苯丙氨酸羟化酶基因突变的生物信息学分析*
2019-04-26来怡君章印红朱宝生唐新华
来怡君,陈 红,章印红,张 杰,朱宝生,唐新华
(1.昆明理工大学医学院,昆明 650500;2.昆明理工大学附属医院/云南省 第一人民医院遗传诊断中心,昆明 650032)
苯丙酮尿症(phenylketonuria,PKU)是一种常见的氨基酸代谢障碍性疾病,属常染色体隐性遗传病,临床表现为智力低下、自闭症、运动障碍等[1]。该病主要因肝脏中的苯丙氨酸羟化酶(phenylalanine hydroxylase,PAH)缺乏或者活性不足导致苯丙氨酸不能正常代谢为酪氨酸而产生[2]。早在1983年PAH基因就被成功分离和克隆,并被公认为是PKU的致病基因[3]。近几年,随着基因组和外显子组测序的发展,很多新发突变,特别是一些症状相对较轻的突变也逐渐被检测出来[4]。截至目前,已经在患有PKU的患者中鉴定了约1 000种不同的PAH基因突变体[5]。并且PAH基因具有突变异质性,不同种族与地区的突变位点及分布有较大差异[6]。本研究采用云南省汉族PKU患者中发现的6例PAH基因新发突变为材料,从生物信息学角度分析PAH基因的错义突变与蛋白结构,以及进一步可能导致的功能改变的关系,并利用生物信息学方法尝试探讨新发剪接突变的发生机制,为以后关于PKU的研究提供有价值的信息。
1 材料与方法
1.1材料 (1)6例数据库中未收录的PAH基因新突变见表1。(2)使用的网络资源,包括美国国立生物技术信息中心(NCBI,网址:https://www.ncbi.nlm.nih.gov/)、Swiss-Model软件(https://www.swissmodel.expasy.org/)、CRYP-SKIP(http://cryp-skip.img.cas.cz/)、BDGP软件(http://www.fruitfly.org/seq_tools/splice.html)、HSF(http://www.umd.be/HSF/)等。
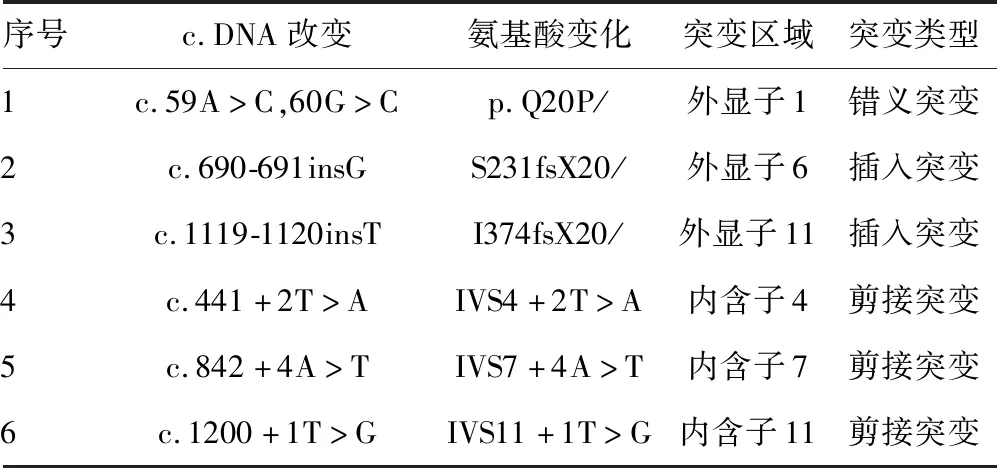
表1 云南地区PKU患者PAH基因新突变[7]
1.2方法
1.2.1获取人类PAH基因全序列及其分析 从NCBI的GenBank中基因数据库获取人类PAH基因组定位和其编码的蛋白质序列信息。
1.2.2PAH新发错义突变及插入突变位点所在外显子的同源建模及功能分析 利用Swiss-model在线工具对1例错义突变和2例插入突变所在外显子区域进行外显子同源建模,分析基因突变对其所在外显子的蛋白结构和功能的影响。
1.2.3PAH新发剪接突变的生物学效应分析 采用3种基于不同网络算法的生物信息学工具分析新发现的剪接变异。其中,在CRYP-SKIP界面上直接将变异序列按其要求的格式输入即可获得预测结果,包括因剪接变异导致的隐蔽剪接位点激活(PCR-E)和外显子跳跃(1-PCR-E)的可能性大小两个结果[8]。柏克莱果蝇基因组项目(Berkeley Drosophila Genome Project,BDGP)和人类剪接查找服务器(the Human Splicing Finder,HSF)可预测可能存在的隐蔽剪接位点及基因变异对剪接位点的改变,使用时分别将野生型基因序列和变异型基因序列进行在线分析,即可获得变异前后目标基因序列所有可能的剪接位点及分值。
2 结 果
2.1PAH结构 在GenBank中基因数据库查找PAH基因可知:PAH基因全长79 278 bp,位于人类12号染色体长臂上(12q23.2),包含13个外显子,12个内含子,CDS编码452个氨基酸,在66~1 421 bp见图1。

图1 人类PAH基因结构
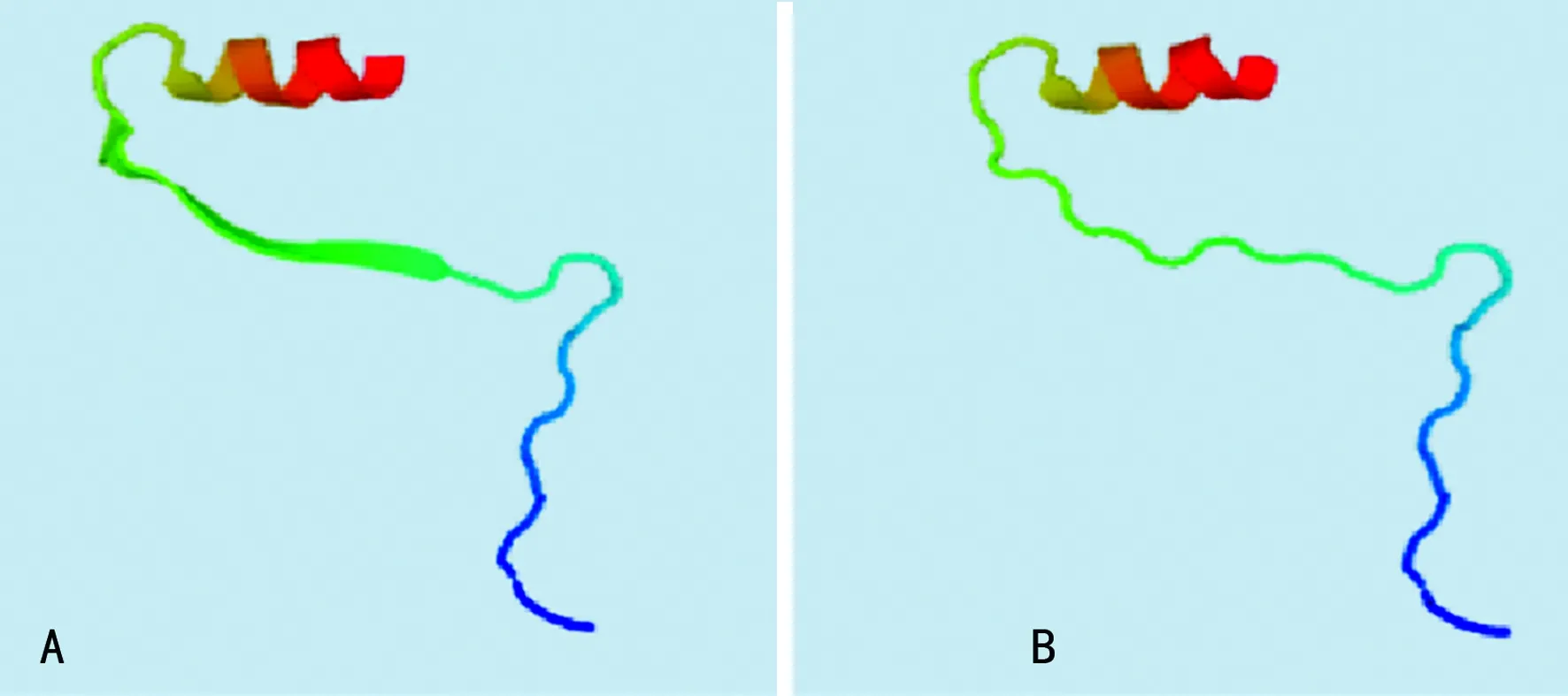
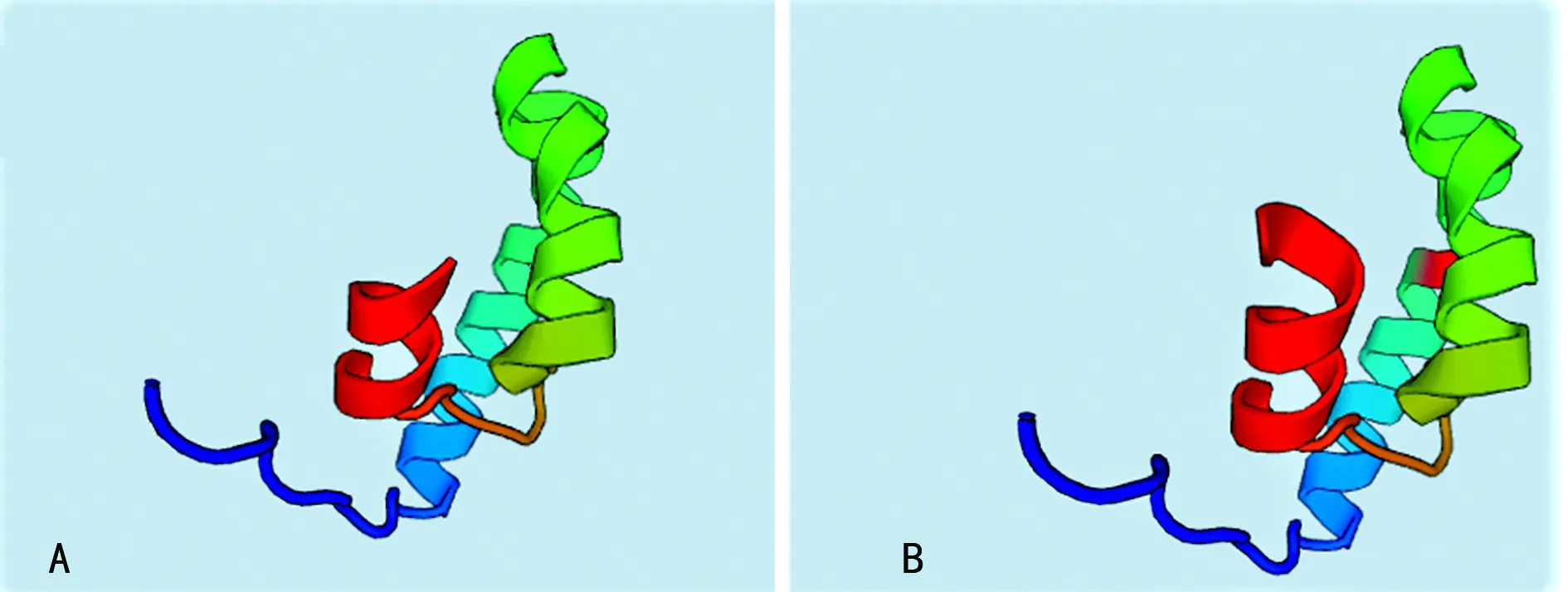
2.2PAH突变位点外显子的同源建模及功能分析 由于本研究中的3例突变分别发生在外显子1、外显子6和外显子11上,而外显子1因其序列太短未找到同源序列,因此将外显子1,2结合起来对外显子1处发生的点突变(c.59A>C,60G>C)进行同源建模,并对突变后的外显子6和外显子11分别进行同源建模,见图2。其中,图2A是正常的外显子1,2结构,图2B是点突变(c.59A>C,60G>C)突变后的外显子1,2预测结构,可以看出突变导致外显子的一处β片层消失。图3A是正常的外显子6的空间结构,图3B中由于(c.690-691insG),导致移码突变S231fsX51,即该三联密码子下游第51位氨基酸处蛋白合成终止,致使外显子6的空间构象发生明显改变。外显子11上的第1 119位和1 120位插入碱基T(c.1119-1120insG),导致移码突变I374fsX,即374位后第20位氨基酸处蛋白合成终止,由于此处突变可能会影响外显子12和13的合成,因此选择从正常的外显子10开始到外显子13进行同源建模,同时对(c.1119-1120insG)突变后的外显子10之后的部分进行同源建模。图4A是正常的外显子10,11,12,13的结构;图4B是正常的外显子10,11的结构;图4C是(c.1119-1120insG)突变后外显子10,11,12,13的结构,可以看出(c.1119-1120insG)的突变导致外显子11三级结构部分缺失。
2.3PAH新发剪接突变的生物学效应分析 经3种生物信息学软件综合分析上述突变,CRYP-SKIP预测值中,PCR-E(在右边的刻度盘上用红色指针表示)取值介于0和1之间,较高的值表示支持隐蔽剪接位点激活,较低值则表示支持外显子跳跃。突变预测结果显示c.441+2T>A突变后第4外显子5′端产生隐蔽剪接位点(PCR-E)的可能性为0.35,3′端的值接近于0,c.842+4A>T突变后第7外显子5′端产生隐蔽剪接位点(PCR-E)的可能性为0.39,3′端的值接近于0。c.1200+1T>G突变后外显子11中5′端产生隐蔽剪接位点(PCR-E)的可能性为0.66,3′端的值接近于0。
BDGP显示正常PAH基因第4内含子5′端剪接位点(AGgt,g为突变处)的预测值为0.94(预测值为0~1),c.441+2T>A突变后第4内含子5′端剪接位点(AGgt,g为突变处)的预测值则为0.96(预测值为0~1),而IVS4+2T>A和IVS7+4A>T突变后BDGP预测值均消失(预测值为0)。
HSF预测中CV值高于80者具有较强剪接信号,CV值为70~80的剪接位点剪接信号较弱,少部分剪接位点的CV值为65~70[9](预测值为0~100)。预测结果显示c.441+2T>A使第4内含子的5′剪接供体共识值(CV值)由正常的83.22变为56.39。c.842+4A>T使第7内含子的5′剪接供体CV值为由82.11变为73.32,而3′剪接受体CV值由68.35变为39.4。c.1200+1T>G突变使第11内含子的5′剪接供体CV值为由95.95变为69.12,而3′剪接受体CV值并无明显变化。
A:正常的外显子1,2结构;B:外显子1处点突变突变后的预测结构
图2外显子1,2的同源建模结果
A:正常外显子6的空间结构;B:外显子6空间构象发生改变
图3外显子6的同源建模结果
A:正常的外显子10,11,12,13的结构;B:正常的外显子10,11的结构;C:突变后外显子10,11,12,13的结构
图4外显子10-13的同源建模
3 讨 论
一般认为,PKU是由于PAH基因突变引起PAH蛋白功能损坏或丧失而导致的,PAH基因中细微的改变都会引起酶蛋白三维结构的改变,并可造成PAH活性降低甚至完全丧失,导致在临床上产生PKU病患。而PAH蛋白的三级结构是一个同源四聚体。此前对于PKU基因型与表型关系的研究多是利用质谱技术和 X射线衍射技术构建PAH的3D晶体结构模型来进行的[10],而最新研究发现了全长哺乳动物(大鼠)PAH的第1个X射线晶体结构[11]。本研究中使用Swiss-model对1例错义突变和2例插入突变所在的第1、第6和第11外显子的同源建模分析中发现(c.59A>C,60G>C)突变导致外显子1,2区域的片层结构消失,另外此突变位于外显子与内含子的交界处,可能会严重影响PAH基因后续的转录、翻译和蛋白折叠等[6]。(c.690-691insG)突变导致外显子6的空间构象发生改变;而外显子11上的(c.1119-1120insG)突变不仅影响了外显子11的空间构象,并且严重影响了后续外显子12和外显子13的构象,这3例突变预测的结果显示突变会直接影响PAH单体的结构完整性。而最近有研究揭露PKU相关的基因突变会对PAH同源四聚体的稳定性产生影响,造成PAH蛋白不同程度的降解,从而导致严重的PKU表型[12]。例如PAH基因R408W突变后导致翻译的蛋白质极易降解,最终在患者体内或体外测不出PAH,从而导致严重的PKU表型,而R413P突变则导致PAH四聚体结构的高度不稳定,影响正常的单体状态和蛋白酶的功能,临床表现为严重代谢障碍型PKU。所以作者大胆推测,这3例突变甚至有可能导致突变体的稳定性受到影响,致使PAH蛋白降解,从而影响其功能。
目前,在PKU相关的PAH基因缺陷中,约13%的突变影响保守的3′和5′剪接位点,而大多数报道的剪接突变都是通过影响5′端剪接供体位点从而引起疾病的[13]。本研究中对于3种剪接突变的多种预测结果显示,第7内含子处的c.842+4A>T突变可能导致外显子的3′端发生了跳跃,第4内含子的c.441+2T>A突变和第11内含子处c.1200+1T>G的突变都有可能是由于外显子的5′端影响剪接,其中第4内含子的c.441+2T>A突变可能原因是其5′端发生了跳跃。在最近一项PAH基因外显子11处两个致病剪接突变(c.1199+17G>A和c.1199+20G>C)的研究中发现,与5′剪接位点相结合的小核糖核蛋白颗粒U1 snRNA在与外显子11的剪接过程中起着重要作用,而突变则会破坏U1 snRNA和内含子5′剪接位点的结合,从而影响剪接。此外还证明了通过修饰U1 snRNA可以实现对剪接突变的校正[14]。本研究中第11内含子处c.1200+1T>G突变的预测结果说明,突变可能降低了11内含子5′端隐蔽剪接位点的活性,但c.1200+1T>G突变是否同样破坏U1 snRNA和内含子5′剪接位点的结合还不得而知,需要进一步的实验验证。
尽管本研究采用生物信息学方法对PAH基因突变进行了分析,但基因突变对功能的影响是一个复杂的过程,且生物信息学方法还有待提升,具体的基因突变与功能的关系及突变后的机制还要以实验结果为准,这需要后续进一步的系统研究。
