不同温度下PEG-6000渗透胁迫对歪头菜种子发芽的影响
2019-06-04南志标
唐 伟,南志标
(兰州大学草地农业生态系统国家重点实验室 / 兰州大学农业农村部草牧业创新重点实验室 /兰州大学草地农业科技学院,甘肃 兰州 730020)
植物生命周期中种子发芽和早期幼苗生长阶段容易受环境因子的影响[1-2]。低温、高温和干旱胁迫可能抑制种子发芽或幼苗生长[3-4]。温度是调节种子发芽的重要环境因子[5],它影响着酶活性及物质代谢。温度过低或过高,会抑制酶的结构和代谢,从而影响酶的活性,最终抑制种子的正常代谢和发芽[6-7]。种子在适宜的温度范围内发芽对早期幼苗的生长和后期的植物群体建植有重要的作用[8-9]。多种豆科植物如紫花苜蓿(Medicago sativa)[2]、白三叶(Trifolium repens)[10]和箭筈豌豆(Vicia sativa)[4]种子发芽对温度响应的研究均发现,适宜温度可提高种子发芽率和发芽速率。
水分是影响种子发芽的另一个关键环境因子,他不仅影响发芽开始时间,而且还决定着发芽率,速率及其稳定性[5, 11]。以往的研究表明,水分胁迫会抑制豆科植物种子发芽,且水势的高低影响种子的吸水膨胀和发芽率。种子只有吸水到一定的临界点才能发芽,这个临界点的水势为种子发芽的基础水势,即种子发芽所需的最低水势[4, 12-13]。研究发现,当外界条件高于基础水势时,种子发芽速率与外界水势呈正相关关系,且种子群体的最终发芽率随水势的降低而降低。尽管干旱胁迫抑制种子吸胀从而限制发芽,但适宜的温度可以缓解干旱对种子发芽的不利影响[8, 14]。种子发芽对温度-水分互作具有适应性,这决定了田间种子发芽时间及幼苗生长能力[5]。因此,研究种子发芽所需的温度和水分条件有助于分析田间条件下植物种子发芽、幼苗生长及建植难易程度的原因[1-2]。
歪头菜(Vicia unijuga)是豆科野豌豆属的多年生草本植物,主要分布在中国东北、华北、西北、华东、华中以及西南等地,分布区海拔在440~4 400 m[15]。歪头菜是青藏高原重要豆科牧草,具有营养丰富、再生性能强、适口性好、产量高等特点,成为该地区家畜饲料中蛋白质的重要来源[16]。但由于青藏高原海拔高,气候严酷,春季低温和干旱胁迫等自然客观因素限制了歪头菜的种子发芽和幼苗生长[17]。前人对歪头菜的研究主要集中在独立的温度或渗透胁迫对种子发芽特性的影响[4, 18-19],然而,在不同温度条件下,不同浓度的PEG-6000渗透胁迫对歪头菜种子发芽和幼苗生长的影响较为系统的研究未见报道。基于此,本研究以歪头菜种子为试验材料,在不同温度条件下,分析聚乙二醇(PEG-6000)模拟渗透胁迫对其发芽和幼苗生长的影响,探究其种子发芽和幼苗生长所需的适宜条件,以期为这一优良豆科牧草在青藏高原播种应用及建植提供理论依据。
1 材料和方法
1.1 供试材料
供试歪头菜种子于2015年9月采自甘肃省甘南藏族自治州夏河县兰州大学草地农业科技学院野生歪头菜驯化选育基地 (海拔 3 000 m,102°30′ E,35°16′ N)。根据夏河县气象站近 30年的记录,该地区年均降水量451 mm,其中4月到9月的降水量占全年80%以上。年均气温3.5 ℃,年均蒸发量 1 343 mm,年均日照时数 2 425 h,年均无霜期81 d。采回的种子风干后,贮藏于4 ℃种子低温贮藏室备用。
1.2 试验设计
歪头菜种子具有物理休眠。在开展种子发芽试验前,用硫酸浸种法打破其物理休眠[18];歪头菜种子用硫酸浸种15~20 min,再用自来水冲洗15 min。破除休眠的种子在5%次氯酸钠中浸种消毒5 min,用去离子水清洗,备用。
本研究设置3个温度(15、20和25 ℃)和6个不同浓度的PEG-6000溶液(0、-0.2、-0.4、-0.6、-0.8和-1.0 MPa),0 MPa以无菌蒸馏水作对照,每处理4次重复。根据Michel和Kaufmann[20]的公式配置不同浓度的PEG-6000溶液:

式中:ψS为溶液的水势(bar);C为PEG-6000的浓度 (g·kg-1),T为温度常数 (℃)。
种子发芽试验采用TP法。随机选取50粒种子置于铺有两层滤纸的培养皿中,每皿分别加入5 mL蒸馏水或相应浓度的PEG溶液。为保持PEG溶液浓度稳定,将培养皿用封口膜进行密封避免水分蒸发,每两天更换一次。将各处理在相应恒温培养箱中培养,光照强度为 60 μmol· (m2·s)-1(12 h 光/12 h暗),湿度50%。每天记录正常发芽的种子数(以胚芽突破种皮2 mm为标准)。当连续培养15 d不再有种子发芽时,试验结束。
1.3 计算方法
发芽率(GP)=(在发芽结束期正常发芽种子数/供试种子数)×100%;
发芽速率(GR)=1/(20%种子发芽时间);
发芽指数(GI)=∑(Gt/Dt)。
式中:Gt指t时间内的发芽数,Dt指相应的发芽日数[21-24]。
活力指数(VI)=GI×(SL+RL)。
式中:GI指发芽指数,SL和RL分别为发芽15 d后幼苗的胚芽长(mm)和胚根长(mm)[21-24]。
种子发芽基础水势的估计:通过重复概率单位回归分析(repeated probit)计算得到发芽的基础水势,水势时间值代入间隔为0.1 MPa·d,直到所获回归方程残差均方最小[25]。然后根据下面公式计算水势时间值:

式中:Ψ为发芽水势,θH为水势时间,Ψb(50)为基础水势均值,δΨb为发芽基础水势标准偏差。
1.4 数据分析
使用Excel 2013软件进行数据统计,利用SPSS 22.0(SPSS公司,美国)数据分析软件对所测数据进行分析,制图用Origin 9.0 (OriginLab公司,美国)完成。采用双因素方差分析探究温度和水势及其互作对歪头菜种子发芽和幼苗生长性状的影响。平均值多重比较采用LSD法在0.05水平下分析显著性,Duncan法对上述发芽指标进行多重差异性比较;采用回归分析种子发芽的概率单位与发芽的水势值的关系。
2 结果与分析
2.1 不同温度和渗透势对歪头菜种子发芽的影响
双因素方差分析结果显示,温度、渗透势及其互作对歪头菜种子发芽率、发芽速率、发芽指数和活力指数有显著 (P< 0.001)影响 (表1)。
2.1.1 种子发芽率
歪头菜种子发芽率(GP)在不同温度(15~25 ℃)和水势(-1.0~0 MPa)条件下的变化如图1所示。平均水势,GP在20 ℃条件下最高,为61.3%,显著(P<0.01)高于 15 ℃(57.5%)和 25 ℃(49.8%),15 ℃ 下 GP显著 (P< 0.01)高于 25 ℃。随着渗透胁迫的增加,GP显著 (P< 0.05)降低。较低水势 (-1.0~-0.6 MPa)对GP的抑制显著(P< 0.05)高于较高水势(-0.4~-0.2 MPa)。渗透胁迫抑制种子吸胀从而限制发芽,但适当的增温可以缓解渗透胁迫对GP的不利影响,即使在最低水势-1.0 MPa条件下,20 ℃下GP依然明显高于15 ℃,提高了31.5%。但在较低水势(-1.0~-0.6 MPa)时,25 ℃ 下 GP明显低于 15 ℃。
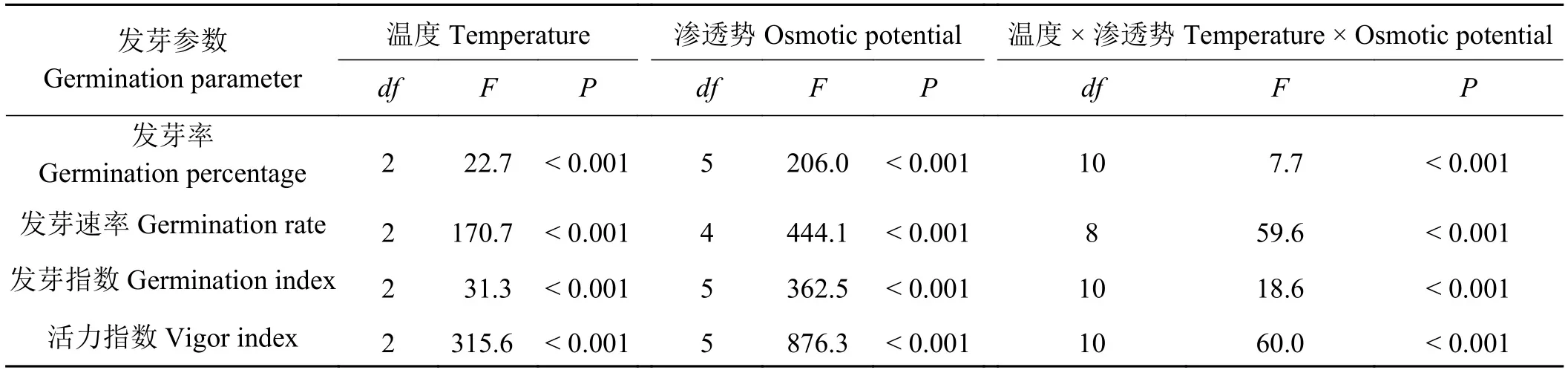
表1 温度和渗透势对歪头菜种子发芽影响的双因素方差分析Table 1 Two way ANOVA analysis of the effects of temperature, osmotic potential and their interaction on the seed germination traits of Vicia unijuga
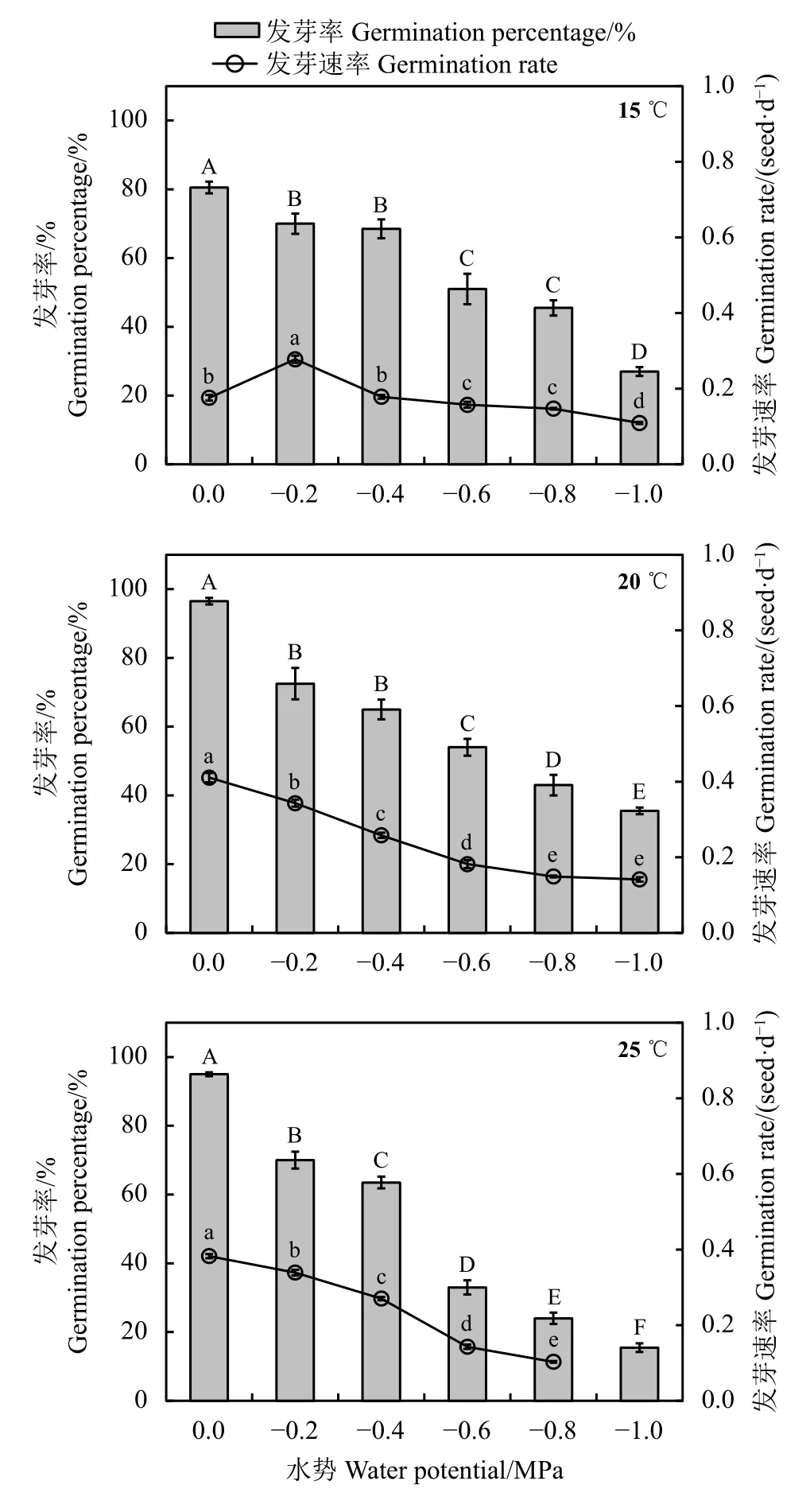
图1 歪头菜种子在不同温度和不同水势条件下的发芽率(GP)和发芽速率(GR)Figure 1 Germination percentage (GP) and germination rate(GR) of scarified V. unijuga seeds at different treatments
2.1.2 发芽速率
歪头菜种子发芽速率(GR)在不同温度(15~25 ℃)和水势 (-1.0~0 MPa)条件下的变化如图1所示。在不同温度处理下,GR存在显著差异(P< 0.05)。平均水势在 20 ℃ 条件下 GR 最高,达 0.269 seed·d-1,其次是 25 ℃ 条件下的 GR(0.248 seed·d-1);并且 15 ℃条件下 GR(0.187 seed·d-1)显著 (P< 0.05)低于 25 ℃。在对照0 MPa水势下,GR在20 ℃和25 ℃下最高,分别为 0.411和 0.383 seed·d-1。而在 15 ℃ 条件下,GR 在-0.2 MPa 水势下最高,为 0.278 seed·d-1。此外,适宜的增加温度可以缓解由渗透胁迫降低对GR的负面影响,例如,与15 ℃条件相比,20 ℃和 25 ℃ 均显著 (P< 0.05)提高歪头菜种子在-0.4~0 MPa 水势下 GR;在-1.0 MPa 水势下,20 ℃ 条件下 GR 明显 (P< 0.05)高于 15 ℃。在-1.0~-0.8 MPa水势下,15 ℃ 条件下 GR 明显 (P< 0.05)高于 25 ℃。
2.1.3 发芽指数
歪头菜种子发芽指数(GI)在不同温度(15~25 ℃)和水势(0~-1.0 MPa)条件下的变化如图2所示。平均水势,20 ℃ 条件下种子 GI显著 (P< 0.05)高于15 ℃ 和 25 ℃,15 ℃ 下 GI与 25 ℃ 无显著差异 (P>0.05)。随着渗透势的增加,歪头菜种子GI逐渐(P<0.05)降低。具体而言,在对照0 MPa条件下,歪头菜种子分别在15、20和25 ℃下的GI最高。较低水势(-1.0~-0.6 MPa)对GI的抑制作用显著高于较高水势(-0.4~-0.2 MPa)。适当的增温可以缓解渗透胁迫对GI的抑制作用,即使在最低水势-1.0 MPa条件下,20 ℃ 条件下 GI显著 (P< 0.05)高于15 ℃。但在较低水势(-1.0~-0.6 MPa)条件下,25 ℃ 下 GI显著 (P< 0.05)低于 15 ℃。
2.1.4 活力指数
歪头菜种子活力指数(VI)在不同温度(15~25 ℃)和水势(-1.0~0 MPa)条件下的变化如图3所示。平均水势,VI在 20 ℃ 条件下显著 (P< 0.001)高于 15 ℃和 25 ℃,同时 25 ℃ 显著 (P< 0.001)高于 15 ℃。随着渗透胁迫的增加,VI显著(P< 0.05)降低。较低水势(-1.0~-0.6 MPa)对VI的抑制高于较高水势(-0.4~-0.2 MPa)。适当的增温可以缓解渗透胁迫对VI的不利影响,在较低水势(-1.0~-0.6 MPa)条件下,20 ℃ 条件下 VI显著 (P< 0.05)高于 15 ℃。但在较低水势 (-1.0~-0.8 MPa)条件下,25 ℃ 下VI显著 (P< 0.05)低于 15 ℃。
2.2 基于Probit分析对发芽基础水势的估计

图2 不同温度和渗透势对歪头菜种子发芽指数的影响Figure 2 Effect of different temperatures and osmotic potentials on Vicia unijuga seeds germination index

图3 不同温度和水势对歪头菜种子活力指数的影响Figure 3 Effect of different temperatures and water potentials on Vicia unijuga seeds vigor index
由重复Probit回归分析获得的歪头菜种子的Ψb因温度而异,Ψb的变化随温度的升高呈先升后降的趋势;20 ℃条件下歪头菜种子抗渗透胁迫能力最强,Ψb为 -1.27 MPa,25 ℃ 条件下歪头菜种子抗渗透胁迫能力最弱,Ψb为-1.05 MPa。15 ℃~25 ℃条件下,种子发芽的平均水势时间为4.61 MPa·d(表2)。
2.3 不同温度下PEG渗透胁迫对歪头菜幼苗芽和根生长的影响
温度、水势及其互作对歪头菜幼苗胚芽长(SL)和胚根长 (RL)的影响均达显著水平(P< 0.001)(表3);平均水势,20 ℃条件下SL最高为24.4 mm,显著(P< 0.001)高于15 ℃(14.1 mm)和25 ℃(19.3 mm),15 ℃ 下 SL 显著 (P< 0.01)低于 25 ℃(表4)。随着渗透胁迫的增加,SL显著(P< 0.05)降低。较低水势(-1.0~-0.6 MPa)对 SL的抑制显著 (P< 0.05)高于较高水势(-0.4~-0.2 MPa)。渗透胁迫抑制胚芽生长,适当的增温可以缓解渗透胁迫对胚芽生长的不利影响,即使在最低水势-1.0 MPa条件下,歪头菜种子在20 ℃条件下的SL,可达10.4 mm,高于15和25 ℃,并且15 ℃下SL高于25 ℃时的。
平均水势,20 ℃条件下RL最高为11.3 mm,显著 (P< 0.001)高于 15 ℃(8.7 mm)和 25 ℃(8.1 mm)(表4)。随着渗透胁迫的增加,RL变化呈下降趋势。25 ℃条件下,在对照0 MPa水势时RL最高;15和20 ℃条件下,在-0.2 MPa水势时RL最高。较低水势(-1.0~-0.6 MPa)对RL的抑制显著(P<0.05)高于较高水势 (-0.4~-0.2 MPa)。渗透胁迫抑制胚根生长,适当的增温可以缓解渗透胁迫对胚根生长的不利影响,即使在最低水势(-1.0 MPa)条件下,歪头菜种子在20 ℃条件下RL,可达5.3 mm,高于 15 ℃ 和 25 ℃。
3 讨论
地域性气候与植物种子的发芽特性是息息相关的,这种特性是植物对此环境长期适应而形成独特的发芽策略[5]。种子发芽是植物生活史的重要阶段,容易受环境因子(温度和水分)的影响[5],探究种子发芽应对低温和高温或干旱胁迫的响应策略对植物种群建植具有重要的意义[26]。本研究发现,歪头菜种子发芽率,发芽速率、发芽指数和活力指数以及胚芽长和胚根长在20 ℃时最大,显著高于15或25 ℃。这与聂斌[18]对甘南歪头菜种子发芽率和发芽指数的研究结果类似,较高温度(20~25 ℃)下发芽率和发芽指数显著高于较低温度(5~15 ℃);Hu 等[4]研究表明,20 ℃ 条件下,歪头菜的发芽速率最大。周娟娟等[27]对高山鹅观草(Roegneria tschimganica)和纤毛鹅观草(Roegneria ciliaris)、宋采博和王波[28]对白三叶、鱼小军等[29]对醉马草(Achnatherum inebrians)也报道了类似的研究结果。可能的原因是种子发芽过程中,一定范围内的增温提高种子内部的酶活性和物质代谢水平,进而促进种子发芽;亚适温条件下水解酶活性较低导致种子中的养分不易水解,引起发芽缓慢;超适温条件引起呼吸速率及淀粉酶活性高峰提前,种子内的营养物质衰竭,导致种子发芽率与发芽速率下降[30-34]。青藏高原海拔高,早春阶段的低温抑制歪头菜种子发芽,这与在高山草原上低温抑制长芒草(Stipa bungeana)、山野豌豆(Vicia amoena)和窄叶野豌豆(Vicia angustifolia)种子发芽和早期幼苗建植的结果类似[4, 35]。尽管如此,低温让这些未能发芽的歪头菜种子保持休眠状态,在土壤中形成土壤种子库;这种延迟发芽的策略对于其在青藏高原环境下的适应性和后期的成功建植具有重要意义。此外,歪头菜种子发芽积温时间较高[36],这一特性能够避免歪头菜种子在短暂高温条件下迅速发芽,从而降低了由于青藏高原气候剧烈变化造成的播种后发芽的种子和出土幼苗迅速死亡带来的建植失败风险。

表2 基于重复Probit回归估计的基础水势Table 2 Estimation of water potential value based on repeated probit analysis for Vicia unijuga with three different temperature

表3 温度和渗透势对歪头菜幼苗生长影响的双因素方差分析Table 3 Two way ANOVA analysis of the effects of temperature and osmotic potential on the seedling growth parameters of Vicia unijuga
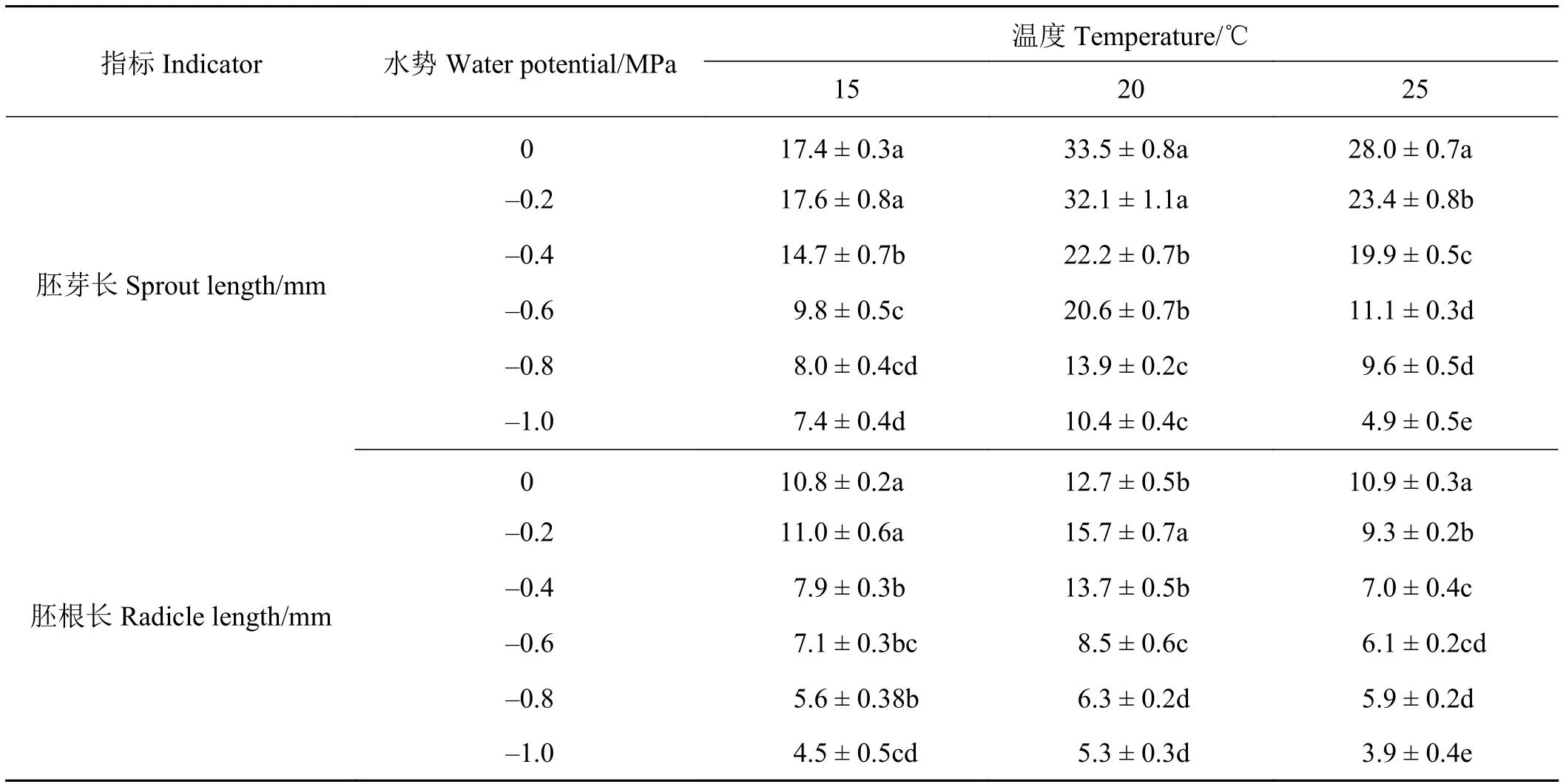
表4 不同温度和水势对歪头菜胚芽长和胚根长的影响Table 4 Effect of different temperatures and water potentials on Vicia unijuga sprout length and radicle length
种子发芽期间,干旱胁迫抑制植物种子发芽和生长,进而限制作物播种后实现快速建植,并造成种子的发芽损耗[4, 37-40]。在青藏高原,晚春时期的温度逐渐升高有利于歪头菜种子发芽,但是此时的降水量依然很低,导致歪头菜种子不能快速地发芽。本研究表明,随着渗透胁迫的增加,歪头菜种子的发芽率、发芽速率、发芽指数、活力指数、胚芽长和胚根长的变化随渗透胁迫的加剧表现出持续下降趋势;尤其是高浓度的PEG溶液(-1.0 MPa)抑制歪头菜种子的发芽,平均发芽率低于30%,并且出现发芽进程减缓,出苗不整齐,生长缓慢和幼苗弱小等现象。这与Hu等[4]的研究报道一致,他们发现极低水势严重抑制野豌豆属(Viciaspp.)植物种子发芽。可能的原因是干旱胁迫抑制种子体内酶的代谢和酶活性,进而抑制正常的生理代谢,降低种子活力[41]。本研究发现,适当的增温可以缓解干旱胁迫对种子吸胀、发芽和生长的抑制作用。较低的水势(-1.0~-0.6 MPa)下,20 ℃条件下种子发芽和幼苗生长显著高于15或25 ℃;这些结果的差异反映了温度和模拟干旱胁迫对歪头菜种子发芽的交互影响,说明歪头菜种子具有较强的适应能力,是其长期生活在高寒干旱的环境中形成了发芽策略,有利于其在气候多变的青藏高原环境下构建种群。本结果与适宜的温度可缓解干旱胁迫对豇豆(Vigna unguiculata)[42]和柠条锦鸡儿(Caragana korshinskii)[26]种子发芽不利影响的报道一致。在适宜的温度条件下,干旱胁迫对种子发芽的抑制强度低于亚适温和超适温条件,通过激活体内酶保护机制,累积渗透调节物质用以抵御水分胁迫,维持正常的生理代谢,进而保证种子的正常发芽和幼苗生长[14]。
在一定的温度范围内,种子的吸水能力、胚根和胚芽的生长,可以决定其发芽的最低水势[43]。在不同温度下歪头菜种子的基础水势值存在显著差异,在25 ℃最高,而在20 ℃最低。这进一步说明歪头菜在青藏高原长期的进化或选育过程中,20 ℃下种子较低的基础水势值有利于抵抗青藏高原不规律降水带来的干旱胁迫,提高种子发芽率。类似结果在青藏高原生长的窄叶野豌豆[36]和箭筈豌豆[19]等豆科植物的研究中均有发现。此外,歪头菜种子发芽的水势时间值较大,播种后由于春季短期阵性降水使种子短暂吸水并进入水合状态,但并未发芽,保持次生休眠以抵抗干旱胁迫,贮存于土壤中,有效降低了“闪苗”的风险,进而降低种子的发芽损耗,维持稳定的种子发芽和幼苗生长,最终建植成功[44-45]。因此,在降水多变的青藏高原,歪头菜对此环境长期适应而形成独特的发芽策略,即稳定的发芽率,提前或是推迟发芽时间有利于增加其种子的存活率和适应能力[46]。
综上所述,20 ℃是歪头菜种子发芽和生长最适宜的温度;在渗透胁迫下,20 ℃可以对其种子发芽和幼苗生长起着积极的作用,温度-水分互作对歪头菜种子发芽和幼苗生长具有重要的生态意义,本研究的结果可以为歪头菜种质资源在青藏高原的合理利用提供有效的策略。根据青藏高原年均温和降水的变化规律以及歪头菜种子的发芽特性,春季播种容易受到低温和干旱对种子发芽和幼苗生长的不利影响;因此,歪头菜更适合早夏播种,此时升高的气温和充足的降雨有利于歪头菜种子发芽,幼苗生长和建植,进而提高农业生产。
