 293–299
Kim GH, Moon KH, Kim JYA revaluation of algal diseases in Korean() sea farms and their economic impact. Algae, 2014, 29(4): 249–265
Lin DH, Lin YS. Studies on the green disease ofChang & Zheng caused by nitrogen deficiency. Hydrobiologia, 1984, 116–117(1): 453–455
Ma JH, Zhang LM, Ji CL,. Studies on crape disease of. Journal of Fishery Sciences of China, 1999, 6(2): 82–88 [马家海, 张礼明, 吉传礼等条斑紫菜缩曲症的研究. 中国水产科学, 1999, 6(2): 82–88]
Ma JH. A preliminary study on the red rot disease of. Journal of Shanghai Fisheries Univeristy, 1996, 5(1): 1–7 [马家海. 条斑紫菜赤腐病的初步研究. 上海水产大学学报, 1996, 5(1): 1–7]
Ma JH. An investigation of chytrid-disease inin the coastal water of south Jiangsu. Journal of Shanghai Fisheries Univeristy, 1992, 1(3–4): 185–188 [马家海. 江苏省南部沿海条斑紫菜壶状菌病的调查研究. 上海水产大学学报, 1992, 1(3–4): 185–188]
Nakao Y, Onohara T, Matsubara T,Bacteriological studies on diseases of cultured laver-I: Green spot rotting-like deterioration of laver frond by bacteria,. Nippon Suisan Gakkaishi, 1972, 38(6): 561–564
Yan Y, Ma JH, Xu P,., the causative agent of green-spot disease of. Journal of Fishery Sciences of China, 2002, 9(4): 353–358 [闫咏, 马家海, 许璞等1株引起条斑紫菜绿斑病的柠檬假交替单胞菌. 中国水产科学, 2002, 9(4): 353–358]
Zeng CK, Wang SJ, Liu SJ,. Marine algae culture science. Shanghai: Shanghai Scientific and Technical Publisher, 1985 [曾呈奎, 王素娟, 刘思俭. 海藻栽培学. 上海: 上海科学技术出版社, 1985]
Isolation and Identification the Pathogen ofGreen Spot Disease
LI Jie1, MOU Zongjuan1,2, YANG Huichao1,3, MAO Yunxiang2, YAN Yongwei1, MO Zhaolan1,3①
(1. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Key Laboratory of Maricultural Organism Disease Control, Ministry of Agriculture and Rural Affairs, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071; 2. College of Marine Life Sciences, Ocean University of China, Qingdao 266003; 3. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306)
Green spot disease is one of the major disease considerations in the farming of. Green spot disease could be found in all the periods of thallus, and outbreaksusually occur in November and December. At the start of infection, small light red spots could be found on the thallus, and then the spots gradually turned green and expanded quickly. Holes from lesions formed later in the surface of thallus. At last, the entire thallus became green and fell off. In this research, bacteria strains were isolated from.with green spot disease farmed in Rizhao. Experimental infection showed that the strain Y1 could cause.green spot disease. Biochemical characterization and genes analysis of 16S rRNA,andindicated that the pathogen Y1 was. Influence of environmental factors on the outbreak of disease were also characterized by experimental infection with different temperature, stocking density and gravity of sea water. The results showed that higher temperature and stocking density will accelerate the spread of the disease, but gravity of sea water within a certain range did not affect the occurrence of the disease. In this study, we described the green spot disease caused by., which provides information for disease control in.cultivation.
; Thallus; Green spot disease;
MO Zhaolan, E-mail: mozl@ysfri.ac.cn
S946.2
A
2095-9869(2019)04-0140-07
10.19663/j.issn2095-9869.20180710002
* 现代农业产业技术体系(CARS-50)、国家高技术研究发展计划(2012AA10A406)、国家科技基础条件平台建设运行项目(2018DKA30470)和鳌山科技创新计划(2015ASKJ02)共同资助[This work was supported by China Agriculture Research System (CARS-50), National High-Tech R&D Program (2012AA10A406), National Science and Technology Infrastructure Platform Construction (2018DKA30470), and Aoshan Technology Innovation Program (2015ASKJ02)]
李 杰,E-mail: lijie@ysfri.ac.cn
莫照兰,研究员,E-mail: mozl@ysfri.ac.cn
2018-07-10,
2018-07-30
李杰, 牟宗娟, 杨慧超, 茅云翔, 阎永伟, 莫照兰. 条斑紫菜()绿斑病病原菌的分离鉴定. 渔业科学进展, 2019, 40(4): 140–146
Li J, Mou ZJ, Yang HC, Mao YX, Yan YW, Mo ZL. Isolation and identification the pathogen ofgreen spot disease. Progress in Fishery Sciences, 2019, 40(4): 140–146
(编辑 马璀艳)
