青藏高原东北缘3种灌木数量性状、龄级结构及自疏特征
2019-06-04徐文兵瓦勒塔胡廷花于应文
徐文兵,赵 敏,瓦勒塔,施 颖,胡廷花,于应文
(兰州大学草地农业生态系统国家重点实验室 / 兰州大学农业农村部草牧业创新重点实验室 /兰州大学草地农业科技学院,甘肃 兰州 730020)
高寒灌丛是青藏高原高寒草甸生态系统的重要组成部分,其由耐寒的中生或旱生灌木为优势种而形成的一类植被类型,广泛分布于青藏高原东部海拔 3 200 - 4 500 m 的山地阴坡以及潮湿的滩地,灌丛下生长着藏系家畜喜食的牧草,是青藏高原主要的夏秋牧场[1-3];其在青藏高原生态环境保护、源涵养及藏民族地方经济建设中起着重要作用[4]。
植物种群数量性状(包括盖度、高度、密度、生物量、龄级及自疏法则等)是群落最基本特征,其与群落生产力、生态系统组成、结构和功能等有紧密联系,在生态系统的稳定性及群落演替等方面扮演着重要作用[5]。生物量是植物种群数量特征的核心内容,一定程度上反映植物种对环境的适应能力及其在群落中的地位和作用[5-6]。进行青藏高原高寒灌丛灌木地上生物量分析,探究优势灌木对高寒环境的适应性特征,对于高寒灌丛生态系统稳定性维持机制的深入揭示具有重要意义。通常,灌木地上生物量测定用剪割法,虽客观直接,但该法是一种破坏性方法,不仅繁琐、费时,还存在不确定性,即所测定样本量和均匀性较难满足统计学要求,故在脆弱的青藏高原高寒灌丛灌木测定中使用该法不适合[7]。同时,由于灌木比草本高大,且在水平分布上具明显不连续和不均匀性,故不能用草本样方收获法测定其地上生物量[8]。因此,采用灌木数量性状如冠幅直径、分枝数、株高等,构建高寒灌丛灌木地上生物量适宜估测模型,不仅对高寒灌丛破坏性极小,也是高寒灌丛灌木生物量精准估测的有效手段[9]。
种群年龄结构和自疏特征是植物生态学研究又一主要内容,种群年龄结构是指不同年龄组的个体在种群中的占比或者配置状况,研究种群年龄结构对于分析种群稳定性及动态变化具有重要价值,其研究对象多为乔木,草本及灌木中则研究较少[10]。种群年龄结构不仅反映种群的稳定性和数量动态特征,还反映种群与环境间的相互关系及其在群落中的地位和作用[11]。优势种群在群落中的重要性和地位决定群落发展趋势及植被构成,影响群落稳定性和生物多样性[12]。种群自疏(selfthinning)特征是指生长于较高密度的种群个体植株,因密度制约,种群内个体为获取各生态因子(如水、光、肥等)发生竞争而逐渐死亡,使种群个体数降低,直至达到种群数目平衡,这是植物种群对环境的资源的一种适应性表现[13-14]。因此,进行植物种群年龄结构和自疏特征研究,可以一定程度揭示植物种群的演变动态和对环境的适应性特征。
以往学者对高寒灌丛的研究集中于群落土壤理化特征、群落结构特征、植被类型、物种多样性和生物量关系及CO2通量等方面[15-19]。也有学者研究了高寒灌丛中金露梅(Potentilla fruticosa)、小叶锦鸡儿(Caragana microphyilla)、裂香杜鹃(Rhododendron anthopogonoides)等物种的地上生物量估测模型[17, 20];但有关青藏高原高寒灌丛主要优势灌木的数量特征、年龄结构及种群自疏现象的整合研究相对缺乏。因此,本研究以青藏高原东北缘高寒灌丛广为分布的灌木优势种金露梅、高山柳(Salix cupularis)和头花杜鹃(Rhododendron capitatum)为对象,通过其株丛数量特征的定量测定,对其数量特征关系、株丛年龄结构和自疏特征进行分析,构建3种灌木株丛生物量估测模型,确定3种灌木株丛年龄结构模式及分枝数与分枝重之间是否存在自疏关系,以揭示高寒灌丛主要灌木对高寒生境的适应性机制。
1 材料与方法
1.1 试验区自然概况
研究区位于青藏高原东北缘甘肃省天祝藏族自治县抓喜秀龙乡金强河区域的高寒灌丛草地,地理坐标为 102°40′-102°47' E,37°11′-37°14′ N,海拔 2 920~3 500 m。年均气温-0.1 ℃,极端最高气温26 ℃,极端最低气温-30 ℃,7月均温12.7 ℃,> 0 ℃ 年积温 1 380 ℃·d;年降水量 266~630 mm,主要集中于7-9月,全年无绝对无霜期,仅分冷热两季,冷季长达7个月,植物生长季为120~140 d[21]。高寒灌丛类型主要有金露梅灌丛、高山柳灌丛和杜鹃灌丛等。高寒灌丛占该地区草地总面积的60%以上,主要作为藏系家畜的夏季放牧地,放牧时间占全年的1/3[22]。
1.2 样地设置
2017年7月,在研究区分别设置以金露梅、高山柳和头花杜鹃为灌木优势种的面积约100 m ×100 m的灌丛样地各1块,作为3种灌木株丛特征测定样地。其中,金露梅灌丛的灌木盖度为35%~49%,灌木优势种为金露梅,伴生种有高山柳和高山绣线菊(Spiraea alpina)等。草层盖度30%~65%,主要草本为垂穗披碱草(Elymus nutans)、珠芽蓼(Polygonum viviparum)、矮嵩草(Kobresia humilis)、线叶嵩草(K. capillifolia)、草地早熟禾(Poa pratensis)、甘肃马先蒿(Pedicularis kansuensis)、乳白香青(Anaphalis lactea)等。高山柳灌丛的灌木盖度为30%~42%,灌木优势种为高山柳,伴生种有高山柳和高山绣线菊等。草层盖度35%~53%,主要草本为矮嵩草、高山嵩草(K. pygmaea)、线叶嵩草、高山紫菀(Aster alpinus)、矮火绒草(Leontopodium nanum)等。头花杜鹃灌丛的灌木盖度为30%~45%,灌木优势种为头花杜鹃,伴生种有百里香杜鹃(Rhododendron thymifolium)、高山柳、鬼箭锦鸡儿(Caragana jubata)等。草层盖度12%~25%,主要草本为线叶嵩草、珠芽蓼、黑褐苔草(Carex alrofusca)等。草本层不明显,但灌丛内苔藓层发达,苔藓盖度 > 90%。
1.3 测定方法
在已设置的金露梅、高山柳和头花杜鹃灌丛地上,先随机设置 10 m × 10 m 的亚样地 3~4 块,再分别测各个亚样地内3种灌木的株丛高度、冠幅直径、分枝数、单株重(植物地上部分)和分枝重(植物地上部分);对于金露梅灌丛,仅测定其亚样地内的所有金露梅株丛数量特征,高山柳和头花杜鹃种群数量特征测定亦是如此。具体测定时,因不同灌木分布密度不同,故金露梅、高山柳和头花杜鹃实际分别测定了102株、105株和123株。依据所测定3种灌木株丛的高度、冠幅直径和株丛分枝数,并结合3种灌木自身生长特性及指标的统一性和可比性,对其年龄结构进行等级划分。在株丛高度划分中,金露梅、高山柳和头花杜鹃分别从5、10和5 cm起,均按10 cm的级差分级统计;在株丛冠幅直径和分枝数划分中,3种灌木均从10 cm起,均按10 cm级差分级统计;根据3种灌木在不同生长层次的植株密度计算各生长层次的比例及频度,并用横柱的高低位置表示由幼年到老年的不同龄组,横柱宽度表示各龄组个体数及其所占百分比,绘制年龄结构图。株高年龄分级:金露梅 30~40 cm 幼年期,40~55 cm 中年期,55~65 cm老年期;高山柳60~90 cm幼年期,90~120 cm中年期,120~140 cm老年期;头花杜鹃 50~60 cm幼年期,60~70 cm中年期,70~80 cm老年期。冠幅直径年龄分级:金露梅30~60 cm幼年期,60~90 cm 中年期,90~110 cm 老年期;高山柳 45~105 cm幼年期,105~145 cm中年期,145~205 cm老年期;头花杜鹃 20~40 cm 幼年期,40~60 cm 中年期,60~80 cm老年期。分枝数年龄分级:金露梅和头花杜鹃在0~60个属幼年期,60~100个属中年期,100~140个属老年期;高山柳株丛在10~70个属幼年期,70~130个属中年期,130~170个属老年期。
1.4 数据处理
用Excel进行数据处理及制图。应用SPSS16.0中的Person相关性,对每种灌木单株重与其高度、冠幅直径和分枝数进行相关性分析。以每种灌木的株高、冠幅直径和分枝数为自变量,采用SPSS16.0中的多元线性逐步回归方法,构建每种灌木的地上部分单株重估测模型。以每种灌木的单株分枝数和单株重数据为基础,先计算每种灌木每株的个体枝条重(等于单株重除以其株丛分枝数),并对每种灌木每个株丛的个体枝条重和株丛分枝数数据进行自然对数标准化处理后,采用SPSS 19.0中的一元线性回归方法,分析3种灌木个体枝条重和株丛分枝数之间的回归关系。
2 结果与分析
2.1 3种灌木株丛数量性状
3种灌木5个数量性状中,株高和冠幅直径的变异度相对较小,分枝重变异度居中,株丛分枝数和单株重较大(表1)。说明株高和冠幅直径的稳定性较强,分枝数和单株重的稳定性较弱。
2.2 3种灌木单株重估测模型
3种灌木的地上单株重与其株高、冠幅直径、分枝数均极显著相关,其相关性为冠幅直径 > 分枝数 > 株高(表2)。说明,株高、冠幅直径和分枝数可以作为3种灌木单株重估测模型指标。据此,基于株高、冠幅直径和分枝数构建的3种灌木单株重多元线性回归方程结果显示,金露梅单株重可用其与冠幅直径之间的一元线性模型很好地估测,高山柳单株重可用其与冠幅直径、分枝数和株高之间的一元线性模型、二元线性模型和三元线性模型很好地估测,头花杜鹃单株重可用其与冠幅直径和分枝数之间的一元线性模型和二元线性模型很好地估测(表3)。说明,冠幅直径可作为3种灌木单株重估测模型最适宜指标。
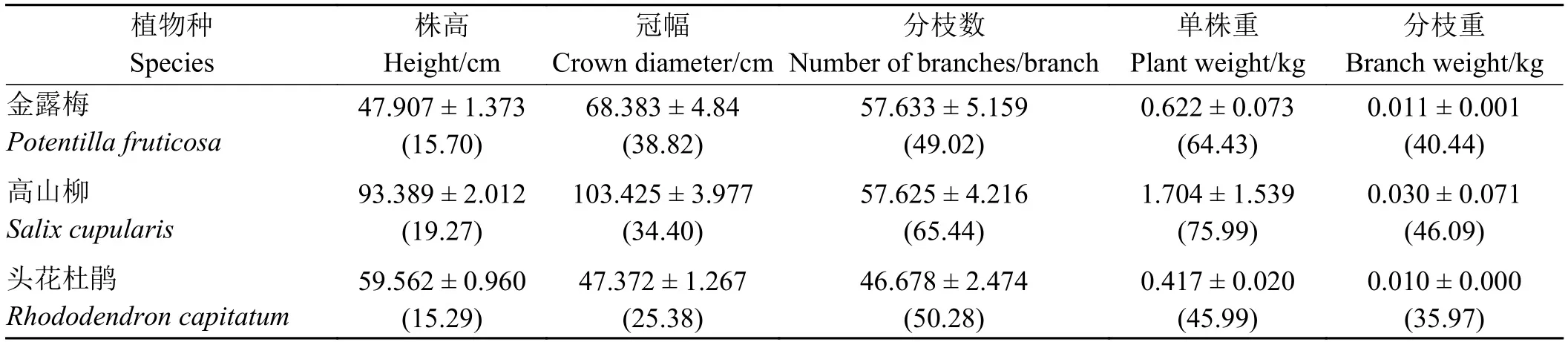
表1 金露梅、高山柳和头花杜鹃株丛数量性状Table 1 The quantitative traits of Potentilla fruticosa, Salix cupularis and Rhododendron capitatum

表2 金露梅、高山柳和头花杜鹃的单株重与各自株高、冠幅直径和分枝数的相关性Table 2 The relationship between plant weight and height, crown diameter and number of branches for Potentilla fruticosa,Salix cupularis and Rhododendron capitatum

表3 金露梅、高山柳和头花杜鹃单株重估测模型Table 3 The estimated model of plant weight for Potentilla fruticosa, Salix cupularis and Rhododendron capitatum
2.3 3种灌木年龄结构
金露梅、高山柳和头花杜鹃的株丛高度分级的年龄结构,除金露梅的高度、冠幅和分枝数的年龄结构基部和顶部较窄、中部较宽外,其余都表现为中部最宽,基部宽于顶部(图1)。说明,金露梅灌丛种群幼年个体数量少,成年个体储备丰富,老年个体数量少,表现为稳定型种群结构;高山柳和头花杜鹃灌丛幼年个体数量充足,成年个体丰富,老年个体数量少,表现为增长型种群结构。
2.4 3种灌木自疏特征
金露梅和高山柳的株丛的个体分枝重与各自株丛分枝数之间无显著相关性,头花杜鹃个体分枝重与其株丛分枝数之间呈极显著相关(表4)。说明,头花杜鹃种群存在明显自疏特征。

表4 金露梅、高山柳和头花杜鹃的个体分枝重与分枝数自疏关系Table 4 The self-thining relationship between branch weight (BW) and number of branches (NB) for Potentilla fruticosa, Salix cupularis and Rhododendron capitatum
3 讨论与结论
利用多元线性逐步回归方法建立灌木地上生物量估测模型时,不仅减弱以原始的直接收获法带来的繁琐和破坏性,而且大大减少工作量,并提高测量精度[7]。本研究得知,二元和多元线性模型建立的灌木地上生物量估测模型比一元线性模型的生物量准确,其原因是灌木地上总生物量等于地上各部分器官生物量之和,自变量越多就越能体现灌木地上总生物量。由于灌木地上生物量估测模型中自变量的选择与其自身形态特征密切相关,同种灌木最优生物量估测模型因其自变量选择不同而产生差异,往往单个变量所模拟的植物种群生物量回归方程不理想;对同一灌木来说,通过不同变量的适当组合才能得到较准确的灌木地上生物量估测模型[7]。
本研究发现,冠幅直径是金露梅、高山柳和头花杜鹃种群单株重估测模型关键指标,基于冠幅直径,可建立3种灌木的株丛地上生物量较理想估测模型;其原因是冠幅直径是一个测量误差较小、估测模型拟合效果较好的变量,这与张贞明和韩天虎[1]的研究结果类似。同时,本研究也发现,3种灌木的冠幅直径具明显差异,其原因是头花杜鹃个体分枝高度较整齐,株丛个体数分布精密,整体呈近圆锥或圆柱体[23],故采用冠幅直径和分枝数组合建立的其地上生物量估测模型较优;高山柳株丛个体分枝数高度不整齐,其下分布大量家畜喜食草本,且家畜对高山柳亦存在一定程度采食,使其株丛冠幅直径和高度变化较大,故采用高度、冠幅直径和分枝数组合建立的其地上生物量估测模型较优;而金露梅由于相对低矮,分枝重较少,且家畜干扰小,故采用冠幅直径建立了其最优生物量估测模型。地域及灌木种类不同,造成灌木地上生物量估测模型自变量的选择不同,从而本研究所建立的3种灌木地上生物量估测模型,是否适宜其他灌木或其他生境的相同灌木,还有待进一步证实。
本研究中,金露梅表现为稳定型种群年龄结构,高山柳和头花杜鹃表现为增长型种群年龄结构,这不仅与3种灌木对生境长期适应性和养分资源的利用方式及其对放牧干扰的强度不同有关[24],还与金露梅根系在地下长距离横走分布,根系分布范围大,且部分位点常长出新植株,头花杜鹃根系根幅小且密集于地表面,高山柳根系分布较深但根幅分布范围较小等有关。因此,在科学管理和有效保护高寒灌丛生态系统方面,对于年龄幼龄数量较少、中龄级植株数量较多的高山柳和头花杜鹃来说,应减少人为干扰和牲畜采食,使其逐渐发育为幼龄级植株数量较多、老龄级植株数量较少且具增长趋势的稳定群落;对于生长稳定的金露梅灌丛应继续保护,使其幼枝得以顺利生长,以自我恢复,保持种群长期稳定性[25]。
植物种群自疏的实质,是由种群个体间由于密度过大,数量较多而对有限资源的竞争导致的[13]。本研究中,金露梅和高山柳均不存在明显自疏特征现象,而头花杜鹃存在明显自疏特征现象。一方面是由于头花杜鹃植株相对低矮,冠幅较小,形态近圆锥或圆柱体[23],其灌丛主要伴生种为线叶嵩草、珠芽蓼、黑褐苔草等,草本层不明显,灌丛内苔藓层发达,为夺取资源和养分,进而加剧其与其他植物种之间的竞争,而高山柳与金露梅植株相对高大,稀疏,株丛冠幅较大,对其下层枝条无明显遮盖作用,而且对光、水、养分等资源的竞争能力也受到限制,因此减弱了灌木种间的相互竞争[26];另一方面是由于在相同面积的样地里头花杜鹃的密度比高山柳和金露梅的大造成的。在密度制约下,头花杜鹃种群内个体趋于死亡,数量减少直至达到新的平衡,发生种群内自疏现象。此外,植物自疏规律及其机制问题一直以来都是种群生态学热点的讨论问题,生态系统是一个能够自我调节和自我适应的高度复杂系统,这种复杂的自调节和适应过程是造成植物自然稀疏的原因之一[14]。本研究仅对特定区域3种灌木的自疏法则进行了一定程度分析,而植物自疏规律及对外界环境的适应机制等有待进一步深入研究。
