饥饿及复投喂对金钱鱼肠型脂肪酸结合蛋白基因表达的影响
2019-07-20刘建业陈华谱江东能吴天利田昌绪李广丽朱春华邓思平
刘建业 陈华谱 江东能 吴天利 田昌绪 李广丽 朱春华 邓思平
(1. 广东海洋大学水产学院, 湛江 524088; 2. 广东省名特优鱼类生殖调控与繁育工程技术研究中心, 湛江 524088;3. 海洋生态与养殖环境湛江市重点实验室, 湛江 524088)
脂肪酸结合蛋白(Fatty acid-binding proteins,Fabp)属于脂质结合蛋白超家族成员, 这一类蛋白质的分子量较低, 广泛存在于脊椎动物和非脊椎动物的多种组织细胞的胞浆中, 是一种胞质蛋白质,对细胞内长链脂肪酸的摄取、转运及代谢调节发挥着重要作用, 主要参与脂肪酸的运输, 可将脂肪酸从细胞膜运送到甘油三酯和磷脂合成的位点[1]。
目前发现的Fabp蛋白有18种类型, 都是以分离或鉴定的第一种组织命名, 有肠型、心型、脂肪型、肝脏型、脑型、回肠型、上皮细胞型和髓磷脂型等[1—3]。肠型脂肪酸结合蛋白(Intestines fatty acid binding protein, Ifabp)是首先从肠黏膜细胞液中分离出来的脂肪酸结合蛋白, 拥有一个单配体结合位点[4], 因此能够特异结合未酯化的游离长链脂肪酸并参与其运输[5]。目前在大黄鱼(Larimichthys crocea)[6]、建鲤(Cyprinus carpiovar. jian)[7]、斑马鱼(Danio rerio)[8]、大西洋鲑(Salmo salar)[9]等少数鱼类克隆得到ifabp基因序列, 饥饿及复投喂对大黄鱼ifabp基因表达具有显著影响, 但在金钱鱼中, 肠型脂肪酸结合蛋白基因(ifabp)的分离和在脂肪代谢中的作用尚未见报道。
金钱鱼(Scatophagus argus)隶属鲈形目(Perciformes), 金钱鱼科(Scatophagidae), 金钱鱼属(Scatophagus)广盐性鱼类, 且杂食性, 广泛分布于温带和亚热带及热带地区, 具有极强的环境适应性和抗逆性, 是一种具有观赏和食用价值的名贵海水经济鱼类。目前, 金钱鱼的研究主要集中于生物学[10]、繁殖生物学[11]、温度和鱼油对其卵巢发育的影响[12,13]和雌激素受体在卵黄生成中的功能[14]。有关金钱鱼生长调控的研究仅见生长激素在雌雄间的二态性表达[15], 不同性别养殖金钱鱼生长性能及消化酶活性的比较[16]、饥饿复投喂对spexin[17]和igfbp基因表达的影响[18]。为阐明Ifabp在金钱鱼脂肪代谢中的作用, 本文克隆并分析了金钱鱼2种脂肪酸结合蛋白基因(ssifabp2a和ssifabp2b), 半定量反转录-聚合酶链反应(Semi-quantitative reverse transcription PCR, RT-PCR)检测了这2种基因在组织中的分布情况, 实时荧光定量PCR(Real-time fluorescence quantitative PCR, qPCR)检测了饥饿-复投喂处理后, 这2种基因在肠和肝脏中的表达变化, 研究结果可为探讨ifabp基因在脂肪代谢中的作用提供理论依据。
1 材料与方法
1.1 材料
基因克隆和组织表达实验所采用的金钱鱼(体长: 15—19 cm; 体重: 100—180 g)购自广东省湛江市霞山区水产品批发市场, 取雌和雄各3尾用于组织表达。饥饿和复投喂所采用的实验鱼为海捕金钱鱼鱼苗, 并在广东海洋大学东海岛海洋生物研究基地喂养至1龄左右的金钱鱼(体重: 52—60 g)。所有金钱鱼均采用MS-222麻醉后于冰上取所需组织后立即投入液氮保存, 随后转于-80℃低温冰箱保存用于总RNA的提取和反转录成cDNA。
1.2 方法
总RNA提取及反转录采用TRIzol(Invitrogen)试剂提取总RNA, 实验操作根据试剂盒说明书的要求进行, cDNA利用PrimeScriptTMRT reagent Kit with gDNA Eraser反转录酶(TaKaRa)合成。
基因克隆和序列分析利用本实验室建立的金钱鱼肝脏转录组数据库, 筛选出与大黄鱼ifabpb基因相似度较高的unigene片段, 利用软件primer premier 6.0设计引物对(ifabp2a-f: ctgcagag tcgtccagttcac、ifabp2a-r: cttttggctttattaaaaccc)和(ifabp2b-f: ttcttcatcgccagcccagcca、ifabp2b-r: tcttt ccaaggtgcataagact)分别用于包含ssifabp2a和ssifabp2b开放阅读框的序列扩增。PCR反应的条件为: 94℃ 30s, 57℃ 30s, 72℃ 60s, 33个循环。扩增产物经12 mg/mL的琼脂糖凝胶电泳, 切下目的片段, 用琼脂糖回收试剂盒回收、纯化目的片段。将回收产物按3∶1的比例与pMD19-T (TaKaRa)载体连接。连接产物5 μL用于转化Escherichia coliDH5α,通过菌落PCR获得的阳性克隆送生工生物工程(上海)股份有限公司测序。所有引物均由生工生物工程(上海)股份有限公司合成。采用DNAstar软件比较金钱鱼与其他脊椎动物Ifabp氨基酸序列同源性。利用MEGA6.0邻位相联法 (Neighbor-joining),重复1000次, gap处理为缺失, 基于Ifabp氨基酸序列构建各物种系统进化树。
组织表达以下丘脑、垂体、鳃、肝脏、心脏、脾脏、肾脏、胃、肠、性腺和肌肉11个组织反转录出的cDNA为模板, 以获得的ssifabp2a和ssifabp2b序列设计特异引物对(qifabp2a-f1: cgaaact gatcagcagaacttg、qifabp2a-r1: aaaaccctgtgaaattcaaaa ctt)和(qifabp2b-f1: catttgtggttttctcgtaagtg、qifabp2br1: gacatggtgctccctcagtt)RT-PCR分别用于检测ssifabp2a和ssifabp2b在组织中的表达。两对特异引物扩增的长度分别为173和198 bp。β-actin基因做为内参, 内参基因β-actin引物对(β-actinF: gagaggtt ccgttgcccagag,β-actinR: cagacagcacagtgttggcgt)的扩增长度为145 bp。
饥饿及复投喂对肠、肝脏中ssifabp2a与ssifabp2b基因表达的影响用于饥饿和复投喂的金钱鱼随机分为2d对照组、2d饥饿组、7d对照组、7d饥饿组和复投喂组, 每组2个网箱, 每个网箱喂养4尾鱼。对照组为正常投喂2d和7d的实验鱼;饥饿组为饥饿2d和7d的实验鱼; 复投组为饥饿7d后再次投喂, 并在投喂3h后取样的实验鱼。实验鱼经2周驯化后用于饥饿和复投喂处理, 每天定量(饵料按体重的2%投喂)、定时(上午9:00)投喂(饵料购于中国悦群海洋生物研究开发公司)。每次取样为定时取样(上午10:00), 均取2个网箱的全部8尾鱼的肝脏和肠用于总RNA的提取和反转录成cDNA, 并用于基因表达检测。以β-actin基因作为内参, 以与组织表达相同的引物对为引物, 采用ABI 7500荧光定量PCR仪, SYBR Green Realtime PCR Master Mix(TOYOBO, Japan)试剂盒, 利用qPCR检测饥饿及复投喂后金钱鱼的肠和肝脏中ifabp2a和ifabp2b基因表达变化。qPCR反应程序为: 95℃预变性1min;95℃变性15s, 58℃退火15s, 72℃延伸45s并收集荧光信号, 共40个循环; 然后进行融解曲线分析。基于标准曲线方法,β-actin、ssifabp2a与ssifabp2b基因的扩增效率分别为1.06、0.95和0.97。
数据分析每个实验样品重复2次, 根据qPCR所得的Ct值取平均值, 运用2-ΔΔCt计算ssifabp2a和ssifabp2b基因相对表达量。所有数据均以平均值±标准误(x±SE)表示。利用SPSS 19.0软件进行单因素方差分析(ANOVA), 采用Duncan’s多重比较法进行各组之间ifabp2a和ifabp2b基因的表达差异, 当P<0.05时, 各组间具有显著性差异。
2 结果
2.1 ssifabp2a与ssifabp2b基因序列分析
从金钱鱼肝脏转录组数据库筛选出与大黄鱼ifabpb基因相似度较高的unigene片段, 经过亚克隆测序验证后, 得到金钱鱼ssifabp2a(GenBank登录号: MH118291)与ssifabp2b基因(GenBank登录号:MH396540)的cDNA序列。ssifabp2a与ssifabp2b两者开放阅读框长度都为399 bp, 且都编码133个氨基酸。同源性比较发现, Ifabp2a和Ifabp2b与其他硬骨鱼类肠型脂肪酸结合蛋白同源性较高; ssIfabp2a与其他硬骨鱼类Ifabp2a、Ifabp或IfabpX1的同源性为(78.8%—87.9%), 与月光鱼(Xiphophorus maculatus)IfabpX1、欧洲鲈(Dicentrarchus labrax) Ifabp、塞内加尔鳎(Solea senegalensis) Ifabp2a和尖吻鲈(Lates calcarifer) Ifabp的同源性分别为78.8%、87.9%、79.5%和85.6%; ssIfabp2b与其他硬骨鱼类Ifabp2b或Ifabp-like的同源性为(79.5%—87.9%), 与月光鱼Ifabp-like、欧洲鲈Ifabp2、塞内加尔鳎Ifabp2b和尖吻鲈Ifabp2b或Ifabp-like分别为79.5%、84.7%、87.9%和87.1%; ssIfabp2a与ssIfabp2b的同源性为73.5%。聚类结果表明, ssIfabp2a与其他硬骨鱼类Ifabp2a、Ifabp或IfabpX1聚为一类, ssIfabp2b则与其他鱼类的Ifabp2b或Ifabp-like聚为一类(图 1)。
2.2 ssifabp2a与ssifabp2b在组织中的表达
RT-PCR检测ssifabp2a与ssifabp2b在各个组织中的表达情况发现: 在雄鱼中,ssifabp2a在肠中表达最强, 在肾、肝脏、精巢、胃、下丘脑、心和鳃中有微弱表达, 在脑垂体、脾、肌肉没有检测到表达;ssifabp2b也在肠组织中表达最强, 在下丘脑、鳃、心、肝脏、胃、精巢和肌肉中有微弱表达, 在脑垂体、脾和肾中没有检测到表达。在雌鱼中,ssifabp2a在胃中表达最强, 在肾、肝脏和下丘脑组织表达较弱, 在其他组织中有微弱表达, 脑垂体中没有检测到表达; 与ssifabp2a表达情况不同,ssifabp2b在下丘脑、卵巢、心脏和肠中表达较强, 其他组织中有微弱表达, 鳃中没有检测到表达(图 2)。
2.3 饥饿及复投喂对肠、肝脏中ssifabp2a与ssifabp2b基因表达的影响
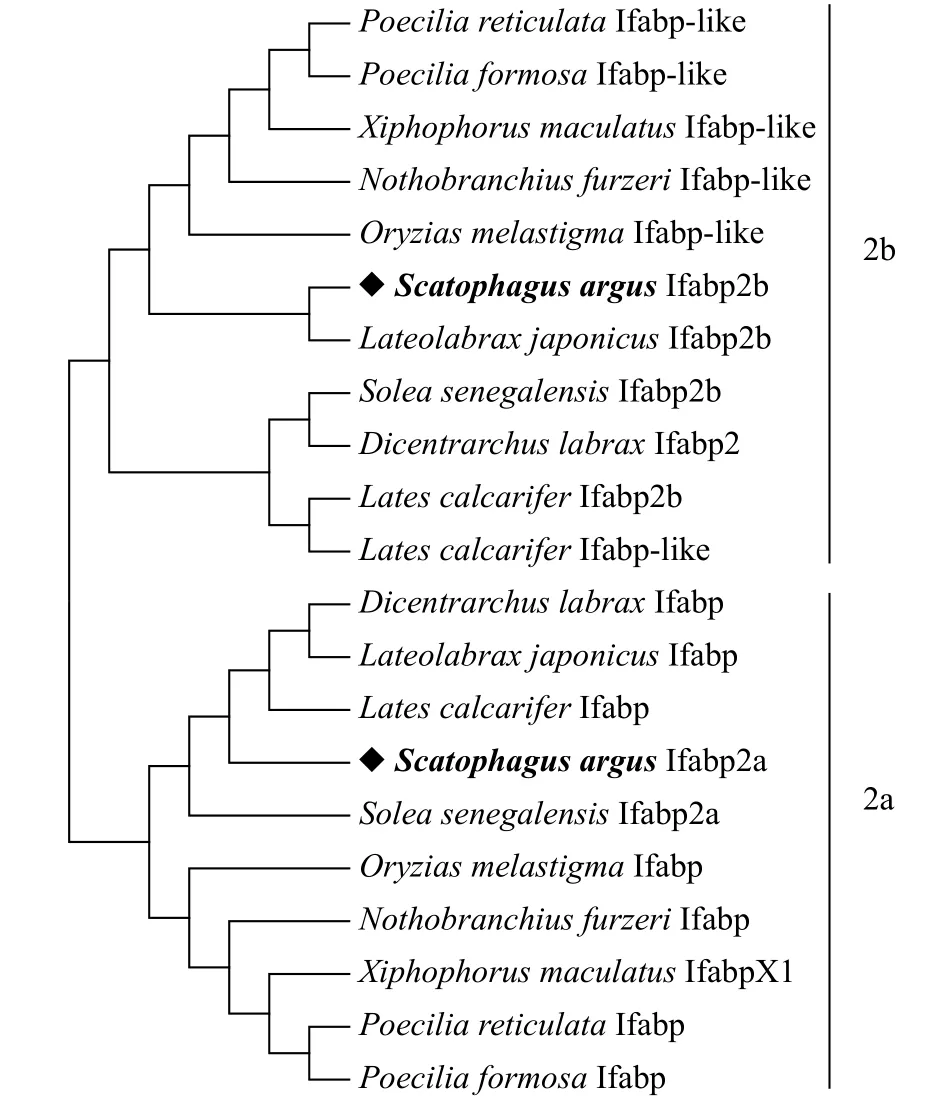
图 1 基于NJ法构建的金钱鱼和其他硬骨鱼类的Ifabp系统进化树Fig. 1 Phylogenetic tree of S. argus and other teleost Ifabp based on Neighbor-Joining method using MEGA6.0本进化树采用MEGA6.0软件(邻位相连法)构建, 系统树中结点处数值代表1000次评估的自举检验置信度。各物种的Ifabp蛋白在GenBank登录号分别为: 花鳉(Poecilia reticulata) Ifabplike: XP_008433909.1; 秀美花鳉(Poecilia formosa) Ifabp-like:XP_007562596.1; 月光鱼(Xiphophorus maculatus) Ifabp-like:XP_005813454.1; 虾虎鱼(Nothobranchius furzeri) Ifabp-like:XP_015802465.1; 青鳉(Oryzias melastigma) Ifabp-like:XP_024144067.1; 花鲈(Lateolabrax japonicus) Ifabp2b:APG58402.1; 塞内加尔鳎(Solea senegalensis) Ifabp2b: ALC 78633.1; 欧洲鲈(Dicentrarchus labrax) Ifabp2: ATJ34039.1; 尖吻鲈(Lates calcarifer) Ifabp2b: AGL33438.1; 尖吻鲈Ifabp-like:XP_018543508.1; 欧洲鲈Ifabp: AHK05999.1; 花鲈(Lateolabrax japonicus) Ifabp: AOW69620.1; 尖吻鲈Ifabp: XP_018551190.1;塞内加尔鳎Ifabp2a: ALC78632.1; 海洋青鳉(Oryzias melastigma) Ifabp: XP_024145490.1; 虾虎鱼(Nothobranchius furzeri) Ifabp: XP_015799257.1; 月光鱼IfabpX1: XP_005807197.1; 花鳉(Poecilia reticulata) Ifabp: XP_008409747.1;秀美花鳉(Poecilia formosa) Ifabp: XP_007551827.1Distances are used to construct the phylogenetic tree and bootstrap values are based on 1000 resampling replicates. The branch length scale in terms of genetic distance is indicated above the tree
QPCR结果发现, 饥饿及复投喂可显著影响ssi-fabp2a与ssifabp2b基因的表达。在饥饿2d后, 肠中ssifabp2a表达量显著降低,ssifabp2b无显著性变化;在饥饿7d后,ssifabp2a的表达量显著下降, 但ssifabp2b与对照组无显著差异; 与7d饥饿组相比较,在复投喂后,ssifabp2a恢复到正常水平,ssifabp2b的表达量显著上升(图 3、图 4)。在饥饿2d后, 肝脏中ssifabp2a表达量无显著变化, 而ssifabp2b的表达量显著升高; 在饥饿7d后, 肝脏中ssifabp2a和ssifabp2b的表达量均显著升高; 在复投喂后,ssifabp2a和ssifabp2b表达量恢复到正常水平(图 5、图 6)。
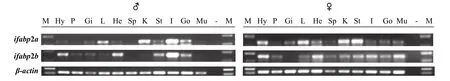
图 2 RT-PCR检测雄性(♂)和雌性(♀)金钱鱼组织中ssifabp2a与ssifabp2b的表达情况Fig. 2 Expression of ssifabp2a and ssifabp2b at tissues in male (♂) and female (♀) S. argus by RT-PCR M. DL2000 Marker; Hy. 下丘脑; P. 脑垂体; Gi. 鳃, L. 肝脏, He. 心脏, Sp. 脾脏, K. 肾脏, St. 胃, I. 肠, Go. 性腺, Mu. 肌肉, -. 空白; βactin为内参基因

图 3 饥饿和复投喂对金钱鱼肠中ifabp2a基因表达的影响Fig. 3 Effects of fast feeding and refeeding on the expression of ifabp2a in the intestine of S. argus字母不同表示同一时间各实验组之间存在显著性差异(P<0.05);下同Values with the same letters are not significantly different at the same time (P<0.05); the same applies below

图 4 饥饿和复投喂对金钱鱼肠中ifabp2b基因表达的影响Fig. 4 Effects of fast feeding and refeeding on the expression of ifabp2b in the intestine of S. argus

图 5 饥饿和复投喂对金钱鱼肝脏中ifabp2a基因表达的影响Fig. 5 Effects of fast feeding and refeeding on the expression of ifabp2a in the liver of S. argus
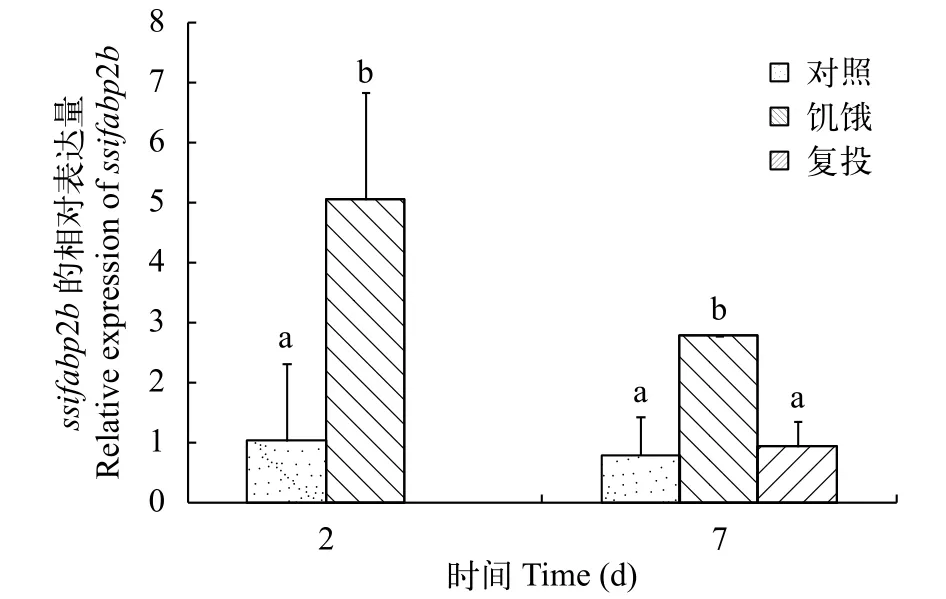
图 6 饥饿和复投喂对金钱鱼肝脏中ifabp2b基因表达的影响Fig. 6 Effects of fast feeding and refeeding on the expression of ifabp2b in the liver of S. argus
3 讨论
从NCBI数据库和现有文献中发现, 很多鱼类拥有2种类型的ifabp基因,ifabp基因命名方式主要有3种: 第一种是ifabp2a和ifabp2b, 如塞内加尔鳎等;第二种是ifabp和ifabp-like, 如花鳉等; 第三种是ifabp-like和ifabpX1, 如月光鱼等。本研究从金钱鱼肠组织中克隆得到了金钱鱼ssifabp2a和ssifabp2b基因的cDNA序列。对ssifabp2a和ssifabp2b基因进行序列分析和同源比对发现, Ifabp2a和Ifabp2b与其他硬骨鱼类肠型脂肪酸结合蛋白同源性较高。在构建的进化树中发现, ssifabp2a和其他硬骨鱼类的Ifabp2a、Ifabp和IfabpX1聚为一簇, ssifabp2b和其他硬骨鱼类的Ifabp2b和Ifabp-like聚为一簇。因而,ifabp2a、ifabp和ifabpX1可能属于命名规则不同的同一亚型的ifabp基因, 而ifabp2b和ifabp-like属于另一类ifabp亚型基因。根据序列分析结果, 本研究将克隆得到的2种金钱鱼肠型脂肪酸结合蛋白命名为ssifabp2a与ssifabp2b。
有研究表明,ifabp基因在鱼类的肠、脑、胃、肝脏和肌肉等组织中均有表达, 在肠中的表达量普遍最高[19,20]。与斑马鱼[8]、虹鳟(Oncorhynchus mykiss)[21]和建鲤[7]肠中表达量最高类似, 在金钱鱼雄鱼肠中的ssifabp2a和ssifabp2b表达量也最高。肠中ifabp的高表达表明, Ifabp在金钱鱼肠道脂肪酸的代谢过程中同样起重要作用。但在金钱鱼雌鱼肠中,ssifabp2b表达较强, 而ssifabp2a表达较弱, 造成这种现象的原因可能与金钱鱼雌鱼生长显著快于雄鱼有关[22]。此外, 与在斑马鱼肝脏、脑、精巢和肌肉中也检测到ifabp微弱表达基本一致, 雄性金钱鱼ssifabp2a在肾、肝脏、精巢、胃、下丘脑、心和鳃中有微弱表达,ssifabp2b在下丘脑、鳃、心、肝脏、胃、精巢和肌肉中微弱表达, 其他组织无表达。在雌鱼中,ssifabp2a在胃、肾、肝脏和下丘脑有较强表达, 在其他组织中有微弱表达, 脑垂体中没有检测到表达; 与ssifabp2a表达情况不同,ssifabp2b在下丘脑, 卵巢和心脏中有较强表达, 其他组织中有微弱表达, 鳃中没有检测到表达。造成这种组织差异表达的原因尚未明确。哺乳动物和鸟类不同类型的脂肪酸结合蛋白一般在单一组织中表达, 如人、鼠、鸡等Ifabp基因仅在肠中表达[23—25]。在金钱鱼中, 在多个组织中都可检测到ifabp基因的表达, 在其他鱼类中也同样发现ifabp基因在多个组织中表达, 如斑马鱼、大黄鱼、虹鳟和建鲤等, 根据表达模式的不同, 相比于哺乳动物和鸟类单一组织表达模式, 推测ifabp基因在鱼类中功能可能更为广泛。
在金钱鱼中发现, 金钱鱼饥饿处理2d和7d后,肠中ssifabp2a都显著下降, 在复投喂后,ssifabp2a的表达量显著升高, 但仍显著低于对照组; 在饥饿2d、7d过程中,ssifabp2b均无显著性变化, 复投喂后显著上升。在肝脏中, 饥饿2d时,ssifabp2a无显著变化,ssifabp2b显著升高, 在饥饿7d时,ssifabp2a和ssifabp2b均显著升高, 复投喂后均显著降低到对照组水平。在大黄鱼中, 饥饿对大黄鱼肠和肝中ifabpb基因表达呈现出先上升后下降的趋势, 与金钱鱼不同, 在复投喂后, 肠和肝脏中ifabpb表达量均显著升高。肠型脂肪酸结合蛋白(Ifabp)在动物体内的主要作用是调节脂肪酸的摄取和胞内转运。有大量的研究表明,ifabp基因的表达受到各种因素的影响, 如哺乳动物小鼠受年龄、饲料中脂肪含量、激素等[26,27], 非洲爪蟾蜍(Xenopus laevis)ifabp受甲状腺激素[28], 大西洋鲑(Salmo salar)受饲料脂肪含量[9], 大黄鱼受饥饿[6]等影响。本研究发现, 在金钱鱼ssifabp2a和ssifabp2b的表达同样受到饥饿的影响。已有研究表明, 调节脂代谢使体内能源平衡是一个复杂的过程, 此过程需要很多调控系统协调完成。在大西洋鲑的脂肪代谢相关研究发现, 脂肪酸作为信号分子结合到转录因子过氧化物酶体增殖物激活受体上并使其激活, 然后通过这个转录因子和Fabps启动子元件相互作用来诱导Fabps基因的转录和翻译[9,29]。尼罗罗非鱼的研究发现, 长期饥饿会导致组织中脂肪降解产生的脂肪酸被大量代谢, 用来产生能量, 当脂肪酸下降到一定浓度时会导致基因ifabp表达量显著降低[1,30]。在肝脏中, 饥饿2d时,ssifabp2a无显著变化,ssifabp2b显著升高, 在饥饿7d时,ssifabp2a和ssifabp2b均显著升高, 复投喂后均显著降低到对照组水平。相比于肠组织, 肝脏中的ssifabp2a和ssifabp2b表达量在饥饿2d和饥饿7d相比于对照组都是显著上升的。推测造成这种现象的原因可能与金钱鱼长期饥饿缺乏脂肪来源, 会导致大量氧化分解体内组织中脂肪酸等物质[30], 从而促进ifabp的表达, 所以肝脏中的ssifabp2a和ssifabp2b表达量在饥饿2d到饥饿7d都显著上升。这与瓦氏黄颡鱼饥饿30d肝中Ifabp基因表达量持续上升[20]和大黄鱼饥饿初期肝中的Ifabpb基因表达有显著性增加[6]的结果一致。在饥饿及复投喂处理中, 肠和肝中ssifabp2a和ssifabp2b的表达量都有显著变化, 表明两者都参与了金钱鱼的脂肪代谢过程, 两者的作用机制有待进一步研究。
