外源水杨酸对草地早熟禾抗褐斑病的诱导与抗病基因PR1和NPR1的表达的影响
2019-06-04尉春雪苏浩天张晓宇何文菡郑大柽尹淑霞
尉春雪,苏浩天,张晓宇,何文菡,郑大柽,尹淑霞
(北京林业大学林学院,北京 100083)
草坪作为城市绿地的重要组成部分,在城市生态系统中发挥着重要作用。与其他植物相比,草坪由于定期修剪,使得病原菌更易侵染,因此草坪病害发生更严重、更普遍。提高草坪抗病性也是草坪科学研究的重点和难点。
褐斑病是草坪的一种重要土传真菌病害,也是所有草坪病害中分布最广的病害之一[1]。该病几乎能侵染所有已知的草坪草品种,具有病原物复杂、发病率高、传染迅速、危害严重和反复发病等特点,在整个北京地区冷季型草坪上普遍发生,发病率高达80%以上[2]。
水杨酸(salicylic acid,SA)是非生物源植物抗病激活剂,能诱导多种植物对多种真菌、细菌及病毒病害产生抗性,且具有高效、广谱、对环境无害、使用简单、成本低廉等优点,在植物抗病反应中起着非常重要的作用[3]。有研究表明,SA可诱导苜蓿(Medicago sativa)对霜霉病产生抗性[4],诱导水稻(Oryza sativa)幼苗抗稻瘟病[5]和白叶枯病[6],诱导高羊茅(Festuca arundinacea)抗弯孢霉叶斑病[7],诱导黄瓜(Cucumis sativus)抗霜霉病[8]、诱导玉米(Zea mays)抗大斑病[9],诱导月季(Rosa chinensis)抗黑斑病[10],诱导匍匐翦股颖(Agrostis stolonifera)抗镰刀菌枯萎病[11],诱导车前(Plantago asiatica)抗菌核病[12],诱导苹果(Malus pumila)抗斑点落叶病[13]等。在草坪草中,外源喷施水杨酸的研究多集中于其对草坪草耐盐、抗旱、耐热等逆境胁迫的效果,对其诱导草坪草抗病性的研究还很少。
草地早熟禾(Poa pratensis)是我国北方地区应用最广泛的草坪草种之一,具有耐寒、青绿期长、坪观质量高等优点。因立枯丝核菌(Rhizoctonia solani)侵染引起的褐斑病是草地早熟禾的重要病害之一,该病常造成草坪坪观质量下降甚至草坪枯死[14]。研究表明,植物在与病原物共同进化过程中,主要形成了被动和主动两类抗病类型,其中主动抗病型主要是通过分子抗病物质来实现。Guo等[15]研究表明C3H型锌指蛋白在烟草(Nicotiana tobacum)对立枯丝核菌抗性中起正调控作用。殷萍萍[16]研究得出结缕草(Zoysia japonica)CPI基因能抑制立枯丝核菌体内蛋白酶,从而抑制其生长;富亮氨酸重复受体蛋白激酶(LRR-RLKs)基因编码的LRR蛋白可直接或间接识别病菌效应子,激活相应的防卫反应。
本研究通过对草地早熟禾施用外源SA,并接种立枯丝核菌,观察其诱导草地早熟禾抗褐斑病的效果,检测相关抗病基因表达情况,探讨外源SA对草地早熟禾褐斑病诱抗效果及分子调控机理,为防治草地早熟禾褐斑病及草坪草抗病育种提供参考。
1 材料和方法
1.1 供试材料
在北京林业大学草坪试验站选取健康、成熟的草地早熟禾品种午夜 (Poa pratensis‘Midnight’)的草皮(草坪草高度4 cm,所用土壤为壤土且养分充足,建植1 a),2017年6月10日移栽到40盆直径约15 cm的花盆中,室外培养,每天进行浇水,9月中旬施过一次复合肥。
水杨酸SA由天津市光复精细化研究所生产,立枯丝核菌由北京林业大学草坪研究所提供,植物总RNA提取试剂盒购于E.Z.N.A公司,反转录试剂盒购于北京全式金生物技术有限公司,荧光定量用UltraSYBR Mixture试剂盒购于CWBIO公司。
1.2 试验方法
1.2.1 喷施SA与接种处理
先通过预试验确定外源SA对午夜褐斑病的最佳诱抗浓度为0.05 mmol·L-1。而后挑选健康、生长一致的早熟禾12盆。设3个处理:A处理为每盆喷施 0.05 mmol·L-1SA 30 mL 并在 3 d 后接种立枯丝核菌(SA+接菌);B处理是每盆喷等量清水并在3 d后与A组一起接种立枯丝核菌(仅接菌);C处理是与A组同时外施SA,但不进行接菌处理(仅SA处理)。每处理4次重复,而后置于保鲜袋中封口进行保湿(相对湿度70%)培养24 h。
1.2.2 立枯丝核菌接种方法
将储存于本实验室的立枯丝核菌菌丝体与高温灭菌的高羊茅种子在密闭的培养瓶中共培养3周,待立枯丝核菌侵染了高羊茅种子后,将种子以30 g·m-2的量均匀撒在盆栽的草地早熟禾草坪上进行接种。接种在SA喷施3 d后进行,A、B处理接种等量的感菌种子,C处理不接种。而后将草坪草置于人工气候箱中进行培养,昼夜温度为35 ℃/25 ℃,光暗时间 16 h∶8 h,相对湿度 70%。
1.3 指标测定
1.3.1 诱导抗病效果观察
接种后每天观察草坪草发病情况,统计发病植株数量,每盆随机挑选20个叶片,按照病叶等级分级标准[17],根据感病面积分为5级,未感病记为0,感病面积1%~20%为1级,21%~40%为2级,41%~60%为3级,61%~80%为4级,81%~100%为5级。按照如下公式计算发病率、病情指数和诱抗效果。
发病率 = (发病植株数 / 总株数) × 100%;
病情指数 = 100 × ∑(各级病叶数 × 各级代表值) /(调查总叶数 × 最高级代表值);
诱抗效果 = (B处理病情指数-A处理病情指数) /B 处理病情指数 × 100%。[18]
1.3.2 抗病基因表达量检测
在SA喷施前(T0)和喷施后第1天(T1)、第3天即病原菌接种前(T2)、病菌接种后第3天(T3)、第4天(T4)、第5天(T5),分别取样并提取叶片组织总RNA,检测抗病相关基因PR1、NPR1表达量的变化。具体为提取各样品总RNA后进行反转录,方法参照反转录说明书(北京全式金生物技术有限公司,Code:AT-311),获得cDNA第一链后将其用ddH2O稀释至 80~100 ng·μL-1置于-20 ℃ 冰箱保存,供下一步荧光定量反应使用。荧光定量反应中内参基因选用草地早熟禾18S基因,待测抗病基因选用草地早 熟禾PR1和NPR1基因,通过 Primer Premier 5.0软件设计特异PCR引物,得到18S、PR1、NPR1上下游引物,3组引物序列如表1所列。
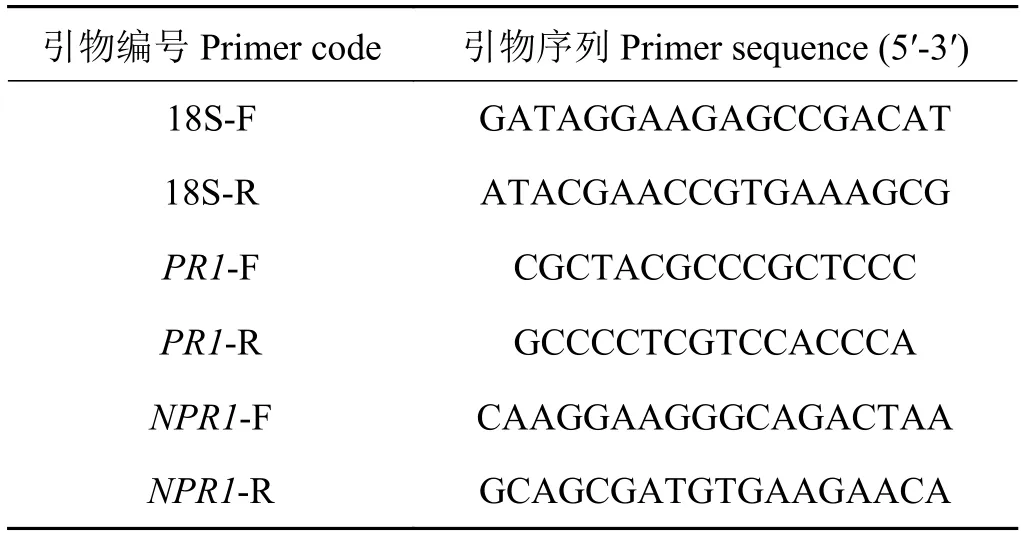
表1 引物序列Table 1 Primer sequence
荧光定量反应方法参照荧光定量说明书(CWBIO,Code: CW0957M)。反应体系为 2x Ultra-SYBR Mix 10 μL,Forward Primer与 Reverse Primer各 1 μL,cDNA 1 μL,ddH2O 7 μL,共 20 μL。反应程序为 95 ℃ 预变性 10 min;95 ℃ 变性 10 s,55 ℃退火 30 s,72 ℃ 延伸 32 s,重复 40 个循环。熔解曲线分析与数据分析使用CFX Manager软件,使用2-ΔΔCt方法进行数据分析。
1.3.3 数据处理
用SPSS 16单因素方差分析法对3组的发病率、病情指数、PR1相对表达量、NPR1相对表达量进行方差分析,LSD方法进行比较,Excel 2007进行作表和制图。
2 结果与分析
2.1 发病率
接种立枯丝核菌后的第3天,A处理和B处理草坪草开始感病(图1),而C处理草坪草因未接种,没有出现感病现象。从接种后第3天草坪草发病,随着时间的推移,A处理与B处理的发病率均呈现上升趋势,且添加了SA的A处理组显著低于B处理组(P< 0.05)。至接种后第6天,B处理发病率达47%,A处理发病率达30.4%,之后A、B处理发病率均明显上升,草坪质量开始急剧下降,至接种后第9天,A处理发病率达到66.2%,B处理发病率达到81.3%。由此可以说明,外施SA能显著减轻草地早熟禾午夜的发病率,但其作用时间有限,接种后第7天A处理发病率达到52.5%,草坪质量已经不可接受。
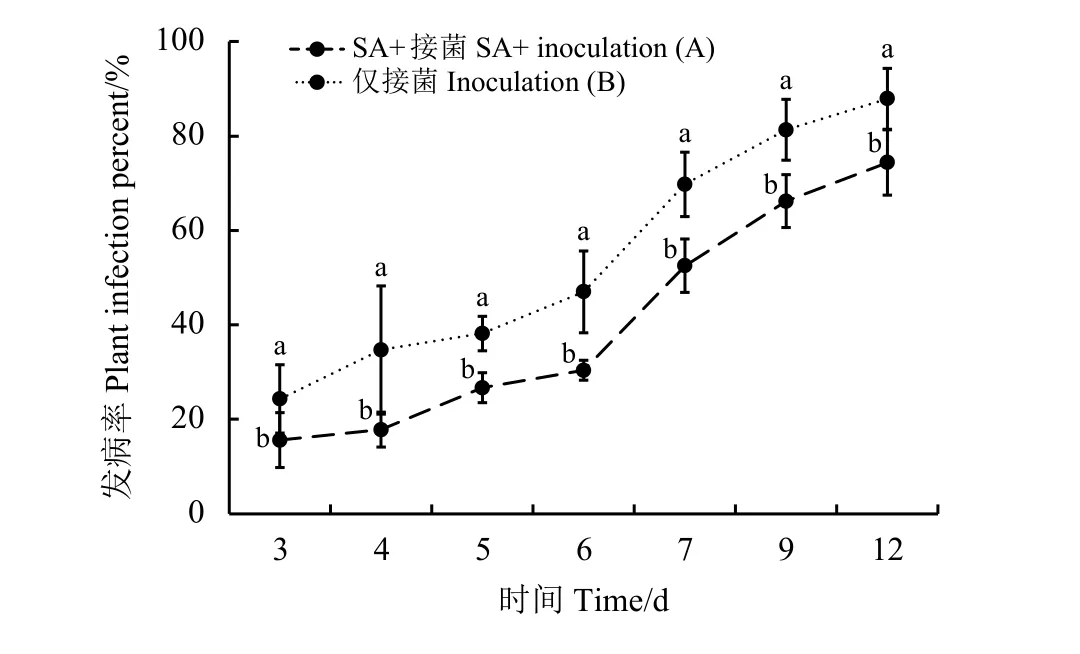
图1 接种后午夜草地早熟禾发病率Figure 1 Plant infection percent of Kentucky bluegrass‘Midnight’ post inoculation
2.2 病情指数
A处理与B处理草坪草的病情指数都随着时间的推移而升高(图2),但在试验期间,A处理的病情指数始终显著低于B处理(P< 0.05)。至接种后第12天,A处理的病情指数为20.8,显著低于B处理的34.5(P< 0.05)。由此说明,外施SA处理不仅可以减轻草地早熟禾褐斑病的发生,还可以显著降低其病情的严重程度。

图2 病原菌接种后草地早熟禾的病情指数Figure 2 The disease index of Kentucky bluegrass ‘Midnight’post inoculation
2.3 诱抗效果
SA对草地早熟禾褐斑病的诱抗效果在接种后第7天,达53%(图3),诱抗效果先降低后升高又降低,除第6天外,其余总体差异不显著(P> 0.05),由此可见,在发病期间,SA对草地早熟禾褐斑病的诱抗效果较明显。
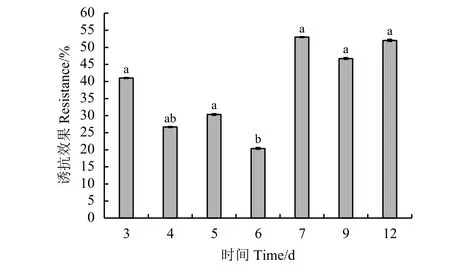
图3 病菌接种后SA对草地早熟禾午夜褐斑病的诱抗效果Figure 3 Efficacy of resistance induced by SA to brown patch in Kentucky bluegrass ‘Midnight’ post pathogen inoculation
2.4 SA诱导后抗病基因表达量变化
A、C处理喷施SA后的第1天(T1),草地早熟禾PR1基因的相对表达量是喷施SA前(T0)的2.7倍(图4),之后,A处理先降低后升高再降低,C处理逐渐降低且与喷施SA前(T0)差异不显著(P> 0.05)。A、B处理,在接菌后第3天(T3)PR1基因相对表达量均达到最高,分别为T0时的3.5倍(P< 0.05)和 3.2倍 (P< 0.05),与 T2即接种前相比差异显著(P< 0.05),可见立枯丝核菌的侵染促进了草地早熟禾PR1基因的表达,这也是植物应对病原菌侵染的一种防御反应。随后二者的PR1基因相对表达量虽有所下降,但在T5即接种后第5天时仍高于T0,可见立枯丝核菌的侵染使得草地早熟禾PR1基因持续高表达,从而引起植物体内与之相关的基因表达发生一系列变化。
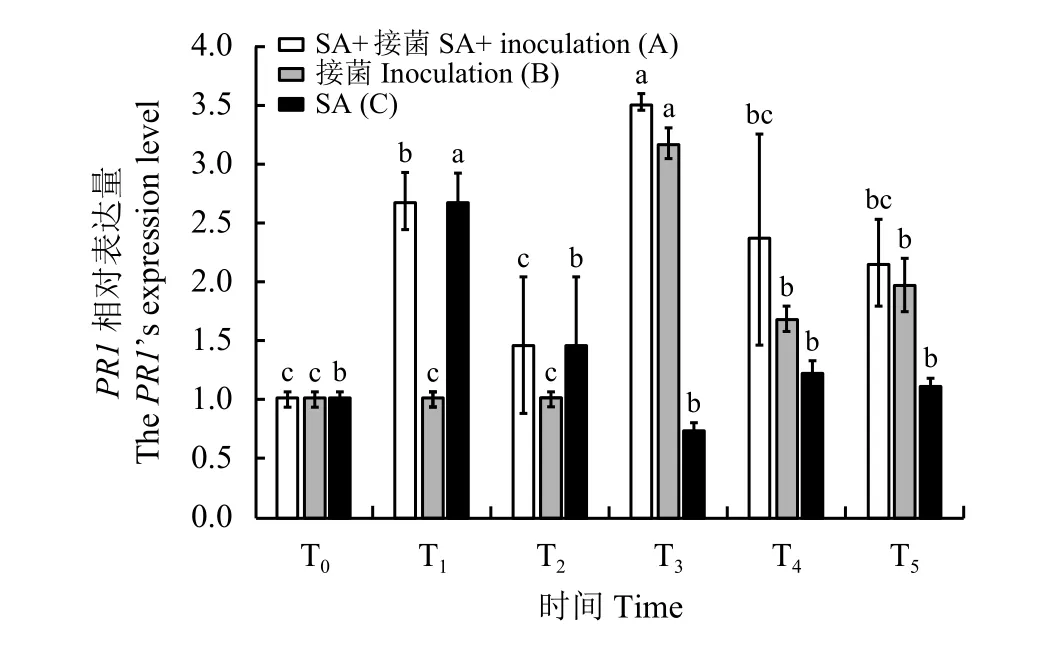
图4 草地早熟禾午夜基因PR1相对表达量Figure 4 The relative expression of PR1 in Kentucky bluegrass ‘Midnight’
A、C处理在T1时,NPR1基因相对表达量显著提高 (P< 0.05),达到 T0的 2 倍 (图5)。之后,C处理NPR1基因相对表达量逐渐降低,且在T3、T5时显著 (P< 0.05)低于 T0。A 处理NPR1基因相对表达量呈显著(P< 0.05)上升趋势,并在T5时达到最大,为喷施前(T0)的5.2倍。B处理在接菌后NPR1基因表达量呈逐渐下降趋势,在T4、T5时NPR1基因相对表达量分别为接种前(T2)的40%和70%,差异显著(P< 0.05),说明在立枯丝核菌侵染下,午夜NPR1基因表达量下降,导致病害严重发生。可见NPR1基因的表达与植物抗病性提高密切相关,A处理在SA喷施后接种病原菌,NPR1基因的表达持续上升,说明外施SA在一定程度上提高了植物的抗病性。
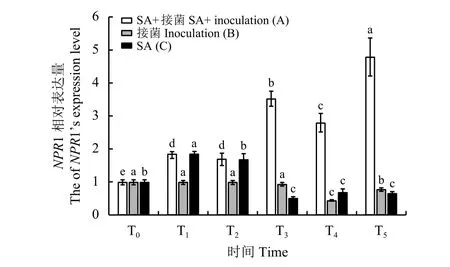
图5 草地早熟禾午夜NPR1基因相对表达量Figure 5 The relative expression of NPR1 in Kentucky bluegrass ‘Midnight’
3 讨论和结论
当植物遭遇逆境胁迫时,体内会产生大量自由基等有毒物质,损害植物细胞结构以致植物死亡[19]。SA是诱导植物抗性的信号分子,可通过诱导植物相关抗病基因表达量的提高使植物体产生系统获得性抗性。抗病基因和防卫反应基因是植物中参与抗性反应的两大类基因,其中病程相关蛋白(PRP)是防卫反应基因编码的产物,在植物抗病性和系统获得抗性中发挥着重要作用[20]。PR1就是17类PRP中的一类[21],在多种植物中已有报道,例如王坤等[22]研究表明PR1基因是小麦(Triticum aestivum)病程相关蛋白基因,本研究也得出在立枯丝核菌侵染的情况下,PR1基因相对表达量持续显著升高 (P< 0.05)。Cao 等[23]研究表明NPR1基因表达与植物抗病性提高有关,缺失NPR1基因的拟南芥对病原微生物感染的敏感性提高,本研究也得出午夜喷施SA并接种立枯丝核菌后,NPR1基因相对表达量持续显著升高 (P< 0.05)。
综上,喷施SA后接种立枯丝核菌会使PR1、NPR1基因持续高表达,对病害具有一定的抗性,在草地早熟禾抵抗病害胁迫中起到积极作用。
