Occurrence, properties and biological significance of pyroglutamyl peptides derived from different food sources
2019-05-26BehzadGazmeRuthBoachieApollinaireTsopmoChiuikeUdenigwe
Behzad Gazme, Ruth T. Boachie, Apollinaire Tsopmo, Chiuike C. Udenigwe
a School of Nutrition Sciences, Faculty of Health Sciences, 415 Smyth Road, University of Ottawa, Ottawa, Ontario, K1H 8L1, Canada
b Food Science and Nutrition Program, Department of Chemistry, Carleton University, 1125 Colonel By Drive, Ottawa, Ontario K1S 5B6, Canada
c Department of Chemistry and Biomolecular Sciences, University of Ottawa, Ottawa, Ontario K1N 6N5, Canada
Keywords:
A B S T R A C T
1. Pyroglumate formation in food
Pyroglutamic acid (5-oxo-proline; pGlu) or pyrrolidone carboxylic acid is a cyclic non-proteinogenic amino acid, containing a γ-lactam ring, that is produced from glutamine or glutamic acid by deamidation or dehydration, respectively [1,2]. Proteins and peptides that have the glutaminyl or glutamyl residues at their N-terminal tend to be transformed into pGlu-containing protein or peptide by intramolecular cyclization, which can occur via non-enzymatic and enzymatic processes [2]. Although the nonenzymatic reactions can occur spontaneously to form a new C–N bond through nucleophilic reaction of the α-amino group to the γ-carboxyl group [3,4], most of the cyclization process is expedited by physical conditions such as heat and pressure [3]. The formation of pGlu via deamination of glutamine is favored under alkaline conditions than in neutral and acid environments [5]. This is because lowering the pH leads to increasing protonation of the amino group,therefore inhibiting the nucleophilic reaction on the side chain amide group. The chemical equation for the formation of pGlu residue in N-terminal glutamine-containing peptides is shown in Fig.1. The reaction proceeds via a nucleophilic reaction of the Nterminal amine. Decreasing the reaction pH should inhibit the cyclic intermediate state because of increasing protonation of the amino group. Consequently, this will lead to a reduction in the amount of pGlu formed from N-terminal glutamine-containing peptides.Moreover, Schneider et al. [6] reported that high-pressure treatment reduced or increased pGlu formation when the sample was dissolved in phosphate buffer (pH 7) and Tris buffer (pH 7), respectively. This difference was explained by the decrease in pH of the phosphate buffer, but not that of Tris buffer, under high pressure[6].
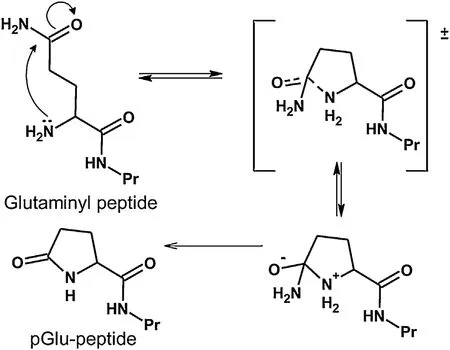
Fig.1. Formation of pyroglutaminyl (pGlu) peptide via cyclization of the side chain of glutamine at the N-terminal position. The reaction can be catalyzed by heat,glutamyl-peptide cyclotransferase or glutaminyl cyclase.
The spontaneous cyclization rate in pGlu formation is higher for glutamine than for glutamic acid. With a change of pH from acidic to neutral, there is a significant decrease in the cyclization rate of glutamic acid, but glutamine cyclization is not significantly affected by pH changes [2]. This may mean that both protonated γcarboxyl group and deprotonated α-amino group are needed for the nucleophilic reaction. The rate constants for spontaneous cyclization are 10−6s−1and 10−9s−1for glutamine and glutamic acid,respectively, in aqueous solution at pH 6.5 [2]. On the other hand,the formation of pGlu residues in peptides can occur in the presence of glutamyl-peptide cyclotransferase or glutaminyl cyclase,which catalyzes the cyclization of both glutaminyl and glutamyl residues, and with glutamyl cyclase that leads to glutamyl cyclization [7,8]. Both cyclases exist in plants and animals and are most abundant in mammalian secretory and neuronal tissues such as the pituitary gland and hypothalamus [9–11]. At these locations,the cyclases play an important role in the maturation of hormones and cytokines such as gonadotropin-releasing hormone and thyrotropin-releasing hormone [7,11]. Using buffers in vitro, optimum pH values of 8 and 6 were reported for mammalian and plant glutaminyl cyclase activities during glutamine and glutamic acid cyclization, respectively [7]. By comparing native-crude and boiled bovine pituitary extract, Wolfgang et al. [4] also showed that the glutaminyl cyclase enzymatic reaction rate for glutaminyl peptides was 70 times higher than the rate for non-enzymatic cyclization [4].
Various studies have reported that enzymes released from bacteria during cheese ripening may play a role in the formation of free pyroglutamic acid and pGlu peptides in cheese products. The amount of pyroglutamic acid or pGlu peptide production depends on the starter microflora rather than the raw milk [10,12–14]. For instance, cell lysates obtained from cultures of Lactobacillus helveticus, L. delbrueckii subsp. bulgaricus, L. delbrueckii subsp. lactis, and Streptococcus thermophilus were found to independently cyclize glutamine to pyroglutamic acid. It is possible that enzymes released during cheese ripening, due to bacterial lysis, remain active in the cheese environment [12,13], and that some of the enzymes are responsible for pGlu formation. The same phenomenon was reported to occur in dry-cured ham due to endogenous enzymes already present in the muscle tissue [15]. Furthermore, pGlu dipeptides are produced during rice fermentation in the production of Japanese rice wine “sake” after rice protein digestion by proteases[3]. Mold (Aspergillus oryzae) is reported to be the major source of protease for pGlu peptide formation in sake, and yeast (Saccharomyces cerevisiae) was thought to have no particular effect in the process [3].
2. Occurrence of pyroglutamyl peptides in food
The first report of the presence of pGlu peptides in food was in the 1970s. Altamura et al. [16] reported the presence of pGlucontaining dipeptides pGlu-Thr, pGlu-Asp, pGlu-Val, pGlu-Leu,pGlu-Phe, pGlu-Gly, pGlu-Ala, pGlu-Glu and pGlu-Pro in aciddigested edible mushrooms (Agaricus campestris). Moreover, wheat gluten is rich in glutenin and gliadin with a high amount of glutaminyl residues at the N-terminal, which can be converted to pGlu residues during processing [17,18]. For instance, some studies have reported that the use of heat during the hydrolysis of wheat gluten hydrolysates produced pGlu residues [18–20]. Sato et al.[18] was the first group to report the presence of pGlu peptides in wheat gluten hydrolysates. A further study demonstrated in vitro that some of the pGlu peptides in the wheat gluten hydrolysate like pGlu-Asn-Pro-Gln, pGlu-Gln-Gln-Pro-Gln, pGlu-Gln-Pro-Gln,pGlu-Gln-Pro-Gly-Gln-Gly-Gln, pGlu-Gln, pGlu-Gln-Pro, pGlu-Ile-Pro-Gln, pGlu-Ile-Pro, pGlu-Gln-Pro-Leu, pGlu-Gln-Phe-Pro-Gln,pGlu-Ser-Phe-Pro-Gln, pGlu-Phe-Pro-Gln, and pGlu-Gln-Pro-Pro-Phe-Ser were resistant to digestion with pepsin and pancreatin[19]. Other works reported the occurrence of pGlu-Leu and pGlu-Gln-Leu in the subtilisin digests of glutenin and gliadin after heat treatment [20], whereas pGlu-Ile, pGlu-Leu and pGlu-Gln, pGlu-Gln-Gln were found in the acidic fraction of a gluten hydrolysates produced with Aspergillus oryzae protease [21]. Notably, it is not apparent if the N-terminal cyclization occurred as a result of enzymatic activity, or heat treatment, or a combination of these factors,during the production of these wheat protein hydrolysates.
Twenty eight pGlu peptides were also identified in the Japanese rice wine, including pGlu-Asn-Ile-Asp-Asn-Pro, pGlu-Asn-Ile,pGlu-Val, pGlu-Leu-Trp, pGlu-Val-Ala, pGlu-Val-Pro, pGlu-Val-Val,pGlu-Asn-Phe, pGlu-Leu, pGlu-Gln, pGlu-Ser-Gln, pGlu-Met, pGlu-Gly-Gln, pGlu-Tyr, pGlu-Phe, pGlu-Asn, pGlu-Ser, pGlu-Gly and pGlu-Ala; notably, dipeptides pGlu-Leu and pGlu-Gln were the major constituents [3,22]. Furthermore, pGlu-Pro, pGlu-Ile and pGlu-Leu were identified in dry-cured ham, and pGlu-Ile, pGlu-Leu and pGlu-Phe in Parmigiano-Reggiano cheese [15,23]. So far,nine pGlu dipeptides: pGlu-Gly, pGlu-Gln [24], pGlu-Asp, pGlu-Glu,pGlu-Pro, pGlu-Val, pGlu-Ile, pGlu-Leu and pGlu-Phe have been reported in fermented soy sauce [25]. Using a peptidomic approach,Yao and Udenigwe [26] found that there were about one hundred pGlu modified peptides in a pepsin and pancreatic potato protein hydrolysate and all have more residues than those reported from the works described above. There is evidence that pGlu-γ3-casein can be formed spontaneously in cheese, especially in hard and extra-hard cheeses where the role of the cyclase enzymes ought to be negligible [9]. In general, pGlu peptides with 2–20 amino acid residues have been identified in different processed foods products,with the highest frequency of occurrence reported for dipeptides and tripeptides (Fig.2). Overall, pGlu-Leu has been the most frequently reported pGlu peptide in the various foods.
3. Quantification of pGlu peptides in food
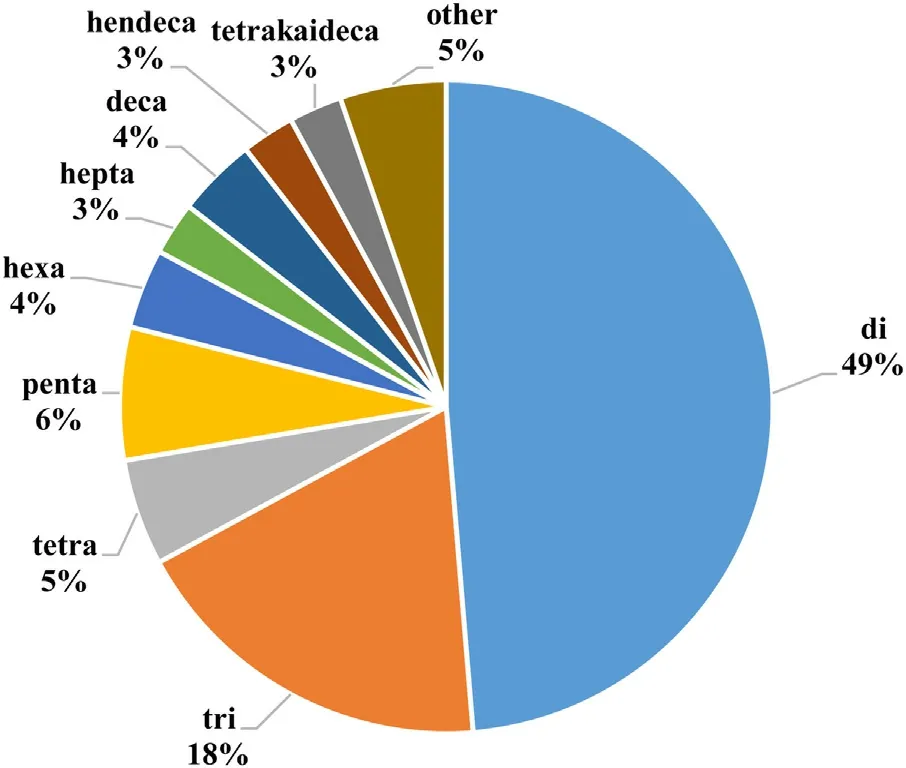
Fig.2. Percentage of sequenced pGlu peptides derived from food sources based on the number of amino acid residues.
Different approaches have been used to quantify pGlu peptides in foods. In some early studies, the content of pGlu residues in peptides was quantified by measuring the amount of free pyroglutamic acid content before and after digestion of the food sample with pyroglutamate aminopeptidase. This enzyme specifically releases pyroglutamic acid from the N-terminal of the pGlu peptides[17,18,27]. After digestion, the isolation of free pyroglutamic acid from other amino acids and peptides is then carried out using methods such as solid-phase extraction [27]. The amount of pyroglutamic acid liberated by the enzymatic digestion is then calculated by subtracting the amount of initial pyroglutamic acid from total pyroglutamic acid present in the pyroglutamate aminopeptidase digest. For example, Higaki-Sato et al. [6] used enzymatic digestion for quantification of pGlu peptides in wheat gluten hydrolysates and its acidic fraction. After pyroglutamate aminopeptidase digestion, they separated pyroglutamic acid from other amino acids and peptides by a strong cation-exchange resin followed by ionexclusion chromatography and suppressed conductivity for analyte detection. The advantage of this method is that it can be conducted with simple equipment, but it is not suitable for detection and quantification of pGlu at sub-micromolar levels. Furthermore,it needs sample fractionation with reversed phase chromatography or additional ion-exclusion chromatography [27], which can decrease pGlu recovery potentially leading to incorrect quantification.
Most recent studies have used more direct, sensitive analytical methods based on liquid chromatography coupled with tandem mass spectrometry (LC–MS/MS) to quantify pGlu peptides in protein hydrolysates and fermented food products. For instance, Sforza et al. [23] used LC/ESI-MS for quantification of pGlu peptides in various cheese products after different durations of ripening. For this purpose, the authors used dipeptide Phe-Phe as an internal standard and spiked samples with synthesized N-pGlu-Phe. The relative response factor was obtained for the synthesized dipeptide and used for quantitation of the pGlu peptides [23]. Moreover, Kaneko et al. [24] determined the amount of taste-active pGlu peptides in fermented soy sauce using LC/TOF-MS. Prior to quantification, several fractionation steps including reverse phase-high performance liquid chromatography (RP-HPLC), gel permeation chromatography and hydrophilic interaction chromatography, were used to process the samples [24]. In another example, pGlu-Leu in various commercial sake products were quantified using LC–MS/MS after separating the pGlu peptide from other peptides using strong cation-exchange resin [3]. Similarly, Frerot and Chen [25] separated an acidic fraction of soy sauce by anion-exchange solid-phase extraction, used LC/MS-MS for pGlu peptides quantification, and then chemically synthesized the pGlu peptides for verification of their existence and quantification by selected reaction monitoring[25]. Although fractionation often used before LC–MS/MS analysis is necessary to simplify the matrix and enhance analyte quantification, the multiple processing steps are likely to reduce the amount of pGlu peptide detected.
pGlu oligopeptide ethyl esters present another challenge in that they can interact with metal ions during sample preparation for MS, and their ionization ratios in ESI-MS can be influenced by the food matrix [29]. Therefore, stable isotope dilution analysis method has been used for pretreatment prior to quantifying taste-active ethyl esterified pGlu peptides in sake. For this purpose, deuterated ethyl esterified pGlu peptides were added to sake samples and applied to a preconditioned column. After preparation of the trapped compound, samples were applied to ESI-MS without LC separation [28,29]. This approach will likely reduce sample loss and can be optimized for more accurate quantification of pGlu peptides in different food product matrices.
One of the latest methods used to identify and quantify pGlu peptides is the shotgun-based peptidomics approach. Yao and Udenigwe [26] used this peptidomics approach to identify the occurrence of several post-translational modifications, including pGlu formation, in peptides generated from potato proteins hydrolyzed with pepsin and pancreatin. This process involves first analyzing the peptides using LC–MS/MS followed by peptidomic analysis using a proteomics software, e.g. PEAKS software. Both de novo sequencing and database search strategy were used to identify the pGlu peptides. Consequently, 100 pGlu peptides were identified in the potato protein hydrolysate, 91 of them from Nterminal glutamine residue cyclization and 9 from the cyclization of N-terminal glutamate residues [26]. Although this method is the most comprehensive so far, in terms of the large number of pGlu peptides reported, smaller pGlu peptides with chain length of less than five amino acid residues could not be accurately identified in the study [26]. A similar approach was used by Paolella et al. [15]for identification and quantification of pGlu dipeptides in a peptide fraction (MW < 10 kDa) and gastrointestinal digests of dry-cured ham. In this case, samples were first analyzed by HPLC-ESI-MS/MS and then peptide sequence identification was performed using the bioinformatics resource portal, ExPASy with the FindPept tool, and Proteomics Toolkit web application, using the obtained mass spectra. Table 1 shows the identity, concentration and methods used in the quantification of pGlu peptides obtained from various food sources.
4. Sensory effects of taste-active pGlu peptides
Food taste plays a decisive role in food selection, intake, absorption and digestion, and therefore it affects the nutritional status of consumers and their decision to purchase food products [30].During processing, food taste is altered as a result of spontaneous chemical changes and those resulting from the activity of microorganisms and enzymes. Proteolysis plays an important role in the development of flavor in protein-rich foods such as cheese, meat and fermented foods. During proteolysis in fermentative or pure enzymatic processes, taste-active peptides and amino acids are produced [31]. Peptides play a significant role in imparting umami and bitter tastes to foods (e.g. hydrophobic proline-containing peptides) [32]. Umami is recognized as the fifth basic taste after sweet, bitter, sour, and salty, and is characterized by a savory flavor that is naturally present in many plant- and animal-based foods [33–35]. Glutamic acid is the first known umami compound.Umami is now defined as the taste of monosodium glutamate and 5′-ribonucleotides [36]. Other compounds that impart umami taste on foods include tyrosine, phenylalanine and pGlu peptides[37–39].
Some studies have reported the presence of umami-tasting pGlu peptides in wheat gluten and soy sauce products. Schlichtherle-Cerny and Amadò [31] reported that pGlu peptides from acid-deamidated wheat gluten yielded a glutamate-like, umami taste and their sequences were determined to be pGlu-Pro-Ser,pGlu-Pro, pGlu-Pro-Glu, and pGlu-Pro-Gln [30]. Moreover, pGlu-Gly and pGlu-Gln were identified as some of the umami tasteenhancers in soy sauce at subthreshold concentration of 2.2 mmol/L and sweet taste above threshold concentration of 1.9 mmol/L[24]. In addition, the presence of pGlu peptides that increase the umami taste in Japanese noodle soup (udon-dashi) has also been reported [40]. On the other hand, some pGlu peptides have bitter or astringent taste. Hashizume et al. [41,42] separated five pGlu peptides from sake with some of them having bitter taste,e.g. pGlu-Leu-Phe-Asn-Pro-Ser. Notably, two of the pGlu peptides also have a C-terminal proline residue, which has a hydrophobic side chain known to contribute to bitter taste. Thus, the combination of the C-terminal proline and N-terminal pGlu residues may have a synergistic effect on the bitter taste of the peptides[41,42]. Furthermore, pGlu oligopeptide ethyl esters have bitter taste, which usually have a negative effect on the quality of sake.So far, pGlu-Leu(ethyl) (bitter taste), pGlu-Leu-Phe-Gly-Pro(ethyl),pGlu-Leu-Phe-Asn-Pro(ethyl) (mild taste) as well as two pGlu decapeptides (bitter taste, with recognition thresholds of 8.1 and 3.8 μg/L) have been identified in sake [28,29,42].
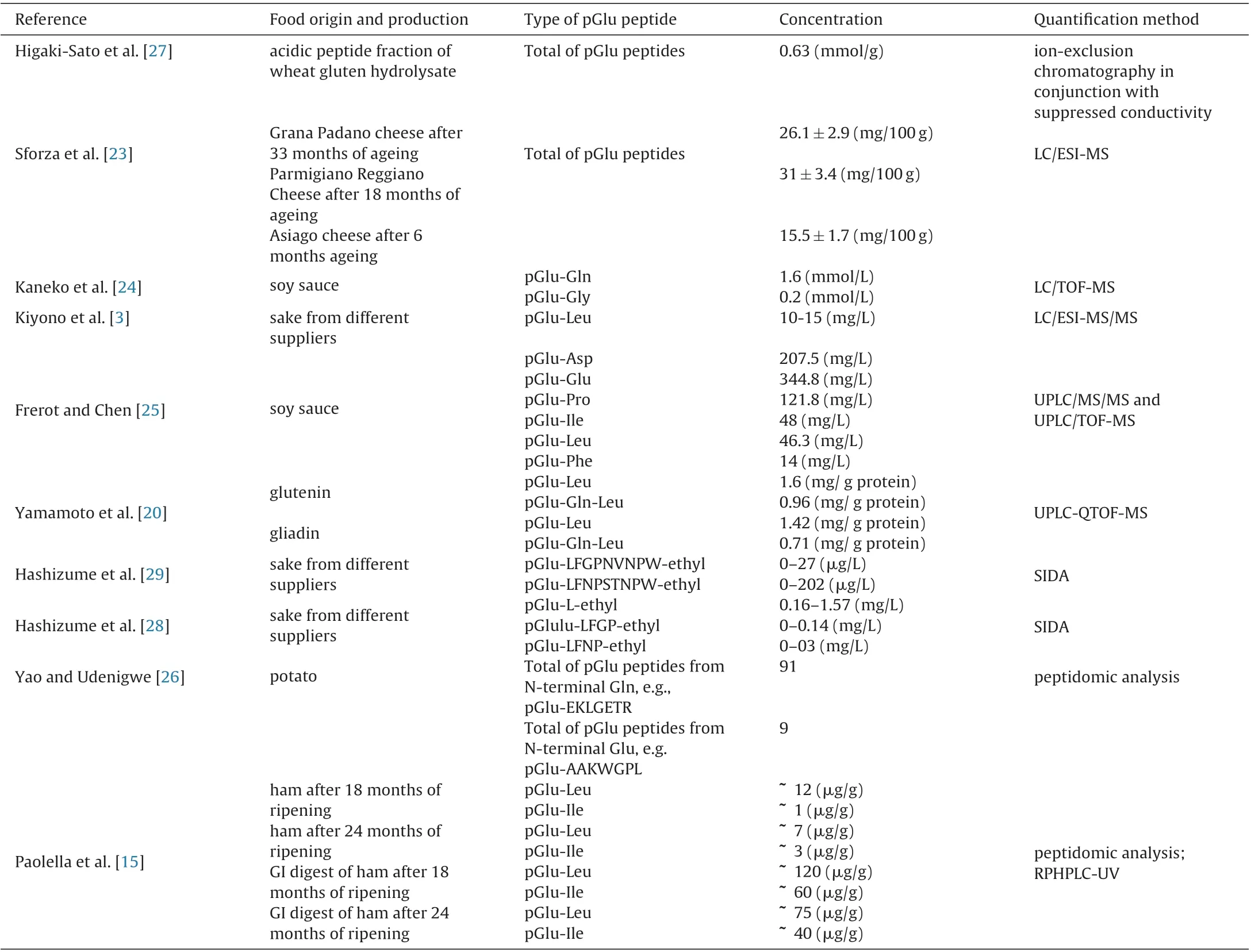
Table 1 Type, concentration and quantification method of pGlu peptides in various food.
Taste is thought to be a key factor for consumer acceptance of peptide-based functional food products [43]. Typically, sweet and umami tastes result in better acceptance in many populations,and bitter taste tend to result in rejection of the food products[44]. As discussed in this section, pGlu peptides have taste properties that may not be accepted by the consumer. Therefore, studies are required to enhance consumer acceptance of pGlu-containing food products. Umami receptors mGluR4 [45], T1R1 + T1R3 [46,47]and mGluR1 [48] have been identified in the taste buds. Bitter compounds bind 25 receptors belonging to a family class G proteincoupled receptors called T2Rs to initiate the bitter taste [49,50]. To date, there is no information about the taste-activation mechanism for pGlu peptides and their respective receptors in taste buds. It is possible to select the appropriate process for debittering of pGlu peptides without changes in their bioactivity or stability through a better understanding of the biological mechanisms, taste receptor and pGlu peptide structures. Some umami peptides are known to exhibit bitter taste after binding bitter taste receptors. Thus, it is likely that the pGlu peptides also induce their taste effects via this mechanism. In addition, due to the structural similarity of some sweet (T1R2 + T1R3) and umami (T1R1 + T1R3) receptors, the specific structure of peptides has a significant effect on their taste[51]. Besides debittering, approaches such as encapsulation and the use of masking compounds can be investigated for suppressing the undesirable taste effect of pGlu peptides in food product formulations.
5. Biological relevance of food-derived pGlu peptides
5.1. Biostability of pGlu peptides
The biological relevance of orally consumed molecules, including peptides, is dependent on their ability to resist gastrointestinal digestion. To date, only a few biostability studies has been reported for pGlu peptides. It has been found that the presence of γlactam or pyrrolidone moiety in pGlu peptides makes them almost resistant to human digestive enzymes. In a study of Sato et al. [18] for example, treatment of an industrial wheat gluten hydrolysate with pepsin and pancreatin showed little changes in glutamine contents and isolated indigestible pGlu peptides using chromatographic separations. Data from that work is an evidence of the biostability of pGlu peptide in the hydrolysate;as expected, further hydrolysis of the sample with pyroglutamyl aminopeptidase resulted in increased concentration of pyroglutamic acid [18]. In a follow up study, the dipeptide pGlu-Leu was detected in the blood of rats after oral ingestion of wheat gluten hydrolysate [21]. In another work, both free and peptide forms of pyroglutamic acid were found in the plasma from the portal blood of rats that ingested a wheat gluten hydrolysate containing several pGlu peptides [27]. An increase in human plasma level of pGlu-Pro, pGlu-Leu-Pro, and pGlu-Gln-Pro was also detected after intake of corn and wheat gluten hydrolysates produced with pancreatin and leucine aminopeptidase [53]. Based on these studies, some peptides containing pGlu moieties are bioavailable and thus can potentially produce activities in biological systems. Unfortunately, the research to date has been limited to di- and tripeptides which can be an indication that larger pGlu peptides are susceptible to cleavage by the digestive enzymes.
5.2. Bioactivities of pGlu peptides in vitro and in vivo
The bioactivity of compounds is determined by their bioavailability and absorption. In the case of peptides, the net charge,the length of sequences and hydrophobicity have been found to be important [26,52]. As discussed in the section above, the formation of γ-lactam ring removes the polar amide group,which then increases the hydrophobicity of pGlu peptides making them likely to be more bioavailable. Bioactivities such as hepatoprotection, antidepressant, and anti-inflammatory activities have been reported for pGlu peptides taken as part of a protein hydrolysate or in pure forms. Orally administered dipeptide pGlu-Leu was reported to reduce acute hepatitis induced with D-galactosamine in mice [21], while the same peptide at a dose of 0.1 mg/kg body weight/day was beneficial against inflammation (i.e. colitis) in mice caused by dextran sulfate sodium as there was less edema and greater population of Bacteroidetes in the colon [54]. The beneficial effect of pGlu-Leu was proposed to occur by stimulating the murine enterocytes to produce bioactive compounds that enhanced the microbiota and then reduced inflammation.
Neurotensin and thyrotropin-releasing hormone are known endogenous pGlu peptides produced in the central nervous system where they activate target proteins to exhibit physiological actions associated with emotional behavior, learning, pain response and metabolism. For this reason, exogenous pGlu peptides have therefore been investigated of possible beneficial effects on the nervous system. In a work by Yamamoto et al. [20], pGlu-Leu and pGlu-Gln-Leu demonstrated antidepressant-like activity by promoting hippocampal neurogenesis via a pathway that was independent of brain-derived neurotrophic factor. They also found that pGlu-Gln-Leu enhanced neurogenesis in the dentate gyrus of the hippocampus in a manner comparable to neuronal proliferation and neurogenesis by antidepressants [20]. The antidepressantlike effect of pGlu-Gln-Leu was observed even at a lower dose(3 mg/kg body weight) compared to the dose of unmodified peptide Gln-Gln-Leu (10 mg/kg body weight), which demonstrates the importance of the lactam ring in the bioactivity. Furthermore,the potency of pGlu-Gln-Leu was observed at a concentration 10-fold lower than that of pGlu-Leu while individually, amino acids pGlu and Leu had no significant effect. This observation emphasizes the importance of a proper peptide sequence for bioactivity.The four peptides pGlu-Leu, pGlu-Val, pGlu-Met, and pGlu-Phe all hindered the release of nitric oxide, tumor necrosis factor-α,and interleukin-6, which are indicators of lipopolysaccharideactivated inflammation in macrophages [55]. Peptides pGlu-Phe and pGlu-Leu had the most suppressing effects on interleukin-6 and nitrate release, respectively. The variation observed could stem from the structural differences between the dipeptides as Leu possesses an isopropyl, Val an isobutyl, and Phe an aromatic group on their side chains, which although different in size can increase hydrophobic interactions with the target biomolecules.
Despite the beneficial effects of small pGlu peptides on brain and inflammation, the formation of γ-lactam ring in large proteins can have deleterious properties. Amyloid-β peptides (Aβ)isolated from brain vascular walls and plaques from Alzheimer’s patients contained pGlu residues (Aβ3pGlu and Aβ11pGlu). Their ability to easily form β-sheets and aggregate in aqueous solutions was attributed to the presence of pGlu residues and the resulting increase in hydrophobicity [56,57]. The aggregation of amyloids is well characterized in Alzheimer’s disease [58].It will be interesting to investigate the ability of food-derived pGlu peptides to cross the blood-brain barrier, and the relationship between their beneficial neuroactivity and the occurrence of Aβ3pGlu and Aβ11pGlu responsible for aggregation of amyloids.
6. Conclusion
During food processing, several factors can lead to structural modification of N-terminal glutaminyl or glutamyl residues to form pGlu peptides. The γ-lactam ring structure makes the peptides to become more resistant to digestive proteases, which may enhance peptide absorption in their native forms. From this, it can be ascertained that health benefits of pGlu peptides could be retained since their forms may not have changed along the chain of food processing to digestion and absorption in target tissues. However,there is a dearth of information on the molecular mechanisms of bioactivity, shelf life in food products, safety, and sensory effects of food-derived pGlu peptides, hence further investigation will be needed to fill these knowledge gaps.
杂志排行

食品科学与人类健康(英文)的其它文章
- Risk assessment of chemical substances of safety concern generated in processed meats
- Seaweed nutraceuticals and their therapeutic role in disease prevention
- Antioxidant activity and total phenolic content of essential oils and extracts of sweet basil (Ocimum basilicum L.) plants
- Antioxidant peptides encrypted in flaxseed proteome: An in silico assessment
- Hyperinsulinemia, cancer and maqui berry: The promise of nutritional supplementation
- Fermentation-enabled wellness foods: A fresh perspective