小麦光呼吸途径电子流分配的模型研究
2019-05-15叶子飘段世华康华靖
叶子飘,段世华,安 婷,康华靖
(1.井冈山大学数理学院,江西吉安 343009; 2.井冈山大学生命科学学院, 江西吉安 343009; 3.温州市农业科学研究院, 浙江温州 325006)
植物的光呼吸是在光照条件下绿色细胞内吸收O2、消耗光合作用产物,同时释放CO2的过程。相对C4植物而言,C3植物表现出较高的光呼吸活性,可消耗总光合作用已固定碳素的20%~50%[1]。所以,该过程显著影响植物光合产物的积累和光合效率的进一步提高,是减少植物光合产物消耗、提高植物高光效研究的重点,也是生物圈中重要的碳代谢途径之一[2-3]。大量文献主要从分子水平上阐述了光呼吸的代谢途径和关键酶的互作关系[4-11],并论述了光呼吸在高光强或其他胁迫环境条件下对植物的光保护作用[10-14 ]。
在植物的光呼吸过程中,当其他消耗电子的途径可以忽略或为常数时,参与碳同化过程的光合电子流(J)可由公式J=[J+8(Ac+Rday) ]/3计算(式中的Ac为净光合速率,J为总的光合电子流,Rday为日呼吸速率),参与光呼吸途径的光合电子流(Jo)则可由公式Jo=2[J-4(Ac+Rday)]/3计算。当植物在确定的环境条件下,其J和Rday的值是确定的,那么由J的表达式可知,分配到碳同化过程的光合电子流随Ac的增加而增加,而Ac对CO2浓度的响应又是非线性的。另外,从Jo的表达式可知,分配到光呼吸过程的Jo随Ac的增加而下降,但Jo对CO2浓度是如何响应则尚不清楚。已有研究结果表明,随CO2浓度的增加,大豆叶片的光呼吸缓慢地增加到最大值,然后下降[15]。理论上其分配到光呼吸路径的光合电子流对CO2浓度的响应应该与光呼吸速率对CO2浓度的响应趋势相同。此外,人们大多只是定性地报道了植物光呼吸速率对CO2浓度的响应问题[11,16-17],定量研究则鲜见报道[12, 15, 18]。植物的光呼吸速率对不同CO2浓度是如何响应的?植物参与光呼吸过程的光合电子流是如何响应CO2浓度的变化?是否分配到光呼吸的光合电子流或光呼吸达到最大值时,光合碳同化也达到最大值?诸如此类的问题尚不清楚。
为此,本研究以小麦Z39-118为材料,探讨小麦叶片光呼吸速率和分配到光呼吸的光合电子流对CO2浓度的响应规律,以期为光呼吸和分配到光呼吸的光合电子流的定量研究提供理论参考和数学工具。
1 材料与方法
1.1 试验材料与方法
试验以小麦Z39-118为供试材料。该材料由中国科学院遗传与发育生物学研究所李振声院士提供。本试验于中国北方黄淮海平原的中国科学院禹城综合试验站(36°40′~37°12′N,116°22′~116°45′E,海拔23.4 m)的试验田上开展。所在地区属于鲁西北地区有代表性的农业中、高产区,主要种植制度为冬小麦-夏玉米一年两熟制。冬小麦为冬性中、晚熟类型,十月初播种,播种前一般翻耕整地,施足基肥,翌年三月浇水追肥;春季冬小麦返青后,四月底至五月初浇孕穗、开花水,六月上旬收割。
1.2 测定指标与方法
数据测量于2013年5月13-19日的晴天进行。此时小麦处于盛花期。随机选长势较为一致的健壮植株,以旗叶作为测量对象[12]。
数据测量时,在自然光下诱导1.5~2 h后,采用开放式气路,设定叶室里的温度为30±1.3 ℃,流速为400 μmol·s-1,空气相对湿度控制在45%~70%,流速为400 μmol·s-1,应用CO2注入系统提供不同的外界CO2梯度(Ca):1 400、1 200、1 000、800、600、400、380、200、150、100、80、50和0 μmol·mol-1;应用Li-6400-40荧光探头提供2 000 μmol·m-2·s-1光合有效辐射强度(PAR),采用自动测量程序(Flr A-Ci curve),同时记录叶片2%O2和21%O2浓度下的气体交换和叶绿素荧光等参数。自动测量时,每次记录最小等待时间为2 min,最大等待时间为3 min,数据记录之前仪器均自动进行参比室和样品室之间的匹配。
1.3 模型简介与数据处理
1.3.1 模型简介
(1)传统计算分配到碳同化和光呼吸光合电子流的模型
参与碳还原的光合电子流[18-20]用J表示为:
J=4(Ac+Rday+R1)
(1)
式中,Ac为净光合速率,R1为光呼吸速率,Rday为日呼吸速率。由于难以直接测量,一般认为Rday是暗呼吸速率(Rd)的一半[21]。
参与光呼吸的光合电子流[18-20, 22]用Jo表示,表达式为:
Jo=8R1
(2)
当其他消耗电子的途径可以忽略或为常数时,则由式(3)和式(4)可知,参与碳同化的光合电子流为:
Jc=1/3[J+8(Ac+Rday)]
(3)
参与光呼吸的光合电子流为:
Jo=2/3[J-4(Ac+Rday)]
(4)
(2)光合电子传递速率对CO2浓度响应模型的构建
光合作用对CO2浓度响应的新模型(模型I)[23]的具体表达式为:
Ac=αc(1-βcC)/(1+γcC)C-R1
(5)
式中,Ac为净光合速率,αc为光合作用对CO2浓度响应曲线的初始斜率,βc和γc为两个与CO2浓度无关的系数,C可以表示为大气CO2浓度(Ca)或胞间CO2浓度(Ci),R1为光呼吸速率(如拟合2% O2浓度下测量的数据,该参数则为日呼吸速率)。利用模型I可得到植物叶片的饱和CO2浓度(Csat,既可以表示为大气饱和CO2浓度Ca-sat,也可以表示为饱和胞间CO2浓度Ci-sat)和最大净光合速率(Ac-max):
与模型I类似,植物叶片的光合电子传递速率对CO2浓度响应模型(模型II)的具体表达式[24]应该为:
J=αec(1-βecC)/(1+γecC)C+Jc0
(6)
式中,J为CO2浓度响应下的电子传递速率,αec为电子传递速率对CO2浓度响应曲线的初始斜率,βec和γec为两个与CO2浓度无关的系数。同样地,模型II中C也可以表示为Ca或Ci。Jc0为CO2浓度等于0 μmol·mol-1时的电子传递速率,该值的大小主要与光呼吸和暗呼吸有关,简称为零点电子传递速率。类似地,利用模型II可得到植物叶片的饱和CO2浓度(Ca-sat)和最大电子传递速率(Jmax):它们分别由

光呼吸速率(R1)对CO2的响应模型参考叶子飘等[15]的数学表达式,即:
R1=α(1-βCa)/(1+γCa)Ca+Rp0
(7)
式中,α是光呼吸速率对CO2浓度的响应曲线的初始斜率,β称为加氧酶系数,γ称为羧化酶系数,Rp0是光强确定而叶室CO2浓度等于0 μmol·mol-1时的光呼吸速率。
由式(7)并结合式(4)可知,分配到光呼吸的光合电子流对CO2浓度的响应为:
Jo=8α(1-βCa)/(1+γCa)Ca+8Rp0
(8)
1.3.2 数据处理
采用“光合计算软件” (http://photosynthetic.sinaapp.com/calc.html,井冈山大学数理学院)对试验数据进行分析。利用模型I拟合光合作用对CO2浓度的响应曲线(Ac-Ca曲线),得到最大净光合速率(Ac-max)及其对应的饱和光强、CO2补偿点等参数。用模型II拟合电子传递速率对CO2的响应曲线(J-Ca曲线),获得最大电子传递速率(Jmax)及其对应的饱和CO2浓度(Ca-sat)和零点电子传递速率(Jc0)等参数。利用2% O2的净光合速率减去21% O2浓度下的净光合速率(ΔAc)获得光呼吸速率对Ca的响应曲线,并用式(7)进行拟合[15],获得最大光呼吸速率(Rl-max)。同时用式(8)拟合光呼吸对CO2浓度的响应曲线得到参与光呼吸的最大光合电子流(Jo-max)及其对应的饱和CO2浓度。采用DPS 7.5 软件对数据进行统计学分析,并利用配对t检验对拟合得到的Jmax与观测值之间差异进行显著性分析。作图由Origin 8.5软件完成,然后用Adobe Illustrator CS 5进行图形处理。用确定系数(r2)表示模型拟合与观测点的符合程度。
2 结果与分析
2.1 光合作用对CO2浓度的响应
在较低的CO2浓度(<600 μmol·mol-1)下小麦的光合作用对CO2浓度几乎为线性响应,在较高CO2浓度下则存在CO2饱和现象(图1a和图1b)。模型I可以很好地拟合小麦在21%和2% O2条件下的Ac-Ca曲线,r2达到了0.995以上。此外,用2%和21% O2条件下小麦的净光合速率之差(ΔAc)可得到小麦的光呼吸速率对CO2的响应曲线(R1-Ca曲线)(图1c)。由图1c可知,Rl随CO2浓度的增加而增加,达到最大值后随之下降,且当CO2浓度等于0 μmol·mol-1时Rl不等于0 μmol·m-2·s-1。
利用模型I和式(7)分别拟合(表1)表明,21%和2% O2条件下小麦光合速率的饱和CO2浓度分别为1 368.27和1 233.32 μmol·mol-1,最大光呼吸速率和外界CO2均为零值时光呼吸速率分别为10.52 和4.27 μmol·m-2·s-1。这些拟合值与观测值非常接近。

图1 小麦在21% O2(a)和2%O2(b)条件下光合作用对CO2浓度的响应,以及光呼吸对CO2的响应(c)

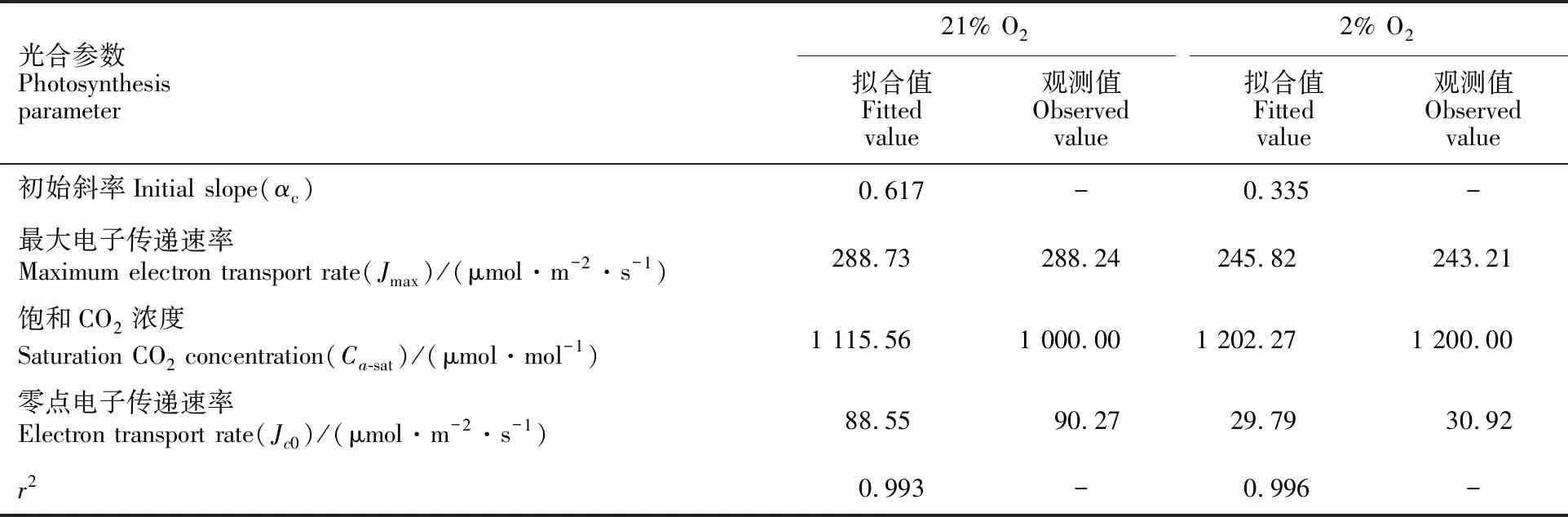
表1 21%O2和2%O2条件下小麦光合作用对CO2浓度响应的光合参数拟合值、最大光呼吸速率等以及其对应的观测数据Table 1 Measured data and fitted results of CO2-response curves of photosynthesis in wheat at 21%O2 and 2%O2, respectively
*:最大光呼吸速率;**:Ca=0 μmol·m-2·s-1时的光呼吸速率。
*:Maximum photorespiratory rate,Rl-cmax;**:Photorespiratory rate atCa=0 μmol·m-2·s-1,Rp0.
2.2 电子传递速率对CO2浓度的响应
在21% O2浓度下,在CO2浓度(<400 μmol·mol-1) 较低时小麦的电子传递速率对CO2浓度几乎为线性响应,当CO2浓度为1 000 μmol·mol-1时小麦的电子传递速率存在CO2饱和现象(图1a);在2% O2浓度下,CO2浓度在0 21%和2% O2条件下小麦最大电子传递速率分别为288.73和245.82 μmol·m-2·s-1,其对应的饱和CO2浓度分别为1 115.56和1 202.27 μmol·mol-1(表2),与观测值均较为接近。 图2 小麦在21% O2(a)和2%O2(b)条件下光合电子传递速率对CO2浓度的响应 表2 21%O2和2%O2条件下小麦电子传递速率对CO2浓度响应的光合参数拟合值以及其对应的观测数据Table 2 Measured data and fitted results of CO2-response curves of electron transport rate in wheat at 21%O2 and 2%O2, respectively 由图3a可知,由公式(4)计算得到的小麦分配到光呼吸的电子传递速率在较低的CO2浓度(约165 μmol·mol-1)下达到最大值,然后随CO2浓度的增加而快速下降;由图3b可知,由公式(8)计算得到的小麦分配到光呼吸的电子传递速率在较高的CO2浓度(约600 μmol·mol-1)下达到最大值,然后随CO2浓度的增加而较快下降。模型II可以较好地拟合由公式(4)和公式(8)计算得到的Jo-Ca曲线,r2分别达到了0.964和0.934。 利用模型II拟合给出在21% O2条件下小麦分配到光呼吸的最大电子传递速率值分别为86.93和84.17 μmol·m-2·s-1,均与观测值(89.12 μmol·m-2·s-1)较为接近(P>0.05);与最大电子传递速率值相对应的饱和CO2浓度分别为164.82和589.42 μmol·mol-1,前者远低于观测值(600 μmol·mol-1)(表3)。 图3 用式(4)和式(8)分别计算分配到光呼吸(a, b)的电子传递速率对CO2的响应 表3 模型II拟合21%O2条件下小麦电子传递速率对CO2响应的光合参数以及其对应的观测数据Table 3 Measured data and fitted results of CO2-response curves of electron transport rate in wheat at 21%O2 已有的研究结果表明,2%和21% O2浓度下净光合速率之差可用来估算C3植物叶片的光呼吸速率[12, 15, 25-26]。所以,本研究也以此方法用于探讨小麦叶片的光呼吸速率对CO2浓度的响应问题,并在此基础上分析小麦叶片分配到光呼吸的光合电子流的变化规律。由图1可知,由式(7)拟合小麦光呼吸速率对CO2浓度的响应曲线与观测点高度符合,r2为0.934。由此可见,式(7)拟合的结果以及在此基础上探讨的电子流分配等问题上具有可信性。 光合电子流与NADPH以及光合电子流在传递过程产生的跨膜质子梯度驱动ATP合酶形成ATP之间存在密切的关系[27-28]。而光合电子流分配又是植物光合控制的一个重要环节[29]。因此,深入研究不同植物种类及其在不同环境条件下光合电子流传递分配规律将有助于阐明“光合控制”的特点、模式和机理[30]。在C3植物中,光呼吸是非环式光合电子流分配的主要途径[31]。特别在CO2限制的情况下,光呼吸能保持相当的线性电子传递和光能利用,避免由于吸收过量的光能而对植物造成的伤害[32]。因此,对光呼吸所分配电子流的定量研究对探讨电子流的分配显得尤为重要[33]。目前对光呼吸途径的电子流分配主要采用Epron等[18-20]的计算方法(即式(4))。然而,康华靖等[33]的研究表明,该传统方法存在缺陷,难以准确估算光呼吸的电子流分配。我们的研究进一步证明了传统方法计算的分配到光呼吸途径的光合电子流对CO2浓度的响应变化偏离了实际测量曲线。本研究中,由式(4)估算的用于光呼吸途径的电子传递速率随CO2浓度的增加而快速增加,当CO2浓度为164.82 μmol·mol-1时达到最大值(86.93 μmol·m-2·s-1),随后则快速下降;而由图3b和表3的数据可知,在21% O2浓度下,小麦分配到光呼吸途径的最大电子传递速率为84.16 μmol·m-2·s-1(最大光呼吸速率为10.52 μmol·m-2·s-1),对应的饱和CO2浓度约为589.42 μmol·mol-1。由此可见,由公式(4)得到与最大电子传递速率的饱和CO2浓度(约164.82 μmol·mol-1)远小于用公式(8)得到的饱和CO2浓度(约589.42 μmol·mol-1)。由于后者是通过气体交换测量得到光呼吸值,再利用公式(8)计算获得,因此其值更为可信。另外,在21% O2浓度下且CO2浓度等于0 μmol·mol-1时,小麦的光呼吸速率约为4.27 μmol·m-2·s-1(表1)。假如光呼吸产生一个CO2分子需要消耗8个电子[19, 20, 22],则小麦所需光合电子流为34.16 μmol·m-2·s-1。该值远低于利用模型II拟合由式(4)计算而得到的结果(Ca为0 μmol·mol-1时分配到光呼吸电子传递速率的拟合值为58.77 μmol·m-2·s-1),且该拟合值(58.77 μmol·m-2·s-1)也远高于观测值(35.24 μmol·m-2·s-1)。此外,公式(4)是在假定光合电子流只分配到光呼吸和碳同化两条路径中,忽略了分配到其他路径的光合电子流。由此可见,用于计算参与光呼吸途径的光合电子流的公式,即式(4)不能准确地描述Jo对CO2浓度的响应趋势。 由表2可以看出,模型II拟合小麦在21%和2% O2条件下光合电子传递速率对CO2浓度的响应曲线(J-Ca曲线),得到的最大电子传递速率及其对应的饱和CO2浓度与观测值均较为接近。同样,利用模型II拟合由式(8)计算而得到的Jo-Ca曲线,得到的分配到光呼吸的最大电子传递速率值及其对应的饱和CO2浓度也均与观测值较为接近(表3)。此外,由表1和表2的数据可知,21%和2% O2下小麦最大净光合速率对应的饱和CO2浓度为1 368.27和1 233.46 μmol·mol-1;而最大电子传递速率对应的饱和CO2浓度为1 115.66和1 202.27 μmol·mol-1。该值显著高于小麦最大光呼吸速率及其最大电子传递速率所对应的饱和浓度(589.42 μmol·mol-1)(表1和表3)。由此可知,小麦分配到光呼吸的光合电子流或光呼吸达到最大值时,光合作用并没有达到最大值。分析认为,一方面与核酮糖-1,5-二磷酸羧化酶/加氧酶(Rubisco)对CO2的亲和力要远高于O2有关,另一方面高CO2浓度抑制光呼吸,有利于Rubisco朝羧化的方向进行。 另外,由图1c可看出,在较低CO2浓度下,小麦的光呼吸速率随着CO2浓度的降低而减小。这与光呼吸速率随CO2浓度增加而降低的生理生化过程不符[34]。同样,图3b也显示,光呼吸分配的电子流也随着CO2浓度的增加而先升高后降低。管雪强等[35]认为,光呼吸CO2释放减少或消失也许并不代表Rubisco的加氧活性降低或消失的程度,而是其释放的CO2被光合重新利用。从叶片细胞的生理结构来看,CO2释放点靠近叶绿体内则,且没有边界层、气孔及细胞壁的限制,因此光呼吸和暗呼吸产生的CO2极易被光合作用重新利用[36]。利用13C同位素的研究已证实光呼吸和线粒体呼吸的CO2释放均可被光合重新利用[37]。康华靖等[12]认为,对小麦光呼吸速率在低CO2浓度下随着CO2浓度的降低而减小的唯一合理解释则是光呼吸CO2被光合作用重新利用。因此,准确估算光呼吸CO2被光合作用重新利用率,对深入探讨总光呼吸速率及其电子流分配,以及完善本文所构建的模型具有重要意义。 总之,本研究构建的模型II可较好拟合小麦光呼吸速率和分配到光呼吸的光合电子传递速率对CO2浓度的响应曲线,并可给出小麦的最大光呼吸速率和分配到用于光呼吸途径的最大光合电子传递速率,以及所对应的饱和CO2浓度。通过对比这些实验测量结果和拟合结果可知,用于计算参与光呼吸途径的光合电子流的公式 并不能准确地描述Jo对CO2浓度的响应趋势,而本研究构建的新模型可准确地定量探讨光呼吸及其光合电子流分配等问题。
2.3 分配到光呼吸的电子传递速率对CO2的响应


3 讨 论
