淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ通过BMP/Runx2/Osx信号通路促进大鼠骨髓间充质干细胞成骨分化的实验研究
2019-06-11訾慧范颖蒋宁谷丽艳董秀
訾慧 范颖 蒋宁 谷丽艳 董秀
辽宁中医药大学中医脏象理论及应用教育部重点实验室,辽宁 沈阳 110847
骨质疏松症是一种增龄性病变,随着人口老龄化,增龄导致骨质疏松症及其骨折的发病率逐年上升,并成为严重的医学和社会问题。中医认为“肾藏精生髓主骨”,肾主藏精,精生髓,髓养骨。中医认为“肾藏精生髓主骨”,肾主藏精,精生髓,髓养骨。肾中精气旺盛则骨就强壮,否则反之。“肾精”也担负成年组织修复的功能。如《灵枢·海论》云:“髓海有余,则轻劲多力;自过其度,髓海不足,则脑转耳鸣,胫酸眩冒,目无所见,懈怠安卧”。现代医学研究证明,干细胞是具有多向分化潜能的成体干细胞,以骨髓组织中含量最为丰富。骨髓间充质干细胞(bone marrow stromal cells,BMSCs)是一种多能间充质干细胞,在一定条件下可以向成骨方向分化,可能受多种信号通路调控,其中BMP/Runx2/OSX信号通路是与骨形成密切相关的通路,能促进成骨分化[1]。骨髓间充质干细胞的分化能力与分化方向体现了骨髓的功能,因此认为补“肾”可以提高骨髓间充质干细胞的分化能力[2-3]。
淫羊藿为中医临床补肾壮骨常用药材,淫羊藿苷为其主要有效成分,具有明显的促进骨形成、抑制骨吸收的活性[4-6]。淫羊藿苷口服给药后,在体内会产生多种代谢产物如淫羊藿素、淫羊藿次苷I、淫羊藿次苷Ⅱ等,且原型药物生物利用度较低[7]。贾晓斌等[8]发现淫羊藿苷经口给药发挥主要功效的物质基础主要为其代谢产物,翟远坤等[9]曾报道淫羊藿苷的代谢产物具有强于其本身的生物活性。笔者[10]前期实验结果也表明,淫羊藿次苷I与其原型药物淫羊藿苷相比,有更好的促进骨髓间充质干细胞成骨分化的生理活性。本文通过体外诱导成骨细胞系统,研究淫羊藿次苷Ⅰ和淫羊藿次苷Ⅱ对BMP/Runx2/OSX信号通路的影响,探讨其可能作用机制,同时比较两者的作用强度,为开发新药和发掘中医药潜在价值奠定基础。
1 材料与方法
1.1 试剂与药品
试剂:DMEM/F12培养基(Gibco,USA);甘油磷酸钠、磷酸化抗坏血酸(ASAP)、地塞米松、二甲基亚砜(DMSO)均购自Sigma公司;MEM-α细胞培养液(GIBCO 12571-063)、碱性磷酸酶(alkaline phosphates,ALP)测定试剂盒(AE91640Ra,批号:201512);骨钙素(BGP)检测试剂盒(AE90937Ra,批号: 201512);Ⅰ型胶原蛋白测定试剂盒(AE90739Ra,201512);BMP-2测定试剂盒(AE90181Ra,201512);NOGGIN重组蛋白(Peprotech公司,美国);β-甘油磷酸钠和茜素红(美国Sigma-Aldrich);类标准胎牛血清(兰州民海生物工程有限公司);RNAisoReagent、TaqDNA 聚合酶、反转录试剂盒、EasyDilution 稀释液均购自TaAaRa 公司,引物由TaAaRa 公司设计合成。
药品:淫羊藿次苷I(批号:121020)和淫羊藿次苷Ⅱ(批号:121117)均购自上海融禾医药科技有限公司(纯度大于98%)。
1.2 仪器
二氧化碳细胞培养箱(THERMO FORMA 3110)、酶标仪(BIO-TECK Synergy H1);蛋白核酸分析仪(DU640,美国贝克曼);低温高速离心机(MR1822,法国Jouan SA)。
1.3 方法及检测指标
1.3.1骨髓间充质干细胞培养及分组:大鼠BMSCs购于广州赛业有限公司,批号: 090405B01。采用胰蛋白酶消化法进行大鼠骨髓间充质干细胞的传代。
实验分组:①空白对照组:DMEM+10 mL/dL胎牛血清(含100 U/mL青、链霉素),即基础培养液;②阳性转化液组: DMEM +10 mL/dL胎牛血清(含 100 U/mL青、链霉素)+0.1 mol/L地塞米松+50μg/mL维生素C+10 mmol/L β-甘油磷酸钠,即成骨诱导液;③抑制剂组:成骨诱导液+1μg/mL Noggin;④淫羊藿次苷Ⅰ组:基础培养液+淫羊藿次苷Ⅰ1×10-5mol/L;⑤抑制剂+淫羊藿次苷Ⅰ组:基础培养液+淫羊藿次苷Ⅰ1×10-5mol/L+1μg/mL Noggin;⑥淫羊藿次苷Ⅱ组:基础培养液+淫羊藿次苷Ⅱ1×10-5mol/L;⑦抑制剂+淫羊藿次苷Ⅱ组:基础培养液+淫羊藿次苷Ⅱ1×10-5mol/L+1μg/mL Noggin。
1.3.2ALP 活性测定:采用ELISA法于成骨性诱导培养第6、9、12天检测ALP水平。具体方法按试剂盒说明进行,显色后于酶联仪450 nm处测定光密度D(λ)值,空白调零,绘制标准曲线,根据样品的光密度值在回归方程上计算对应的样品浓度以μg/L表示。
1.3.3矿化结节染色:将BMSCs 细胞接种后,使用成骨诱导培养基,放置于5.0% CO2,37 ℃的恒温培养箱中培养24 h后,进行上述各组药物干预,诱导14 d 以后将待测孔的培养基吸出,用茜素红染色方法染色,在倒置显微镜下观察。
1.3.4测定蛋白表达量:BGP、Ⅰ型胶原蛋白(COL-Ⅰ)、BMP-2蛋白浓度。采用ELISA法于成骨性诱导培养第12天,检测BGP、COL-Ⅰ、BMP-2含量,具体方法按试盒说明进行。用酶联仪在450 nm波长依序测量各孔的吸收度(OD值),绘制标准曲线,计算样品的实际浓度以μg/L表示(按说明书进行)。
1.3.5检测基因表达量:药物干预48 h,提取细胞RNA,实时定量PCR检测相关基因(ALP、BGP、Osterix、Runx2)表达量。RT-Real Time PCR 所用引物序列见表1。

表1 RT-Real Time PCR 所用引物序列Table 1 Primer sequences for real time RT-PCR
1.4 统计学处理

2 结果
2.1 对ALP活性的影响
ELISA法检测各组在第6、9、12天ALP的水平。与空白对照组比较,淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ组细胞内ALP水平均有升高(P<0.05),随着反应时间延长,细胞裂解液中ALP水平逐渐上升,第12天达到最高水平,达到阳性转化液组的80%以上。相应抑制剂组,BMSCs ALP活性明显降低。淫羊藿次苷Ⅰ组与淫羊藿次苷Ⅱ组比较,ALP活性在第9、12天,前者明显强于后者,且差异有统计学意义(P<0.05)。详见表2。
2.2 对细胞矿化结节的影响
矿化结节的出现是鉴定成骨细胞分化成熟的特征性标志,典型的矿化结节呈红褐色。可以看到,阳性转化液组(B)的矿化结节数目最多,连接成片,且体积较大;淫羊藿次苷Ⅰ组(C)和淫羊藿次苷Ⅱ组(D)的矿化结节均较空白对照组(A)多,且体积明显增大,其中淫羊藿次苷Ⅰ组的矿化结节多于淫羊藿次苷Ⅱ组。抑制剂组(E)的矿化结节数量明显少于阳性转化液组,且体积偏小,矿化水平降低;淫羊藿次苷Ⅰ+抑制剂组(F)及淫羊藿次苷Ⅱ+抑制剂组(G)组的矿化结节数量明显少于相应的淫羊藿次苷Ⅰ组及淫羊藿次苷Ⅱ组,且体积偏小,矿化水平降低。如图1所示。

表2 各组内第6、9、12 天的BMSCs ALP活性比较 Table 2 Comparison of ALP activity between each group on day 6, 9, and 12 after osteogenic induction
注:与空白对照组同期比较,★P<0.05;与相应各组第6天比较,*P<0.05;与阳性转化液组同期比较,△P<0.05;淫羊藿次苷Ⅰ组与淫羊藿次苷Ⅱ组同期比较,▲P<0.05;抑制剂组与相应药物组即⑤组与④组、⑦组与⑥组相比,□P<0.05。
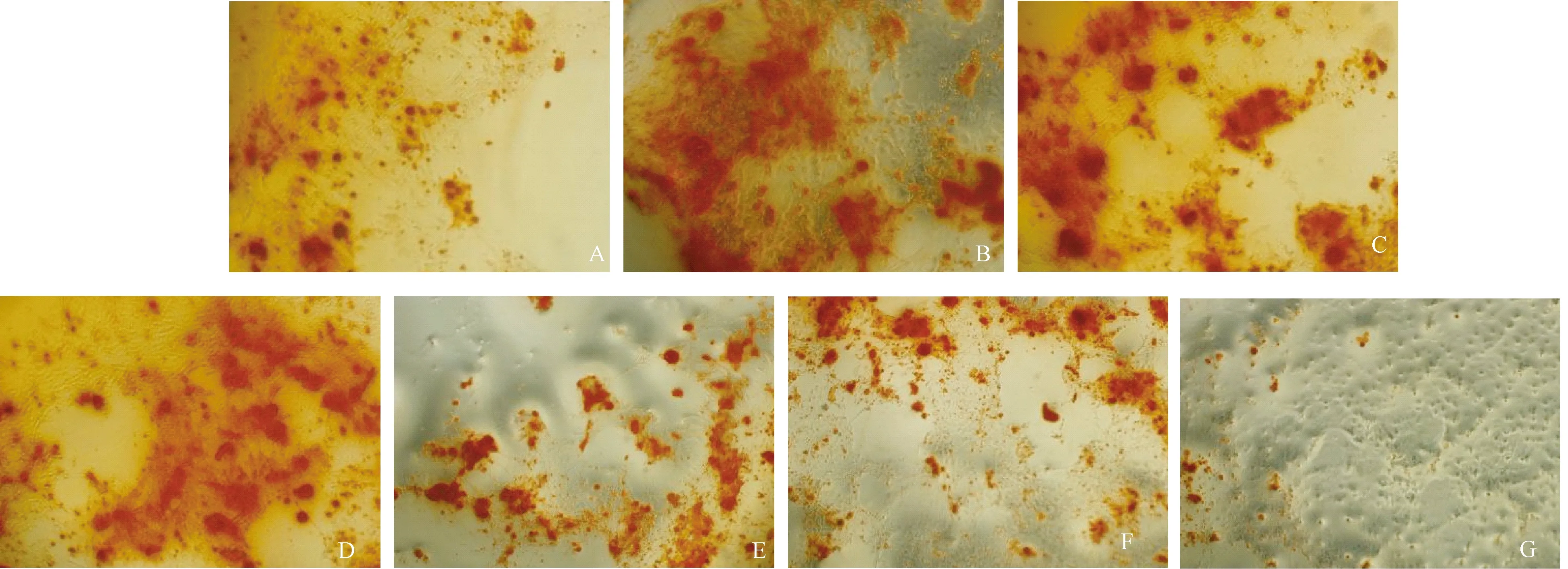
图1 各组对细胞矿化结节的影响(茜素红染色,×40) A:空白对照组; B:阳性转化液组;C:淫羊藿次苷Ⅰ组;D:淫羊藿次苷Ⅱ组;E:抑制剂组;F:淫羊藿次苷Ⅰ+抑制剂组;G:淫羊藿次苷Ⅱ+抑制剂组(放大倍数×40)Fig.1 Effects on mineralized nodule in BMSCs in each group (Alizarin red staining, ×40). A: Blank control group; B: Positive transformation solution group; C: Icariin I group; D: Icariin II group; E: Inhibitor group; F: Icariin I + inhibitor group; G: Icariin II + inhibitor group (×40)
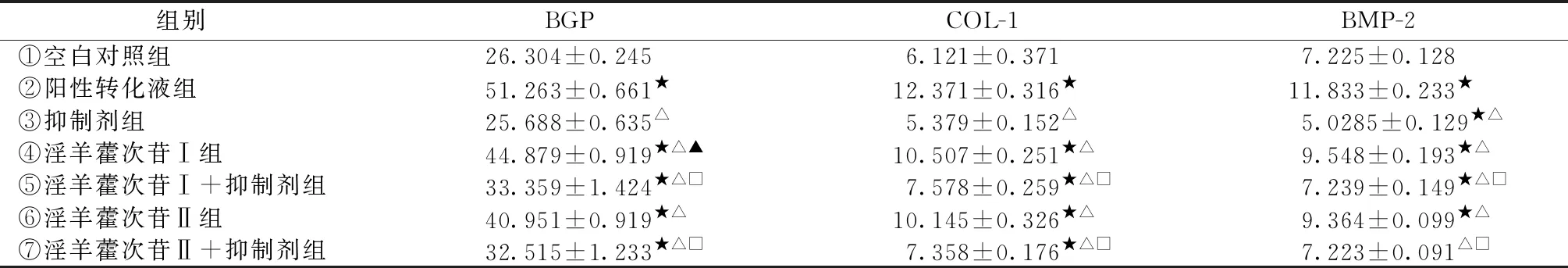
2.3 对骨髓间充质干细胞成骨分化过程中BGP、COL-Ⅰ、BMP-2蛋白表达的影响
BGP、COL-Ⅰ、BMP-2的蛋白表达量随着培养时间的延长而增加,在第12天达到了高峰值。与空白对照组比较,淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ组细胞BGP、COL-Ⅰ、BMP-2表达量均明显增加(P<0.05);与阳性对照液组比较,淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ组各因子蛋白表达量差异有统计学意义(P<0.05),但其活性已达到阳性转化液组的80%以上;相应抑制剂组细胞BGP、COL-Ⅰ、BMP-2表达量均显著降低,即⑤组与④组相比及⑦组与⑥组相比,差异有统计学意义(P<0.05);淫羊藿次苷Ⅰ与淫羊藿次苷Ⅱ组比较,淫羊藿次苷Ⅰ对促进BGP蛋白表达明显强于淫羊藿次苷Ⅱ,且差异有统计学意义(P<0.05),对 COL-Ⅰ、BMP-2的蛋白表达量,两者差异无统计学意义。详如表3。

组别BGP COL-1 BMP-2 ①空白对照组26.304±0.2456.121±0.3717.225±0.128②阳性转化液组51.263±0.661★12.371±0.316★11.833±0.233★③抑制剂组25.688±0.635△5.379±0.152△5.0285±0.129★△④淫羊藿次苷Ⅰ组44.879±0.919★△▲10.507±0.251★△9.548±0.193★△⑤淫羊藿次苷Ⅰ+抑制剂组33.359±1.424★△□7.578±0.259★△□7.239±0.149★△□⑥淫羊藿次苷Ⅱ组40.951±0.919★△10.145±0.326★△9.364±0.099★△⑦淫羊藿次苷Ⅱ+抑制剂组32.515±1.233★△□7.358±0.176★△□7.223±0.091△□
注:与空白对照组比较,★P<0.05;与阳性转化液组比较,△P<0.05;淫羊藿次苷Ⅰ组与淫羊藿次苷Ⅱ组比较,▲P<0.05;抑制剂组与相应药物组即⑤组与④组、⑦组与⑥组相比,□P<0.05。
2.4 对BMSCs骨向分化过程中BMP/RunX2/OSX信号通路相关因子ALP、BGP、Osterix、Runx2 mRNA表达的影响
淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ均可上调ALP、BGP,Runx2,并进一步调节其下游基因Osterix的转录表达。与空白对照组比较,淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ组ALP、BGP,Runx2及Osterix mRNA表达明显增强(P<0.05);与阳性对照液组比较,淫羊藿次苷Ⅰ对各因子mRNA表达差异有统计学意义(P<0.05),但其活性较强。淫羊藿次苷Ⅱ对ALP、Osterix的mRNA表达与阳性对照组无明显差异;相应抑制剂组相关转录因子表达量明显降低,即⑤组与④组、⑦组与⑥组相比,差异有统计学意义(P<0.05);淫羊藿次苷Ⅰ与淫羊藿次苷Ⅱ组比较,对Osterix因子mRNA的表达差异有统计学意义(P<0.05),后者明显强于前者,对其他各因子mRNA的表达二者无显著性差异。详见表4。
表4 各组细胞ALP、BGP、Osterix、Runx2 mRNA表达水平比较Table 4 Gene expression of ALP, BGP, Osterix, and RUNX2 in each group n=5)
注:与空白对照组比较,★P<0.05;与阳性转化液组比较,△P<0.05;淫羊藿次苷Ⅰ与淫羊藿次苷Ⅱ组比较,▲P<0.05;抑制剂组与相应药物组即⑤组与④、⑦组与⑥组相比,□P<0.05。
3 讨论
3.1 骨髓间充质干细胞成骨分化
骨髓间充质干细胞是骨髓中成骨细胞的祖细胞,在一定条件下可以成骨分化。BMSCs向成骨分化的重要表现如早期出现的标志物成骨细胞早期分化程度的碱性磷酸酶及较晚出现的作为成骨细胞向新骨生成过程特异性指标的骨钙素等。
3.2 BMP/Runx2/Osx信号通路与成骨分化
骨形态发生蛋白(bone morphogenetic proteins,BMPs)是转化生长因子(TGF-β)超家族的一员,可诱导骨髓间充质干细胞向成软骨细胞,成骨细胞和成牙本质细胞分化,促进成骨细胞功能。如BMP-2是促进骨形成最重要的细胞外信号分子之一[11]。成骨细胞的分化受多种信号通路调控,而其中BMP/Smads/Runx2/Osx信号通路与骨形成密切相关。BMP-2通过激活Smads信号传导和调节成骨基因转录如BMP-2、Runx2、Osterix等,而发挥其成骨作用。该信号通路同时受到抑制性smads和细胞外BMPs拮抗剂(如Noggin)等的调控[12]。
Runx2作为BMP-2的靶基因,直接调节成骨细胞特异性基因的表达,包括骨钙素、I型胶原和碱性磷酸酶等,在骨形成的多个阶段起调控作用。Osterix是Runx2的下游靶基因,受到Runx2的转录调控。Osterix是成骨细胞晚期分化所必需的特异性转录因子,调控多种重要的成骨基因的表达,如OC、骨粘素、骨桥素、骨钙素和Ⅰ型胶原蛋白等,对骨细胞的分化成熟和骨质形成有重要影响。Osterix与 Runx2调控成骨细胞不同的分化阶段。
本实验茜素红染色对细胞矿化结节的影响结果表明,淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ均具有促进骨髓间充质干细胞向成骨细胞增殖分化的趋势。ELISA实验结果显示,淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ均能促进ALP细胞活性,促进BGP、COL-Ⅰ、BMP-2的蛋白表达量,且随着干预时间延长而呈上升趋势,至第12天达到最高。与阳性对照液组比较,淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ组仍存在显著性差异,但其活性已达到阳性转化液组的80%以上。淫羊藿次苷Ⅰ与淫羊藿次苷Ⅱ组比较,淫羊藿次苷Ⅰ对ALP活性、BGP蛋白表达均明显强于淫羊藿次苷Ⅱ;对 COL-Ⅰ、BMP-2的蛋白表达量,两者无显著性差异。
RT-PCR结果显示,淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ均可显著上调成骨相关转录因子ALP、BGP、Runx2,并进一步调节其下游基因Osterix的转录表达,与空白对照组比较,具有显著性差异(P<0.05);与阳性对照液组比较,淫羊藿次苷Ⅰ对各因子蛋白表达量存在显著性差异(P<0.05),但其活性较强。淫羊藿次苷Ⅱ对ALP、Osterix的mRNA表达与阳性对照组无明显差异;淫羊藿次苷Ⅰ与淫羊藿次苷Ⅱ组比较,对Osterix因子mRNA的表达,二者具有显著性差异(P<0.05),后者明显强于前者;对ALP、BGP、Runx2因子mRNA的表达二者无显著性差异。
本研究表明,中药淫羊藿中的主要有效成分淫羊藿苷的代谢产物淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ,在体外能通过激活BMP/Runx2/Osx信号通路相关基因表达,提示可能通过促进 BMP-2蛋白,继而调控靶基因Runx2、Osterix的表达,进而促进BMSCs的成骨转化。BMP信号通路的阻滞剂(1 μg/mL Noggin重组蛋白)能明显抑制1×10-5mol/L淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ对BMSCs定向成骨分化的作用。本实验结果与翟远坤等[9]研究的淫羊藿次苷Ⅱ促进BMSCs成骨性分化的活性高于淫羊藿苷的实验结果具有一致性。
但是,其促进BMSCs定向成骨分化的信号转导通路可能含有多条且交错成网,其作用机制仍有待进一步深入研究。贺龙刚[13]等研究结果证明,淫羊藿次苷Ⅰ及其代谢产物淫羊藿次苷Ⅱ在体外均能通过AP-1/NFATc1信号通路抑制破骨细胞生成,且淫羊藿次苷Ⅱ作用优于淫羊藿次苷Ⅰ。翟远坤等[14]研究表明,淫羊藿次苷Ⅱ通过激活雌激素信号通路促进骨髓间充质干细胞的成骨性分化。但通过BMP/Runx2/Osx信号通路研究淫羊藿次苷Ⅰ及淫羊藿次苷Ⅱ对大鼠骨髓间充质干细胞成骨分化的实验研究本文尚属首次。
同时研究发现,淫羊藿次苷Ⅰ对ALP活性、BGP蛋白表达促进作用显著强于淫羊藿次苷Ⅱ;淫羊藿次苷Ⅱ对Osterix mRNA的表达要明显强于淫羊藿次苷Ⅰ。淫羊藿次苷Ⅰ虽然能够促进ALP活性、BGP蛋白表达,但对下游Osterix基因表达作用弱于淫羊藿次苷Ⅱ。该结果提示可能与两者化学结构上3位和7位取代基的不同有关。另外,淫羊藿次苷Ⅱ为淫羊藿次苷Ⅰ体内代谢产物,提示淫羊藿次苷Ⅰ可能通过生物转化成淫羊藿次苷Ⅱ,进一步增强对BMP/Runx2/Osx信号通路下游基因的调控,从而促进BMSCs定向成骨分化。这些可能是两者活性差异的主要原因,但具体机理还需做进一步的研究。
