圈养环境下瓶鼻海豚、白鲸、伪虎鲸click脉冲信号特征分析
2019-04-11李赫高大治林建恒迟静
李赫,高大治,林建恒,迟静
圈养环境下瓶鼻海豚、白鲸、伪虎鲸click脉冲信号特征分析
李赫1,4,5,高大治2,林建恒3,迟静2
(1. 中国科学院水声环境特性重点实验室,北京 100190;2. 中国海洋大学海洋技术系,山东青岛 266100; 3. 中国科学院声学研究所北海研究站,山东青岛 266109;4. 中国科学院大学,北京 100049; 5. 中国科学院声学研究所,北京 100190)
通过实地采样采集到的瓶鼻海豚、白鲸、伪虎鲸的click脉冲信号数据,对所采集信号分别从时域、频域、时频特性以及距离分辨力等方面分别进行了分析对比。结果表明,三种海洋哺乳类动物均通过不断发射click脉冲串实现对目标的探测,在这过程中可以调节脉冲间隔或信号波形,其中伪虎鲸一部分click脉冲信号的频谱具有双峰的特殊性质。通过对这三种海洋哺乳动物click脉冲信号模糊函数的分析,发现瓶鼻海豚与白鲸的距离分辨力可以达到毫米量级,伪虎鲸相对较差,但也可以达到厘米量级。而随着频谱双峰信号的出现,伪虎鲸click脉冲信号的距离分辨力有明显提高,也达到了毫米量级。
海洋哺乳动物;click脉冲信号;模糊函数;
0 引言
生物噪声是海洋环境噪声重要的组成部分[1]。在诸多生物噪声源中,海洋哺乳动物长期以来一直是学者们重点研究的对象。海洋哺乳动物是海洋中高等的生物群体,在水下世界中,海豚、鲸类等海洋哺乳动物的视力极差,他们利用回声定位实现觅食、躲避、通信等生理活动。对海洋哺乳动物回声定位能力的研究对于声呐信号设计、仿生学等学科领域有着极大的意义。
瓶鼻海豚是一种常见的海豚,B. K. Branstetter等[2]曾对瓶鼻海豚的三种不同叫声信号分别从时域、频域进行了较为详细的分析,并介绍了其指向性,使人们对瓶鼻海豚的发声信号有了一定的了解与认识。除瓶鼻海豚外,国内外的学者对其他具有发射信号能力的海洋哺乳动物也有一定的研究,这些哺乳动物中有体型较大的须鲸[3]、抹香鲸[4],凶猛的虎鲸[5-6],也有智力极高的白鲸[7-9]和伪虎鲸等[10-12]。
海豚、鲸类等大部分常见海洋哺乳动物的发音大体可分为三类:click脉冲信号、whistle扫频信号、burst爆破音[2]。除海豚外,能够发出click脉冲信号的还有白鲸、伪虎鲸、抹香鲸等。L. E. Freitag等[13]在1992年就曾对瓶鼻海豚的click脉冲信号和whistle扫频信号的回声定位能力进行了研究。在国内,牛富强等[14-15]、汪启铭等[16]曾对中华白海豚、瓶鼻海豚的click脉冲信号进行分析,介绍了其时频域等多方面的特性;除此之外,刘维等[17]、薛山花等[18]对海豚声信号的实验研究也都为本文提供了良好的参考依据。早在1984年,W. W. L. Au等[19]对采集白鲸的click脉冲信号进行特征分析,发现其具有高频宽带的特点,并有一定的调节能力。1995年,W. W. L. Au等[20]将伪虎鲸的click脉冲信号分为了4种类型,并已经发现其具有双频谱峰的特点。而近几年,文献[21]中介绍,通过考察夏威夷海域伪虎鲸click脉冲信号以及whistle扫频信号的声源级,分析了其探测距离。但对于几种海洋哺乳动物click脉冲信号的特性及距离分辨能力尚没有学者进行详细比较,本文分别从时域、频域、时频分析以及模糊度函数等方面,对在青岛海昌极地海洋世界采集到的瓶鼻海豚、白鲸以及伪虎鲸的click脉冲信号的特性进行了对比,分析了其距离分辨力差异。
1 信号采集实验
为了采集几种海洋哺乳动物的声呐信号,记录设备采用带有前置放大器的水听器、National Instrument 公司PXI 1042Q信号采集器,采样频率为200 kHz。信号录取对象为青岛海昌极地海洋世界人工饲养池内的瓶鼻海豚、白鲸以及伪虎鲸,瓶鼻海豚的饲养环境如图1所示,4头瓶鼻海豚饲养在约50 m2的饲养池中,水深约5 m,为防止回波信号带来的干扰,水听器放在水面下2 m,并固定在表演池和饲养池间的闸门上。白鲸和伪虎鲸各两头分别饲养在各自的饲养池当中,饲养池大小约为20 m2。当几种海洋哺乳动物在水下发出声信号时,水听器作为接收端可以接收到信噪比较高的声信号,通过实时数据显示,记录下质量较高的时间节点,以便于实验后的数据查找以及处理。

图1 瓶鼻海豚、白鲸、伪虎鲸的饲养环境
2 三种典型海洋哺乳动物click脉冲信号分析
海洋哺乳动物的click脉冲信号具有持续时间短、频带宽的特点,主要用来实现定位、探测等生理活动。因在实验条件下,几种动物一直处于游动状态,无法保持其与水听器的距离不变,因此本文研究的特征只涉及时间以及频率的特征,而不涉及声强的实际测量值。忽略声强分析对于本文分析研究的影响可以不予考虑。
2.1 瓶鼻海豚click脉冲信号分析
海豚以脉冲串的形式发射click脉冲信号,通过对回波的处理从而实现各种生理活动。每个脉冲串所包含的脉冲个数并不相等,少的仅仅几个,多的几百个。图2是一串典型的瓶鼻海豚的click脉冲串,其中包含几十个单脉冲信号,每两个脉冲信号的间隔大多为几十毫秒,信号间隔随着时间逐步减小。图3给出了脉冲间隔的变化情况,从最初的60 ms最终趋于20 ms左右。图4为脉冲串中一个单脉冲信号的时域波形,每个单脉冲的长度基本恒定,在100 μs以下。
在软件中,利用一维离散快速傅里叶变换可对信号进行谱分析以及时频特性的分析,根据脉冲间隔和单脉冲的持续时间,在保证图像分辨率的前提下,将时间窗长度均取为5 ms。图5是图2所示脉冲串的时频特性图,瓶鼻海豚的click脉冲信号属于宽频信号,频率范围较宽,从10 kHz左右到100 kHz以上均有成分,其中能量在30~60 kHz范围内相对较高,而每个单脉冲的频率范围并不完全一致。
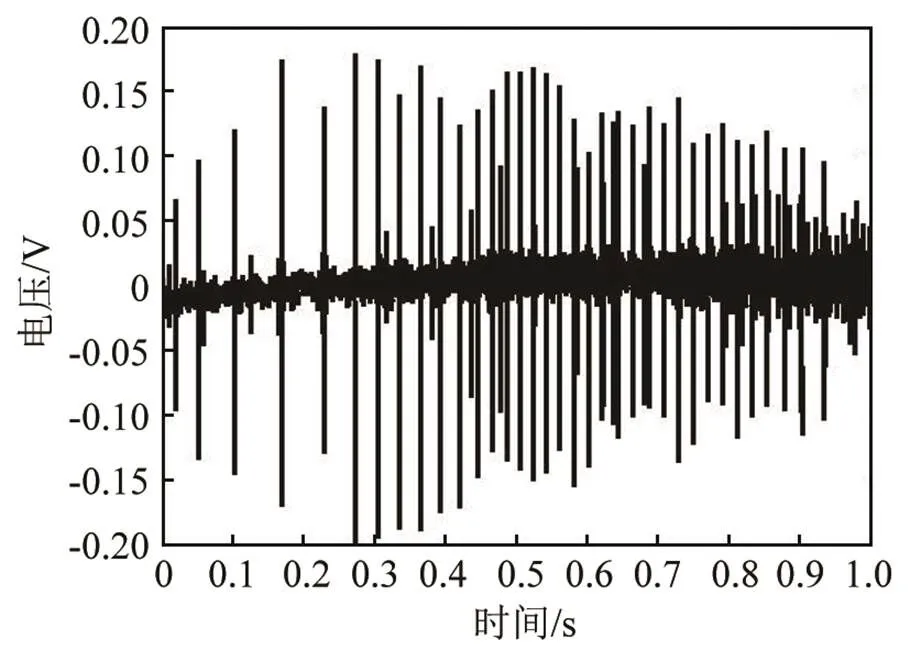
图2 瓶鼻海豚click脉冲串

图3 瓶鼻海豚click脉冲时间间隔变化曲线

图4 瓶鼻海豚单脉冲信号时域波形
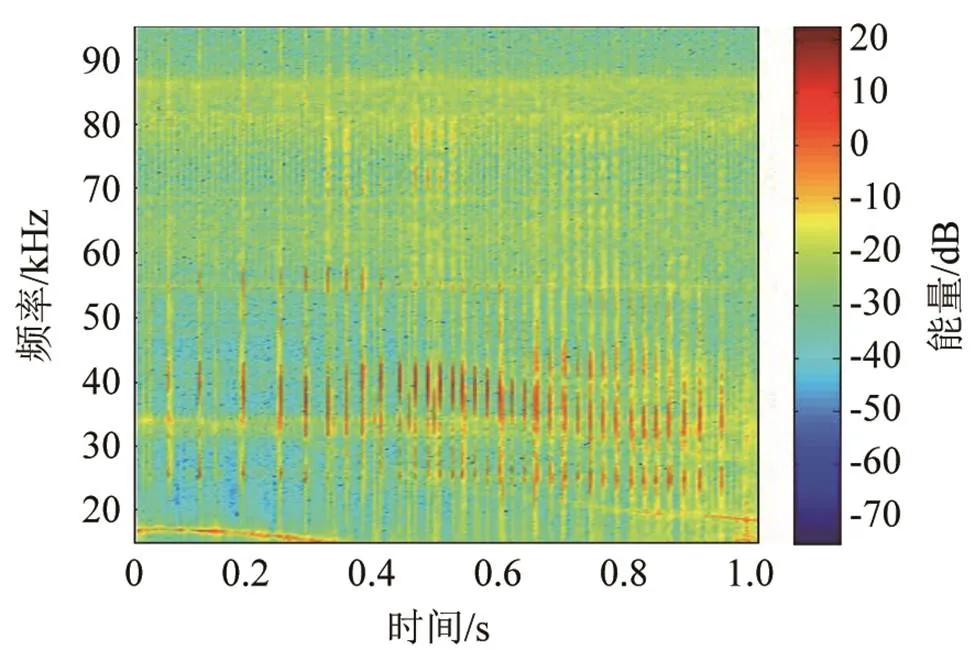
图5 图2中瓶鼻海豚click脉冲串时频特性图



图6 瓶鼻海豚单脉冲信号三维模糊函数图

图7 瓶鼻海豚单脉冲信号模糊度图

图8 瓶鼻海豚脉冲串前20个脉冲时延测量精度变化图
2.2 白鲸click脉冲信号分析
与海豚一样,白鲸的click脉冲信号也用来实现探测、定位等生理功能。在时域上,白鲸的click脉冲串持续时间较长,且每两个脉冲之间间隔较小。图9是截取了一串脉冲信号其中的一部分的信号时域图,脉冲间隔非常稳定,几乎不会随着时间有明显变化,其间隔均在3 ms左右。单个脉冲信号的长度与瓶鼻海豚相近,在100 μs左右,也没有发现其随时间变化,白鲸单脉冲信号的时域波形如图10所示。在频域上,白鲸的click脉冲信号频谱能量在15 kHz左右到100 kHz以上均有分布,其中30~50 kHz范围内能量分布较高,其余范围内较低,如图11所示。相比瓶鼻海豚而言,最低频率要稍高一些,在人耳可听域内能量分布极少,因此在录取的音频文件中很难听到白鲸的click脉冲的声音。

图9 白鲸click脉冲串
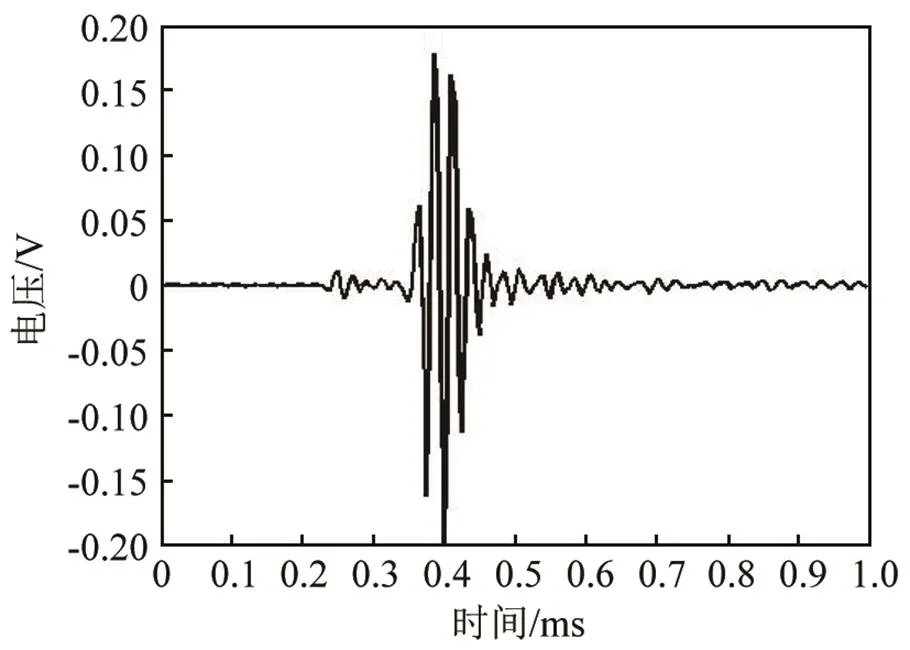
图10 白鲸单脉冲信号时域波形
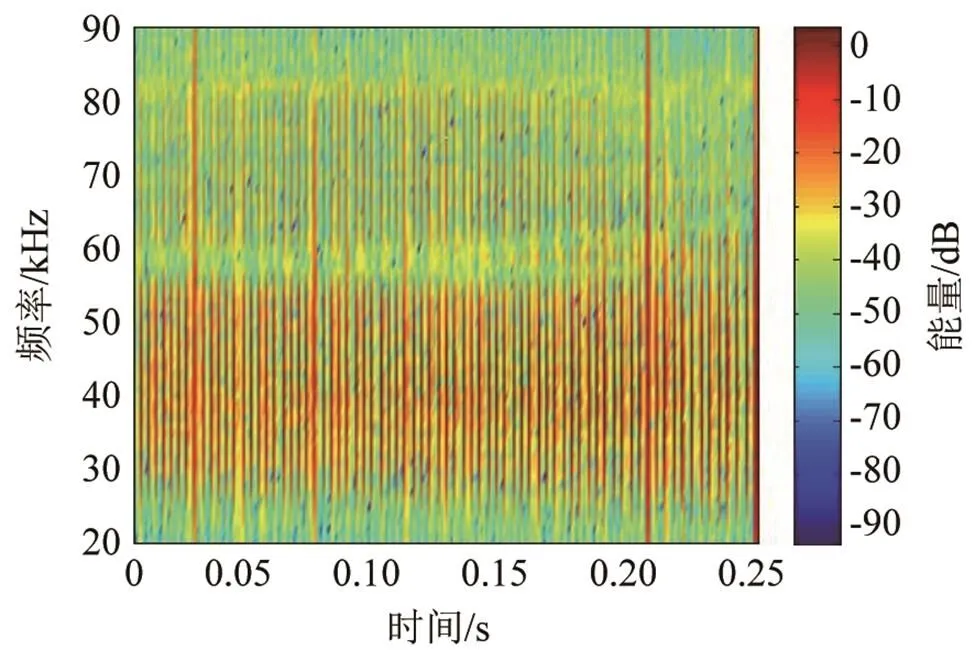
图11 白鲸click脉冲信号时频特性图
白鲸的click脉冲信号对于速度的分辨能力很差,但对距离的分辨能力较强,这点与瓶鼻海豚相同,图12、13是一个较为理想的单脉冲信号的三维模糊函数图以及模糊度图。该脉冲信号的时延测量精度为1.5 μs,在声速为1 500 m·s-1的水中,该脉冲信号对距离的分辨能力为2.25 mm,这与瓶鼻海豚相近;而整个脉冲串的距离分辨力随时间变化也并不明显,每个单脉冲的时间测量精度均在2 μs上下浮动(见图14);从实际信号采样情况上分析,在录取白鲸声呐信号时,饲养池中的白鲸一直在水听器附近徘徊,其到距离基本没有太明显变化。

图12 白鲸单脉冲信号三维模糊函数图
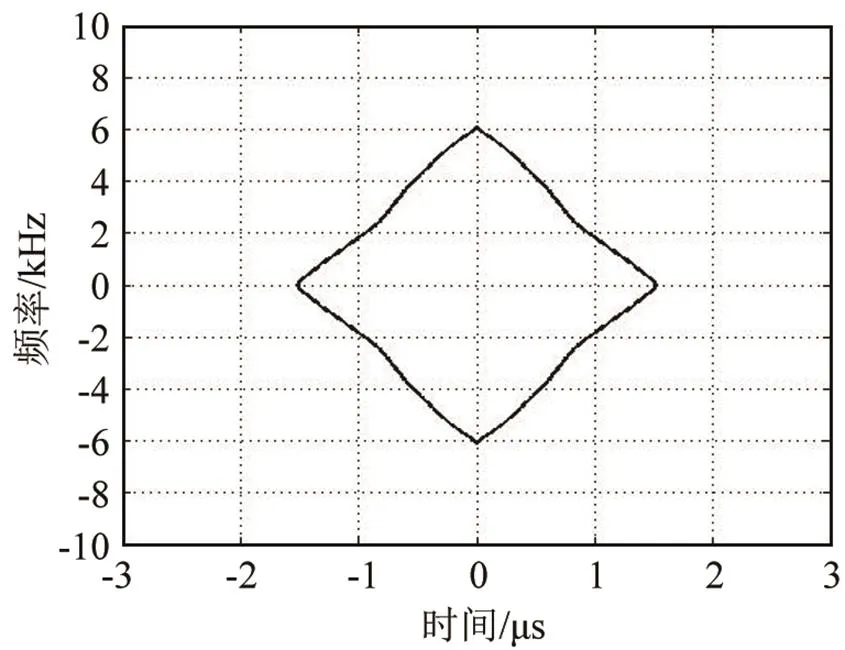
图13 白鲸单脉冲信号模糊度图
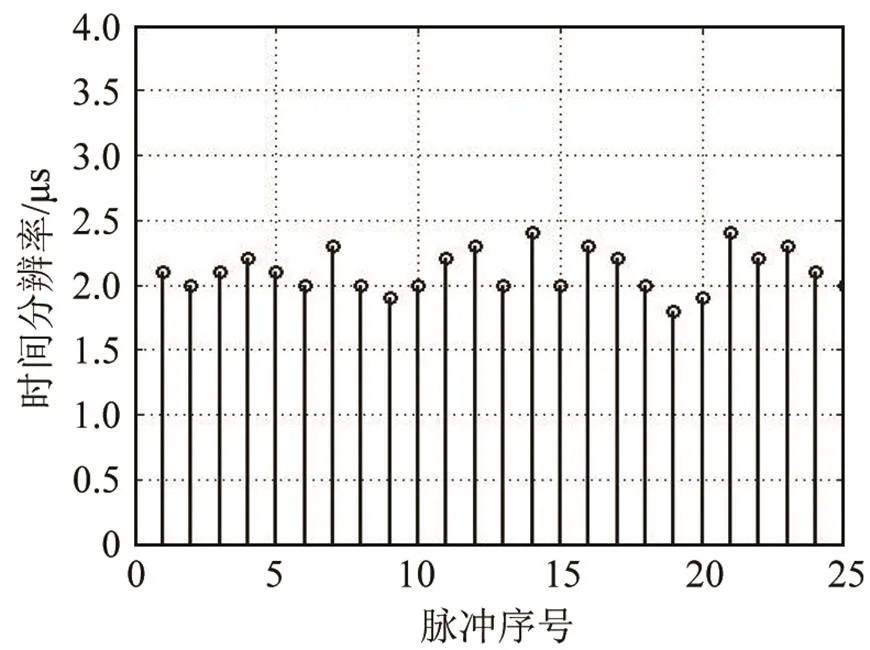
图14 白鲸脉冲串25个脉冲时延测量精度变化图
2.3 伪虎鲸click脉冲分析
图15是一串完整的伪虎鲸click脉冲串及其时频图,其中总共包含15个单脉冲信号,其数量要远小于瓶鼻海豚与白鲸的脉冲串数量。每个单脉冲信号的长度要长于前两者,量级为毫秒,如图16所示。而在已有数据中,其脉冲间隔随时间逐渐扩大,图17给出了其变化趋势,从最初的43 ms逐渐变宽,到最后的150 ms,这显然与瓶鼻海豚的变化趋势相反,且脉冲宽度要远宽于白鲸。
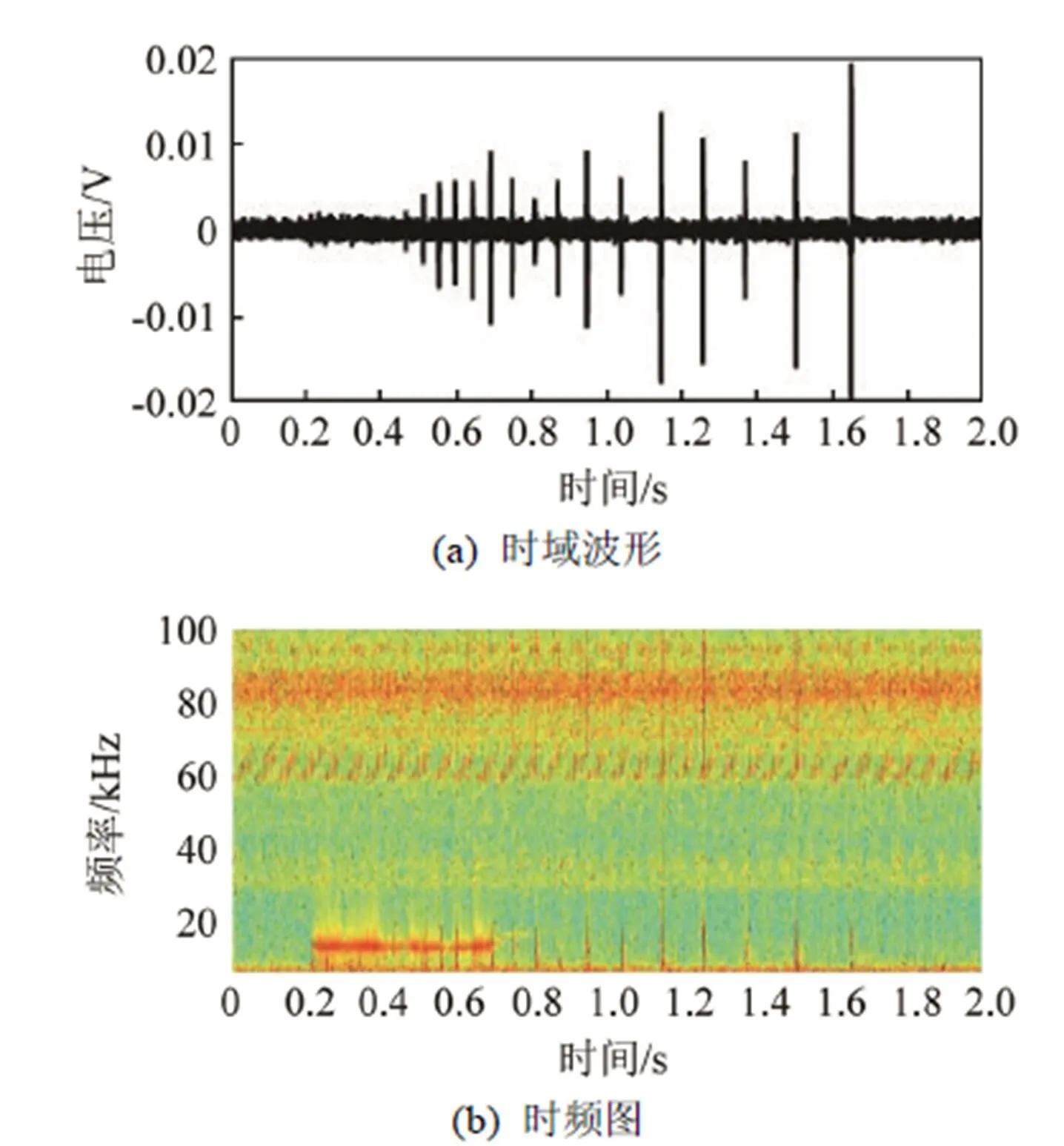
图15 伪虎鲸click脉冲串的时域波形和时频图

图16 伪虎鲸单脉冲信号时域波形

图17 伪虎鲸click脉冲间隔变化曲线
在频域上,伪虎鲸的click脉冲信号显示出了其与前两者截然不同的特点,为了明显的对比,将其放在一起进行比较。对于脉冲串中前一部分脉冲信号,频率范围与前两者相比较低,在10 kHz以下,而后一部分信号的能量主要分布在两个范围内,一部分在10 kHz以下,另一部分在60~100 kHz附近,而中间频段能量非常低,在时频图中并不明显。为了证明这种双频率峰真实存在而不是数据处理过程的错误,图18和19分别给出了该脉冲串中第三个和最后一个单脉冲信号的频谱图(其余图像在此不一一列出),虽然时频图中80 kHz处存在一个干扰,但在70 kHz与90 kHz附近峰值明显,说明这种一个脉冲内含有双峰现象是切实存在的。
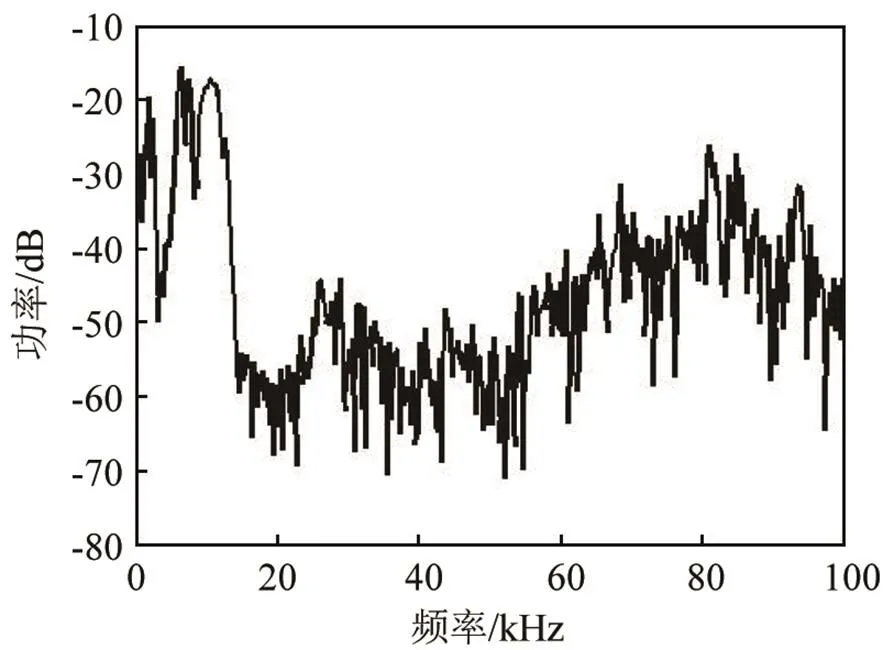
图18 伪虎鲸第3个单脉冲信号频谱图

图19 伪虎鲸第15个单脉冲信号频谱图
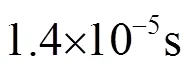

图20 伪虎鲸单脉冲信号三维模糊函数图

图21 伪虎鲸单脉冲信号模糊度图

图22 伪虎鲸click脉冲串时延测量精度变化图
2.4 瓶鼻海豚、白鲸、伪虎鲸click脉冲信号对比
为更准确地比较三种海洋哺乳动物在频域上的特点,在此对100个瓶鼻海豚、100个白鲸、30个伪虎鲸click脉冲信号进行频谱分析,并汇总统计。如图23~25所示。结果表明,本次实验所采集的瓶鼻海豚的click脉冲信号峰值频率出现在30 kHz左右,白鲸的click脉冲信号峰值频率出现在30~50 kHz之间,而伪虎鲸的click脉冲信号峰值频率则在8 kHz左右和60 kHz以上。
表1为三种海洋哺乳动物click脉冲信号对比。
从表1中看出,瓶鼻海豚与白鲸的click脉冲信号的单脉冲长度、频率范围较为相近,且都具有较高的距离分辨力,可以分辨相距几毫米的目标;而伪虎鲸的单脉冲长度以及脉冲间隔都要大于前两者,频谱单峰脉冲信号对距离的分辨能力也与海豚和白鲸相差一个数量级,而在增加了高频峰之后,距离分辨力显著提高,与瓶鼻海豚和白鲸相近。

图23 瓶鼻海豚click脉冲信号频谱统计图

图24 白鲸click脉冲信号频谱统计图

图25 伪虎鲸click脉冲信号频谱统计图
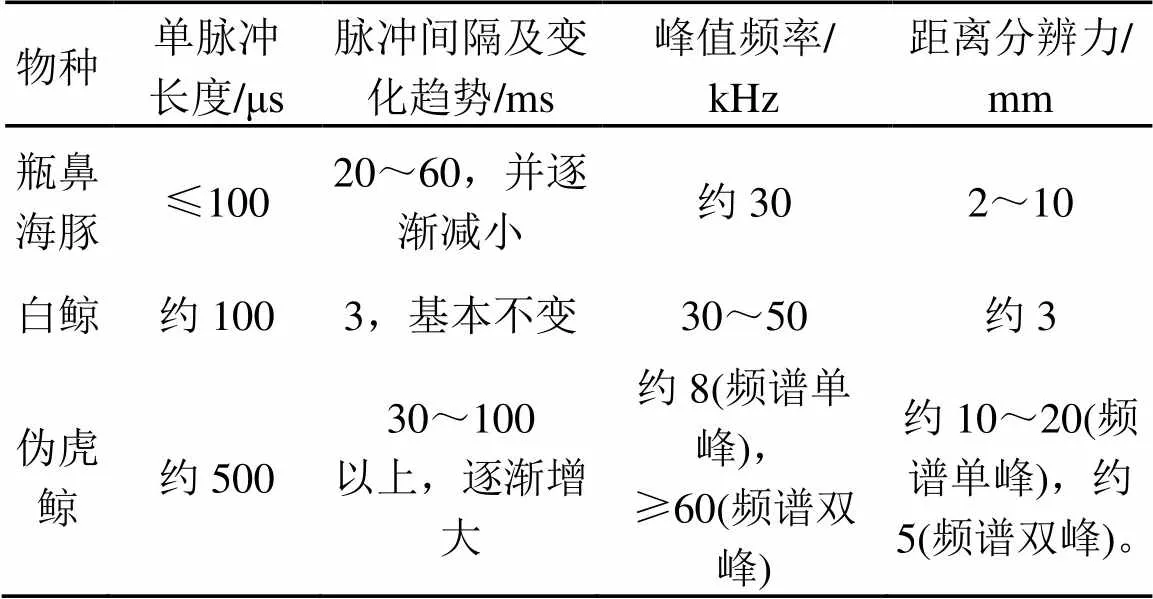
表1 三种海洋哺乳动物click脉冲信号对比
3 结论
瓶鼻海豚、白鲸、伪虎鲸三种海洋哺乳动物均通过click脉冲信号来实现探测、回声定位等生理活动。本文通过实地采样,对采集的三种典型海洋哺乳动物的click脉冲信号分别从时域、频域、时频特性以及模糊函数等角度加以处理和分析。结果表明:三种海洋哺乳动物均通过不断地发射宽频的窄脉冲信号串来实现对目标的探测等生理活动,在这个过程中可以调节所发射信号的形式以及特性。伪虎鲸的单脉冲信号长度要长于瓶鼻海豚和白鲸,前者以毫秒为量级,与后者相差一个数量级;脉冲间隔白鲸最为密集,发射频率较高;瓶鼻海豚和白鲸的能量在10 kHz左右到100 kHz以上均有分布,距离分辨力可以达到毫秒量级;而伪虎鲸的部分信号出现了频谱双峰现象,频谱单峰信号的距离分辨力以厘米为量级,随着频谱双峰的出现,其距离分辨能力有显著的提高,达到了毫米量级。
海洋哺乳类动物发声信号的形式多种多样,每种发声形式都有其特定的功能与规律,本文仅对click脉冲信号进行了简要的分析对比,为日后的海洋哺乳类动物研究工作提供了一定的基础。而海洋哺乳动物的声呐系统极其复杂,它们是否会在活动中随着环境的不断变化自适应地调节信号的特性,本文尚不能给出确定的结论;而伪虎鲸click脉冲信号的频谱双峰的形成机理在此也不能给出详尽的解释,以上都是今后工作的重点。
[1] WENZ G M. Acoustic ambient noise in the ocean: spectra and sources[J]. J. Acoust. Soc. Am., 1962, 3(12): 1936-1956.
[2] BRANSTETTER B K, MOORE P W, FINNERAN J J, et al. Direction properties of bottlenose dolphin (tursiops truncatus) clicks, burst-pulse, and whistle sounds[J]. J. Acoust. Soc. Am., 2012, 131(2): 1613-1621.
[3] NIEUKIRK S L, FREGOSI S, MLLINGER D K, et al. A complex baleen whale call recorded in the mariana trench marine national monument[J]. J. Acoust. Soc. Am., 2016, 140(3): EL274- EL279.
[4] MOHL B, WAHLBERG M, MADSEN P T, et al. The Monopulsed Nature of Sperm Whale Clicks[J]. J. Acoust. Soc. Am., 2003, 114(2): 1143-1154.
[5] SAMARRA FILIPA I P, DEECKE V B, MILLER P J O. Low-frequency signals produced by northeast atlantic killer whales[J]. J. Acoust. Soc. Am., 2016, 139(3): 1194-1157.
[6] ANDRIOLO A, REIS S S, AMORIM T O S, et al. Killer whale (orcinus orca) whistles from the western south atlantic ocean include high frequency signals[J]. J. Acoust. Soc. Am., 2015, 138(3): 1696-1701.
[7] AU W W L, CARDER D A, PENNER R H, et al. Demonstration of adaptation in beluga whale echolocation signal[J]. J. Acoust. Soc. Am., 1985, 77(2): 726-730.
[8] SUPIN A Y, POPOV V V. Evoked-potential recovery during double click stimulation in a beluga whale: implications for biosonar gain control[J]. J. Acoust. Soc. Am., 2015, 137(5): 2512-2521.
[9] BOT O LE, SIMARD Y, ROY N, et al. Whistle source levels of free-ranging beluga whales in saguenay-st. lawrence marine park[J]. J. Acoust. Soc. Am., 2016, 140(1): EL89-EL93.
[10] BRILL R L, PAWLOSKIJ L, HELWEG D A, et al. Target detection, shape discrimination, and signal characteristics of an echolocating false killer whale (pseudorca crassidens)[J]. J. Acoust. Soc. Am., 1992, 92(3): 1324-1330.
[11] SUPIN A Y, NACHTIGALL P E, BREESE M. Interaction of emitted sonar pulses and simulagted echoes in a false killer whale: an evoked-potential study[J]. J. Acoust. Soc. Am., 2011, 130(3): 1711-1720.
[12] THODE A, WILD L, STRALEY J, et al. using line acceleration to measure false killer whale (pseudorca crassidens) click and whistle source levels during pelagic longline depredation[J]. J. Acoust. Soc. Am., 2016, 140(5): 3941–3951.
[13] FREITAG L E, TYACK P L. Passive acoustic localization of the atlantic bottlenose dolphin using whistles and echolocation clicks[J]. J. Acoust. Soc. Am., 1993, 93(4): 2197-2205.
[14] 牛富强, 杨燕明, 许肖梅, 等. 厦门海域中华白海豚定位click信号特性[J]. 厦门大学学报, 2012, 51(5): 951-954.
NIU Fuqiang, YANG Yanming, XU Xiaomei, et al. Study on the characteristics of echolocation clicks from indo-pacific Humpback dolphins() in Xiamen[J]. Journal of Xiamen University, 2012, 51(5): 951-954.
[15] 牛富强, 杨艳明, 文洪涛, 等. 瓶鼻海豚的click声信号特性[J]. 声学技术, 2011, 30(2): 148-152.
NIU Fuqiang, YANG Yanming, WEN Hongtao, et al. Vocalization and signal characteristic of bottlenose dolphin[J]. Technical Acoustics, 2011, 30(2): 148-152.
[16] 汪启铭, 许肖梅, 苏冠龙, 等. 厦门和雷州湾海域中华白海豚click信号的分析与比较[J]. 厦门大学学报, 2014, 53(1): 104-109.
WANG Qiming, XU Xiaomei, SU Guanlong, et al. The Analysis and Comparison of the clicks ofin Xiamen Sea and Leizhou Bay[J]. Journal of Xiamen University, 2014, 53(1): 104-109.
[17] 刘维, 孙尼, 尹力. 海豚声纳信号的脉冲分解及特征分析[J]. 应用声学, 2009, 28(6): 431-438.
LIU Wei, SUN Ni, YIN Li. Pulse separation and characteristics analysis of dolphin’s sonar signal[J]. Applied Acoustics, 2009, 28(6): 431-438.
[18] 薛山花, 刘维, 陈学亮, 等. 海豚声信号采集与数据分析[J]. 声学技术, 2008, 27(5): 398-399.
XUE Shanhua, LIU Wei, CHEN Xueliang, et al. Record and analysis on sound of the dophins[J]. Technical Acoustics, 2008, 27(5): 398-399.
[19] AU W W L, CARDER D A, PENNER R H, et al. Demonstration of adaptation in beluga whale echolocation signals[J]. J. Acoust. Soc. Am., 1985, 77(2): 726-730.
[20] AU W W L, PAWLOSKI J L, NACHTIGALL P E, et al. Echolocation signals and transmission beam pattern of a false killer whale (pseudorca crassidens)[J]. J. Acoust. Soc. Am., 1995, 98(1): 51-59.
[21] 田坦. 声呐技术[M]. 第2版. 哈尔滨: 哈尔滨工程大学出版社, 2009: 22-30.
TIAN Tan. Sonar technology[M]. 2nd Edition. Harbin: Harbin Engineering University Press, 2010: 22-30.
Analyses of click signal characteristics of captive bottlenose dolphins, beluga whales and false killer whales
LI He1,4,5, GAO Da-zhi2, LIN Jian-heng3, CHI Jing2
(1. Key Laboratory of Underwater Acoustic Environment, Institute of Acoustics, Chinese Academy of Sciences, Beijing 100190, China; 2. Department of Marine Technology, Ocean University of China, Qingdao 266100, Shandong, China; 3. Qingdao Laboratory, Institute of Acoustics, Chinese Academy of Sciences, Qingdao 266109, Shandong, China; 4. University of Chinese Academy of Sciences, Beijing 100049, China; 5.Institute of Acoustics, Chinese Academy of Sciences, Beijing 100190, China)
The click pulse signals of dolphins, white whales and false killer whales are collected by field sampling. Then the time-frequency characteristics and distance resolution of the collected signals are analyzed and compared in time domain and frequency domain. The results show that the three marine mammals can achieve target detection by continuously transmitting click pulse trains. In this process, the pulse interval or signal waveform of the click pulses can be adjusted. The spectrum of a part of the click pulse signal of false killer whales has the special character of bimodal. Through the analyses of the ambiguity functions of the click signals of the three marine mammals, it is found that the distance resolution of bottlenose dolphins and white whales can reach millimeter scale; the false killer whales is relatively poor in distance resolution, which reaches centimeter scale. However, with the emergence of bimodal spectrum signals, the distance resolution of false killer whales is improved significantly and can also reach millimeter scale.
marine mammals; click pulse signal; ambiguity function
Q62
A
1000-3630(2019)-01-0024-08
10.16300/j.cnki.1000-3630.2019.01.004
2018-01-06;
2018-02-10
李赫(1993-), 男, 辽宁丹东人, 硕士研究生, 研究方向为水声物理。
高大治, E-mail: dzgao@ouc.edu.cn
