模拟升温和营养盐加富对大亚湾浮游生物群落代谢的影响*
2019-04-11谢福武宋星宇谭烨辉谭美婷黄亚东刘华雪
谢福武, 宋星宇, 谭烨辉, 谭美婷, 黄亚东, 刘华雪
模拟升温和营养盐加富对大亚湾浮游生物群落代谢的影响*
谢福武1, 2, 宋星宇3, 谭烨辉3, 谭美婷3, 黄亚东3, 刘华雪1
1. 中国水产科学研究院南海水产研究所, 广东省渔业生态环境重点实验室, 广东 广州 510300; 2. 海南省海洋与渔业科学院(海南省海洋开发规划设计研究院), 海南 海口 571126; 3. 中国科学院南海海洋研究所, 中国科学院热带海洋生物资源与生态重点实验室, 广东 广州 510301
文章结合模拟升温和营养盐加富, 于2016年8月和2017年1月研究了夏季和冬季大亚湾海域浮游生物群落代谢特征, 以期深入了解浮游生物对升温和富营养化作用的响应机制。结果表明: 升温和营养盐输入均会影响浮游生物总初级生产力(GPP)和群落呼吸率(CR), 且GPP对环境的响应更敏感。夏季GPP除在培养前期(24h)极高温条件下受到显著抑制外, 营养盐输入的影响比温度的影响作用更显著; 而冬季GPP和CR受升温效应的影响显著(<0.05)。总体看来, 升温效应和营养盐输入均能影响浮游生物群落碳代谢平衡, 导致海洋生态系统的稳定性降低, 进而影响海洋生物多样性以及渔业资源产出的数量和质量。
浮游生物; 群落代谢; 营养盐输入; 大亚湾
海洋浮游生物由浮游植物、浮游动物和浮游微生物构成, 通常悬浮在水层中随水流移动(王娜, 2014)。浮游生物群落代谢过程主要包括初级生产和群落呼吸两大基本生态过程, 而群落净生产力(net community production, NCP)大小决定生态系统呈现自养或异养状态(Ducklow et al, 2005)。浮游生物群落呼吸和初级生产能反映海洋生态系统中的碳代谢平衡, 其平衡过程对于海洋生态系统碳通量的分布格局与演变趋势是至关重要的, 是衡量海洋生态系统碳循环和营养物质动力学的重要指标。
水温上升引起浮游生物初级生产与群落呼吸贡献的潜在变化是一个较为复杂的过程, 可通过对观测数据的回归分析或者模拟全球气候变化的升温幅度来研究升温效应对浮游群落代谢特征的影响。一般认为, 温度升高在一定程度上能提高浮游生物总初级生产力和群落呼吸率水平(Regaudie‐de‐Gioux et al, 2012; Vaquer-Sunyer et al, 2013)。随着核电站数量的不断增加以及规模的不断扩展, 核电站温排水所引起的“热污染”对近海生态环境的影响日益受到重视, 这种由于温排水所导致的升温效应可使核电站邻近水域温度升高超过8℃, 而这种较高的升温幅度对周边水体浮游群落代谢特征的影响鲜有报道(Kim et al, 2007; Teixeira et al, 2012)。另外, 沿海核电站往往建设在人类活动影响较大的近海区域, 这些水域常伴随着营养物质输入及潜在的富营养化效应, 无论升温效应还是富营养化效应, 对水体代谢过程均有潜在的促进作用, 但这两个因子对近海初级生产与呼吸代谢的影响的综合效应如何, 两者的影响程度是否存在差异, 仍缺乏研究(Vaquer-Sunyer et al, 2015, 2016)。
大亚湾是广东省东南部近海的重要亚热带半封闭性海湾, 海水主要通过湾口与外海水进行交换, 湾内存在多个核电站, 此外水产养殖的发展与陆源输入增加也使得局部海域出现富营养化趋势, 而目前该海湾浮游群落代谢特征及其受升温与营养盐输入的综合影响状况尚不清楚(Song et al, 2009; Liu et al, 2011; Jiang et al, 2015)。本文通过模拟升温和营养盐加富实验相结合, 分析这两个因素对大亚湾浮游生物群落代谢的影响以及碳代谢平衡的响应规律, 以期深入了解环境变化背景下近海生态系统物质循环及能量流动的途径与传输效率, 完善生态系统整体功能和海洋生态系统碳通量的区域分布格局与演变趋势的评估。
1 材料与方法
1.1 实验设计
分别于2016年8月1日—10日和2017年1月4日—13日在大亚湾海域进行培养实验(图1), 其中M1位于近岸养殖区, 营养盐丰富, 赤潮频发; M2位于湾口, 营养盐含量不高, 水质良好。根据现场环境特征, 将模拟升温实验设计在M1, 交叉实验设计在M2 (表1)。
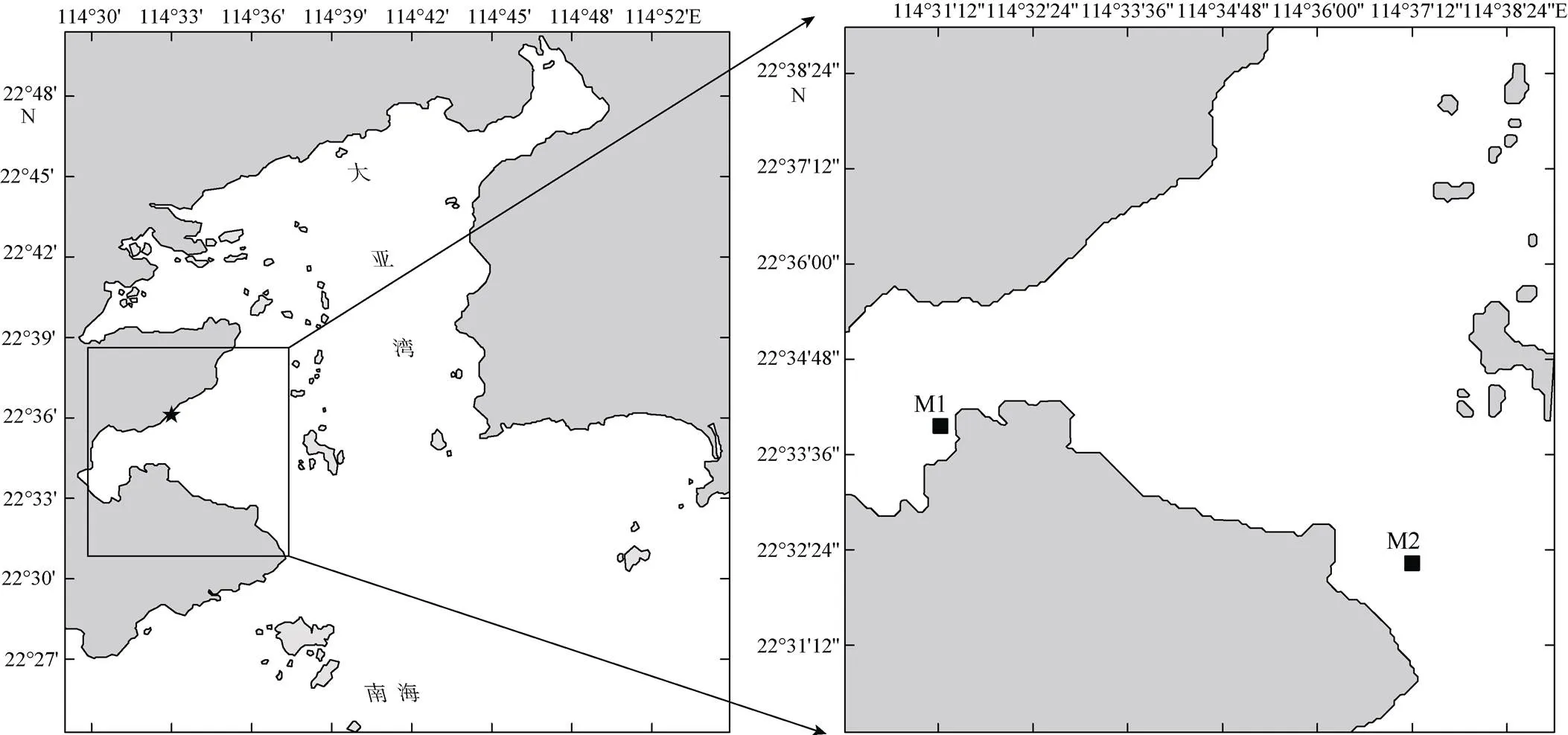
图1 现场温度调查和模拟实验采样站位分布图
左图中*为大亚湾核电站; 右图中M1和M2为实验站位
Fig. 1 Maps of sampling stations in Daya Bay
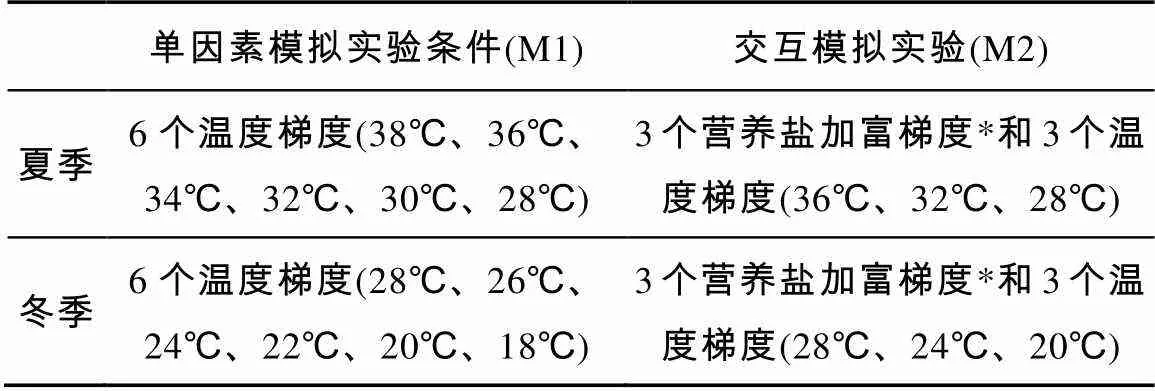
表1 大亚湾模拟实验设计
注: *营养盐加富梯度[低浓度组(LN): 不添加营养盐; 中浓度组(MN): 添加终浓度为10μmol·L-1的硝酸盐及0.6μmol·L-1的磷酸盐; 高浓度组(HN): 添加终浓度为100μmol·L-1的硝酸盐及6μmol·L-1的磷酸盐]
在M1和M2采集原始水样, 采用多组高精度低温恒温水浴模拟不同梯度的温度(控温精度0.05℃), 氙气灯模拟日光光源(光暗比设置与实验地点、季节的日照时长一致)进行培养, 温度梯度设置依据谢福武等(2018)夏冬季现场温排水的相关研究。
1) 模拟升温实验(SE): 根据夏季和冬季现场调查站位(M1)的实际温度结果, 分别设置6个温度(表1), 对采集的水样进行不同温度梯度的培养。
2) 模拟升温和营养盐加富交叉模拟实验(TSE): 夏冬季交叉实验设置3个温度梯度和3个营养盐加富梯度的交叉组合, 共9个梯度组。营养盐加富梯度为低浓度组LN (不添加营养盐)、中浓度组MN (终浓度为10μmol×L-1的硝酸盐和0.6μmol×L-1的磷酸盐)和高浓度组HN (终浓度为100μmol×L-1的硝酸盐和6μmol×L-1的磷酸盐)。
1.2 样品采集与测定
采样站点分布如图1所示, 通过采水器或虹吸法采集表层水, 水样不经处理直接分装至340mL溶氧瓶中(装样前用水样润洗瓶子, 用管插入瓶底加水样至溢出瓶口, 以排除气泡, 轻轻盖紧瓶盖), 每一梯度组的黑瓶和白瓶水样分别设置3个平行样。
培养实验: 将水样均置于低温恒温水浴装置中进行培养, 室内模拟实验水样进行72h连续培养, 并从0h开始中, 每隔24h进行样品测定工作。
零时间组: 直接采用Presens precision sensing Fibox4光纤溶氧仪测定初始溶解氧瓶(带感应芯片装置)中的水样, 记录读数。
白瓶组: 将已测定的0h培养水样进行特定时间的光照培养, 测定其溶解氧读数。
黑瓶组: 将已测定的0h培养水样需用锡箔纸包裹再进行特定时间的黑暗培养,测定相应溶解氧读数。
温度、盐度、pH等参数用YSI6600环境在线监测仪用于现场测量。叶绿素及营养盐的测定分别参照(Parsons et al, 1984; Kirkwood et al, 1996), 采用Turner-Design 10型荧光计和紫外分光光度计进行测量。
1.3 数据分析
浮游生物群落代谢测定: 群落呼吸速率(CR, 单位: mgC·m-3·d-1)、群落总初级生产力(GPP, 单位: mgC·m-3·d-1)和群落净初级生产力(NCP, 单位: mgC·m-3·d-1)的计算公式按照García-Martín等(2014)的方法。
CR=初始瓶溶解氧-黑瓶溶解氧
GPP=白瓶溶解氧-黑瓶溶解氧
NCP=白瓶溶解氧-初始瓶溶解氧
其中, 1mg×L-1(O)相当于(1/氧的相对原子质量)×碳的相对原子质量×1000(C)。
本文站位分布图采用Surfer11.0软件进行绘制, 浮游生物群落代谢和初级生产力等参数之间的差异性采用SPSS 19.0统计软件进行显著性方差分析处理, SigmaPlot 10.0进行图表绘制。
2 结果
2.1 环境理化参数
夏季湾口区(M2)呈现高温高盐寡营养特征, 而近岸养殖区(M1)海水温度和盐度较低(表2)。M1和M2的磷酸盐含量(DIP)相对较低, 溶解无机氮(DIN)含量的整体水平较高, M1站DIN浓度为35.46μmol·L-1, 浮游植物叶绿素(Chl)含量高达5.37μg·L-1。冬季水体垂直混合较均匀, DIP和DIN浓度整体水平要高于夏季。浮游植物叶绿素含量以M2站居高, 为1.43μg·L-1, 夏冬两季近岸养殖区(M1)和湾口外海区(M2) DIN/DIP的Redfield比均大于16 : 1, 总体呈潜在的磷限制状态。
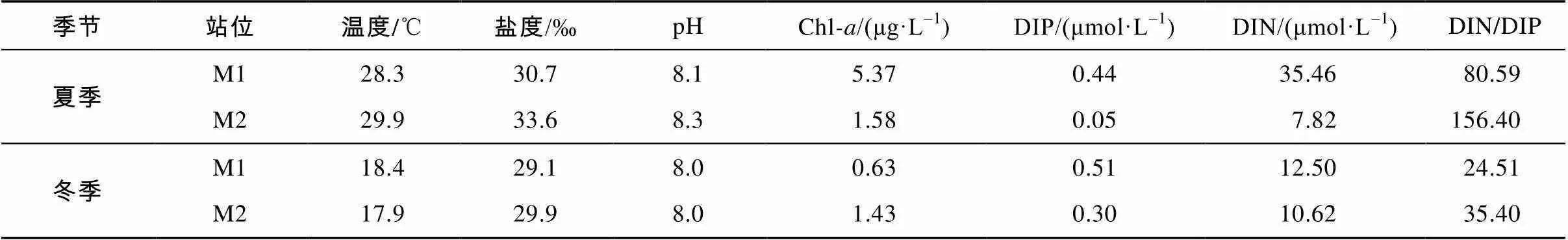
表2 夏冬两季大亚湾表层站位环境参数
2.2 模拟升温实验(SE)实验结果
夏季群落呼吸率(CR)随温度梯度增加呈先增后减的趋势, 随着培养时间的延长, 群落呼吸速率不断升高(图2), 在34℃条件下, 培养72h时CR值最高(486.38mgC·m-3·d-1), 但在极高温(38℃)环境下, 群落呼吸速率低于其他培养组, 不同温度组CR值存在显著性差异(<0.05)。在培养24~48h时, 总初级生产力(GPP)和净生产力(NCP)随升温表现为先增后减的趋势, 于30~32℃条件下GPP值最高, 达到2503.50mgC·m-3·d-1。在培养后期(72h)阶段, 各温度梯度GPP水平整体呈不同程度的上升趋势, NCP值整体呈自养状态。冬季CR值和GPP值均随升温以及培养时间的延长不断增加, 在28℃条件下, 72h时群落代谢速率最高, CR值和GPP值分别为236.25mgC·m-3·d-1和1143.75mgC·m-3·d-1。低温条件下(18℃, 72h), CR值和GPP值最低, 分别为101.25mgC·m-3·d-1和198.75mgC·m-3·d-1。与CR值和GPP值的响应情况类似, 冬季NCP值随温度升高总体呈上升趋势, 并在28℃时达到最高值。整体上夏季浮游生物群落生产和代谢强度强于冬季, 且两个季节不同温度梯度组整体均呈正代谢平衡状态。
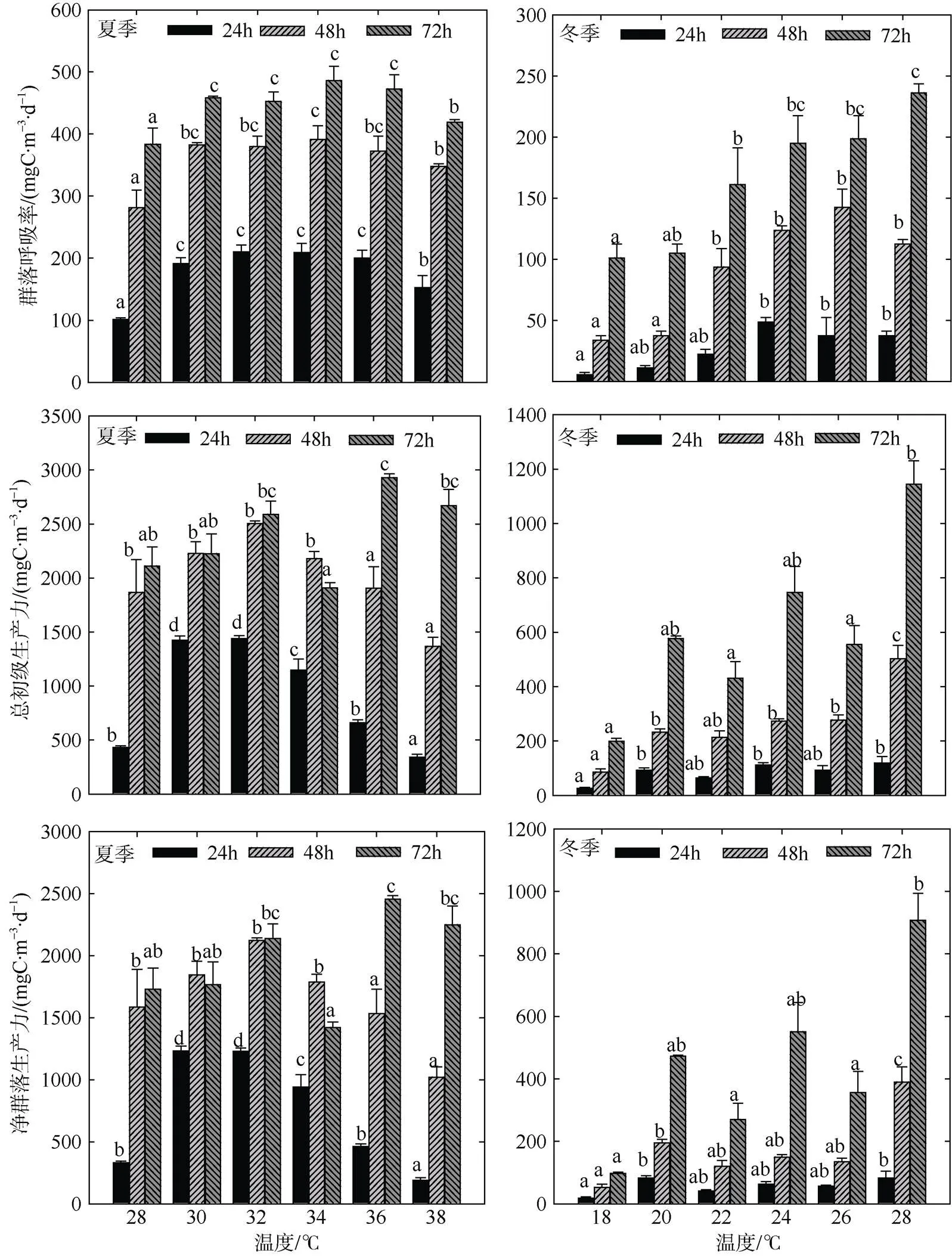
图2 模拟升温实验浮游生物群落代谢特征
标注不同字母的为同一时间内(如24h、48h、72h)的各温度组之间存在显著性差异(<0.05), 相同字母的差异性不显著
Fig. 2 Features of plankton community metabolism in simulated experiments
2.3 双因素交互模拟实验(TSE)实验结果
夏季CR值在培养前期(24h)整体差异较小(图3), 各梯度组CR值在150.00~318.75mgC·m-3·d-1范围内波动, 不同温度梯度以及不同浓度营养盐短期内(24h)对于CR值的影响均不显著(<0.05)。不同梯度组GPP值变动范围在101.25~3150.00 mgC·m-3·d-1之间, 差异较大(图4), 夏季短时间内(24h) GPP值总体平均值随升温而降低, 却随营养盐浓度的升高而增加。方差分析结果显示, GPP值受温度的影响要比营养盐明显, 特别是高温条件(36℃)下对GPP的影响显著于其他两个温度组(<0.05), 而受不同浓度营养盐水平的影响均不显著(<0.05)。
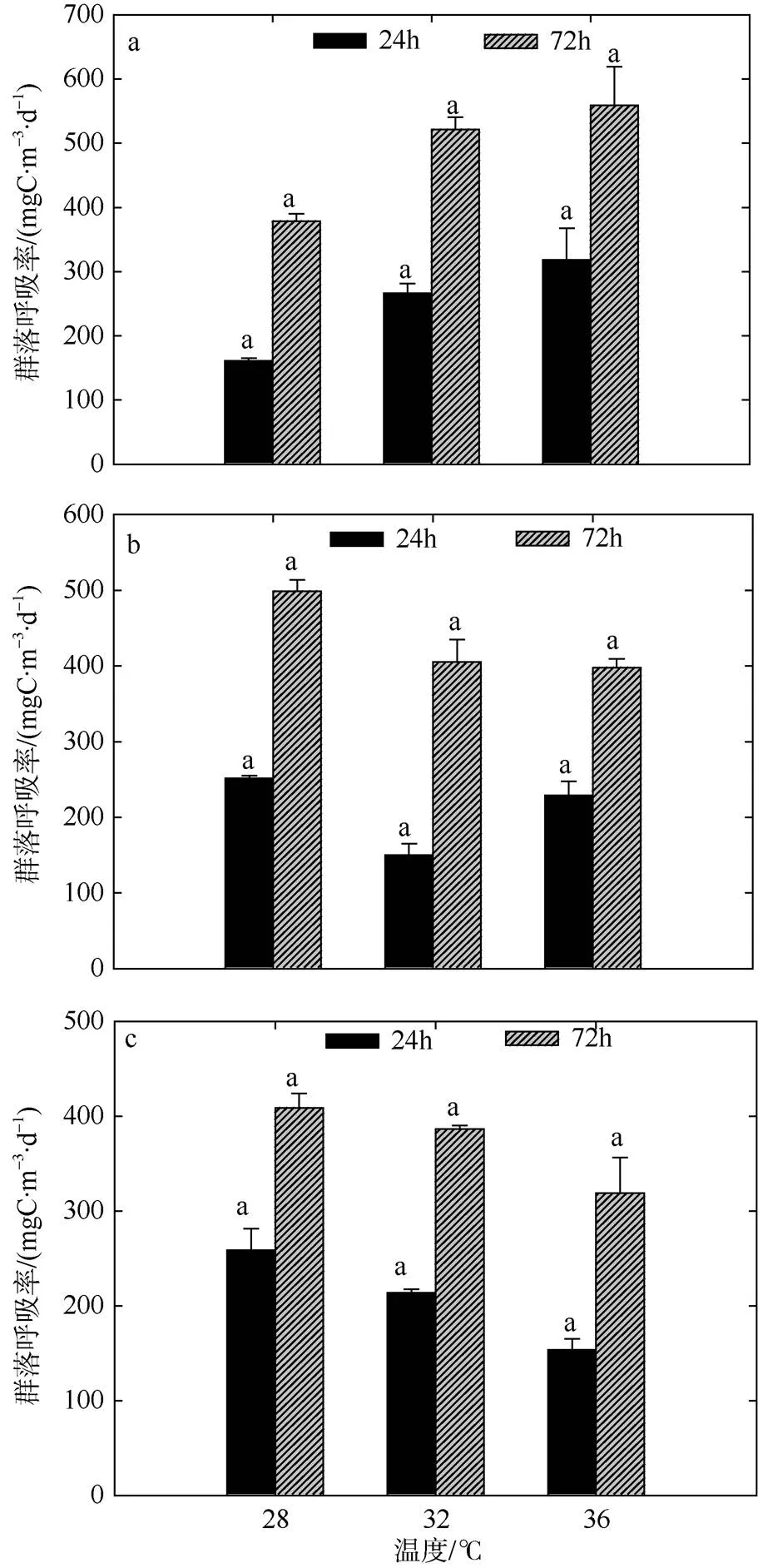
图3 夏季双因素模拟实验浮游生物群落呼吸率的变化
a. 低浓度营养盐; b. 中浓度营养盐; c. 高浓度营养盐。标注不同字母表示温度组之间存在显著性差异(<0.05), 相同字母的差异性不显著; 标注“*”的为营养盐组之间存在显著性差异(<0.05), 不标注“*”的为差异性不显著
Fig. 3 Variation of plankton CR of two-factors simulated experiments in summer. Significant differences among the temperature groups are indicated by using different superscripts (<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”
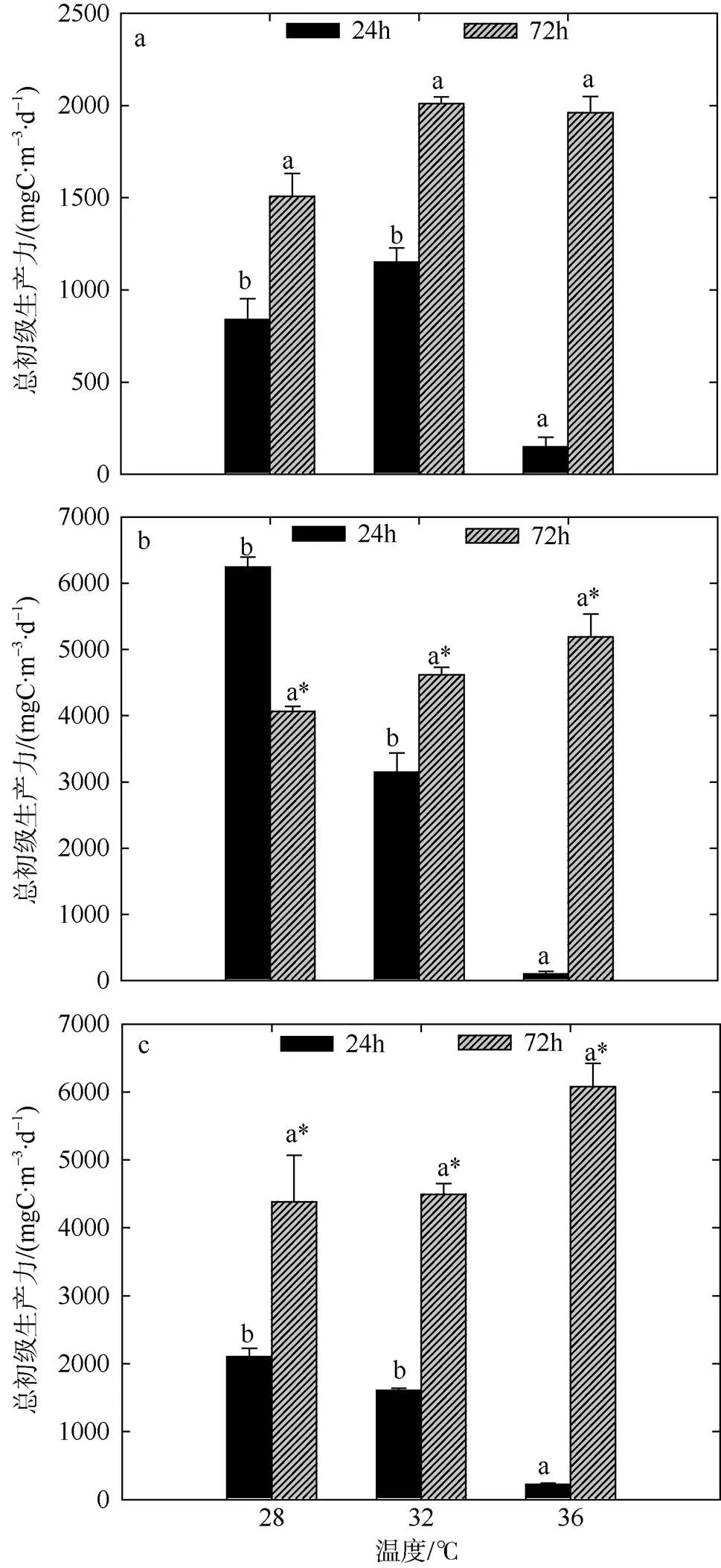
图4 夏季双因素模拟实验浮游生物群落总初级生产力的变化
a. 低浓度营养盐; b. 中浓度营养盐; c. 高浓度营养盐。标注不同字母表示温度组之间存在显著性差异(<0.05), 相同字母的差异性不显著; 标注“*”表示营养盐组之间存在显著性差异(<0.05), 不标注“*”为差异性不显著
Fig. 4 Variation of plankton GPP of two-factors simulated experiments in summer. Significant differences among the temperature groups are indicated by using different superscripts (<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”
高温条件(36℃)不同浓度营养盐环境下, NCP值整体水平较低(图5), 尤其是低营养和中营养组分别为-168.75mgC·m-3·d-1、-127.50mgC·m-3·d-1, 总生产/总呼吸比的平均值为5.70, 水体呈负代谢平衡状态(表3)。在培养后期(72h), 不同梯度组CR值随时间的延长均有提升, 各组之间差异不明显, 变动范围在318.75~558.75mgC·m-3·d-1之间。
GPP相较于培养前期也有所提升, 但不同梯度组间仍有较大差异, GPP在1507.50~6078.75mgC·m-3·d-1之间波动。方差分析结果表明, 培养后期(72h)营养盐的作用要显著于温度, 尤其是中营养和高营养组均显著于低营养组(<0.05), 不同温度间GPP水平差异不显著。不同梯度组间NCP以及GPP/CR整体随时间均有升高。
冬季不同温度和营养盐梯度组间CR于培养前期(24h)存在差异, CR整体水平在30.00~330.00mgC·m-3·d-1的范围内波动(图6)。CR在短时间内受温度的作用要显著于营养盐, 如28℃组CR显著于20℃组(<0.05)。GPP受不同温度以及营养盐的影响, 各梯度组间也具有一定差异(图7), GPP整体变动范围在157.50~ 581.25mgC·m-3·d-1之间。冬季GPP水平随升温而上升, 随营养盐浓度的增加呈小幅度降低趋势, 且培养前期(24h) GPP受温度和营养盐的影响均不显著。
各梯度组NCP值整体呈正代谢平衡状态(图8), 在培养后期(72h), CR值和GPP值均较前期有升高趋势, 分别在183.75~585.00mgC·m-3·d-1和1507.50~ 6078.75mgC·m-3·d-1的范围内变化。方差分析表明, 培养后期(72h)温度对于CR值和GPP值的影响均要显著于营养盐(<0.05), 而不同浓度营养盐间的差异不显著。不同梯度组的NCP水平以及总生产/总呼吸比值均随时间呈上升趋势。
3 讨论
3.1 升温效应对浮游生物群落代谢的影响
生态代谢理论认为升温会对浮游生物群落的GPP和CR存在调节作用, 初级生产和呼吸代谢这两大基本生态过程受温度的作用明显(王娜, 2014; Vaquer-Sunyer et al, 2015)。温度升高在一定程度上能提高浮游生物GPP值和CR值(Nguyen et al, 2012), 而这些结论一般是基于对观测数据的回归分析(Regaudie‐de‐Gioux et al, 2012), 模拟实验中温度的变化范围一般针对气候变化所引起的升温作用(Wohlers et al, 2009; Vaquer-Sunyer et al, 2013)。
a. 低浓度营养盐; b. 中浓度营养盐; c. 高浓度营养盐。标注不同字母表示温度组之间存在显著性差异(<0.05), 相同字母的差异性不显著; 标注“*”表示营养盐组之间存在显著性差异(<0.05), 不标注“*”表示差异性不显著
Fig. 5 Variation of plankton NCP of two-factors simulated experiments in summer. Significant differences among the temperature groups are indicated by using different superscripts (<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”
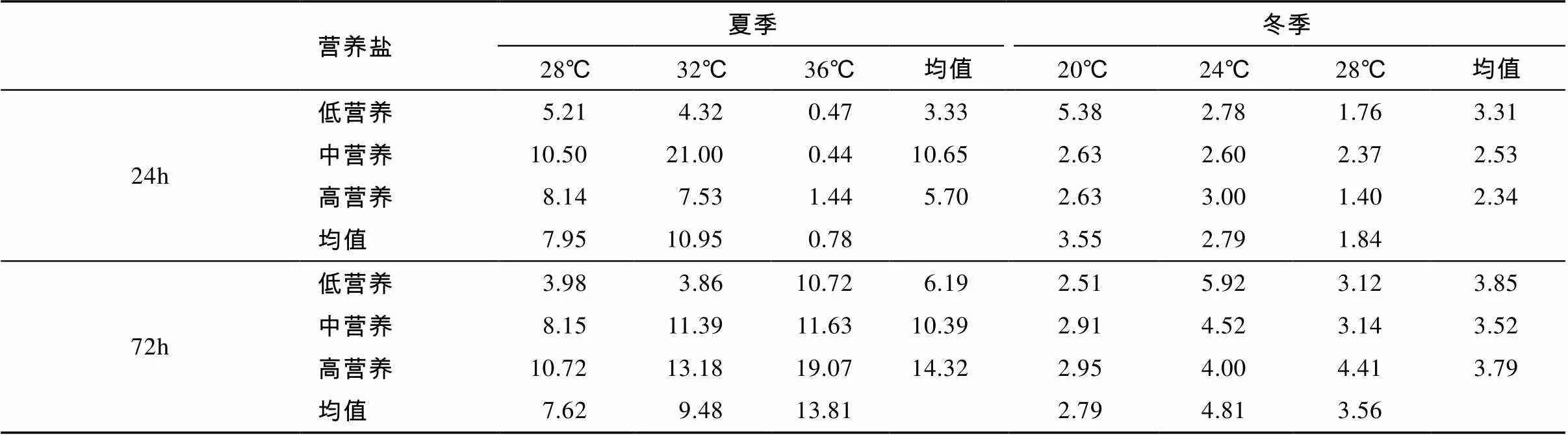
表3 TSE浮游生物总生产与总代谢的比值(GPP/CR)

图6 冬季双因素模拟实验浮游生物群落CR值变化
a. 低浓度营养盐; b. 中浓度营养盐; c. 高浓度营养盐。标注不同字母表示温度组之间存在显著性差异(<0.05), 相同字母的差异性不显著; 标注“*”表示营养盐组之间存在显著性差异(<0.05), 不标注“*”的为差异性不显著
Fig. 6 Variation of plankton CR of two-factors simulated experiments in winter. Significant differences among the temperature groups are indicated by using different superscripts (<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”
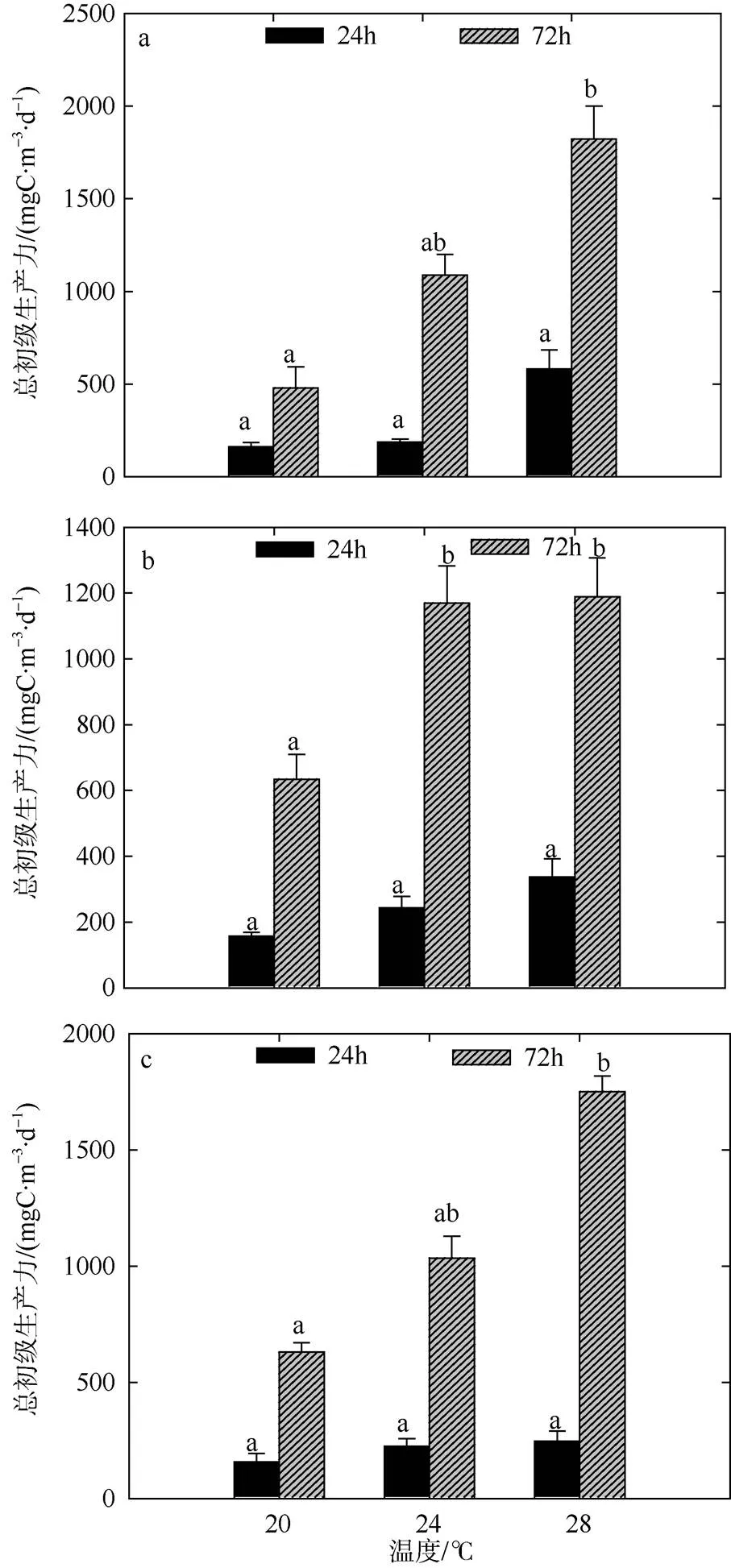
图7 冬季双因素模拟实验浮游生物群落GPP值变化
a. 低浓度营养盐; b. 中浓度营养盐; c. 高浓度营养盐。标注不同字母表示温度组之间存在显著性差异(<0.05), 相同字母的差异性不显著; 标注“*”的为营养盐组之间存在显著性差异(<0.05), 不标注“*”的为差异性不显著
Fig. 7 Variation of plankton GPP of two-factors simulated experiments in winter. Significant differences among the temperature groups are indicated by using different superscripts (<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”
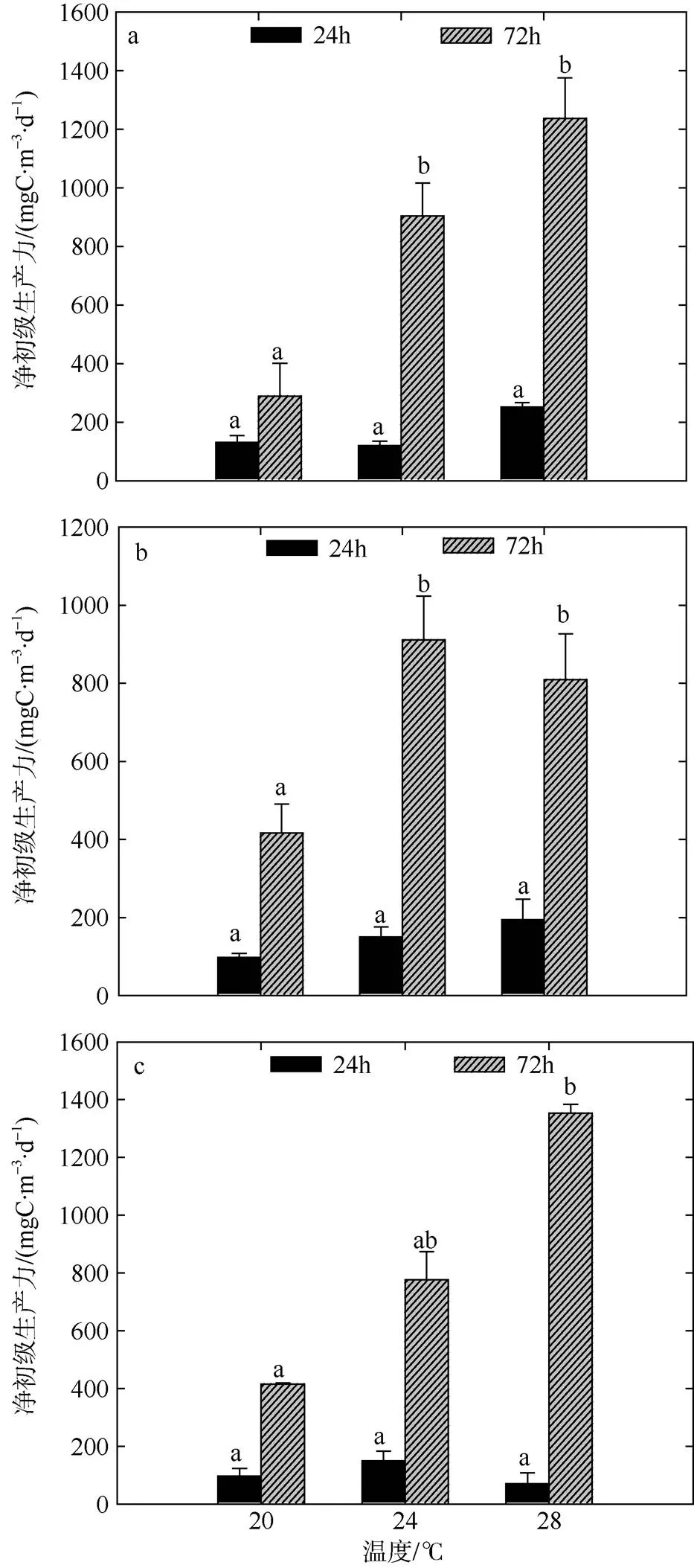
图8 冬季双因素模拟实验浮游生物群落NCP值变化
a. 低浓度营养盐; b. 中浓度营养盐; c. 高浓度营养盐。标注不同字母表示温度组之间存在显著性差异(<0.05), 相同字母的差异性不显著; 标注“*”的为营养盐组之间存在显著性差异(<0.05), 不标注“*”的为差异性不显著
Fig. 8 Variation of plankton NCP of two-factors simulated experiments in winter. Significant differences among the temperature groups are indicated by using different superscripts (<0.05), while no significant difference is shown using the same superscript. LN: low nutrients, MN: medium nutrients, HN: high nutrients. Significant differences among the nutrients groups are marked by “*”, while no significant difference is shown without “*”
本文的现场观测与模拟实验均针对较广的温度范围来研究浮游生物群落代谢所受的潜在影响, 高温条件下群落代谢速率受到明显的抑制作用, GPP值较低, 水体则呈异养代谢状态, 这种高温抑制浮游群落代谢的现象在之前研究鲜有报道。其作用机制可能是由于温排水的适度升温提高浮游生物体内酶的活性或者升温加速溶解有机物的循环利用, 使代谢速率加快,但极高温条件下则抑制其酶活性, 并破坏其类囊体结构使浮游藻类光合作用过程受阻, 抑制浮游动物生长, 降低浮游生物群落代谢速率(El-sabaawi et al, 2006; 蔡泽富, 2011)。本文结果显示大亚湾海域浮游生物初级生产和呼吸代谢极易受到高温效应的影响, 夏冬两季浮游生物群落代谢速率均在温排水影响区的适温(29~30℃)水域内强度最强, 浮游生态系统呈自养代谢状态, 与以往报道一致(Wohlers et al, 2009)。
夏季极高温组(36~38℃)水体代谢水平在培养实验初期受到抑制, 但培养后期, 极高温组GPP快速上升, 呈较高水平。其原因是, 极高温组在培养初期受高温的抑制, 浮游藻类生产力水平较低, 利用营养盐较少, 随着培养时间的延长, 更适应高温水体环境的粒径小的光合细菌很可能变成浮游植物的优势类群, 并可充分利用水体中多余的营养盐进行快速增长, 促进了GPP水平的提升(谢福武等, 2018)。而培养前期其他温度组浮游藻类快速增长, 水体中营养盐被过度消耗, 导致后期营养盐浓度降低不能供给浮游藻类生长, 因此在培养实验后期其代谢水平反而较低。总体而言, 模拟实验结果表明浮游生物群落代谢特征沿温度梯度存在规律性的差异特征。
3.2 温度和营养盐对浮游生物群落代谢平衡的综合影响
浮游生物群落代谢平衡特征存在潜在的季节性差异, 如王娜(2014)研究发现, 夏季厦门港近岸水体GPP值和CR值最高, 冬季的GPP值和CR值较低, 但对于不同季节代谢水平的影响因素及其潜在的季节差异尚不清楚。
温度与营养盐均是影响浮游群落代谢水平的重要因素(Vaquer-Sunyer et al, 2016), 但其对群落生产和呼吸代谢的影响程度可能存在差异。本文研究表明, 夏季浮游生物CR值受温度和营养盐的影响均不显著, 冬季CR值受升温的作用要显著于营养盐。对于GPP而言, 除了培养初期受夏季极高温环境影响存在明显抑制作用外, 夏季受营养盐的促进作用显著强于温度作用, 而冬季GPP整体受温度的影响显著于营养盐。这种环境因子潜在影响力的差异很可能与该海域环境背景的季节性差异有关。在夏季, 大亚湾水体温度较高, 水体易形成层化结构, 磷酸盐和溶解态无机氮含量较低, 因此在夏季这种相对高温、低营养盐的环境下, 大亚湾浮游生物类群的初级生产更易受营养盐输入的促进作用。即使在极高温条件下, 短时间内的剧烈升温能影响光合进程中酶促反应或呼吸反应强度等生理过程(余立华, 2006)进而抑制GPP值, 后期逐步适应了高温环境的浮游生物群落在有足够营养盐补充的情况下仍可能快速繁殖增长。
冬季大亚湾温度相对较低, 由于季风与高盐水团的交互作用, 海水的垂直混合作用剧烈, 上层水体的营养物质可通过垂直混合作用得以补充(丘耀文等, 2005; Fu et al, 2016), 这种相对低温、高营养盐环境下, 温度升高更易影响CR值和GPP值, 因此冬季升温对浮游生物群落代谢的影响显著于营养盐作用, 并造成两个环境因子对浮游群落代谢影响程度的季节差异。
3.3 浮游生物群落代谢的潜在生态效应
升温作用能促进浮游生物呼吸速率加快, 导致溶解态氧被快速消耗引起水体缺氧。此外, 营养盐输入提高GPP水平, 促使浮游藻类快速增长并积累有机物质, 然后藻体大面积死亡, 在分解过程中大量消耗水中的溶解氧, 是驱动水体贫氧的另一重要因素(Vaquer-Sunyer et al, 2015, 2016)。气候变化所引起的升温和有机营养物输入均能影响浮游生物代谢速率, 如Vaquer-Sunyer等(2015, 2016)研究发现波罗的海温度变化和有机营养物输入均能导致浮游生物初级生产和呼吸代谢的变化, 是导致该海域水体缺氧的主要因素。
无论是升温效应还是营养盐输入均对浮游生物群落生产代谢状态产生影响, 尤其是现场及模拟实验中, 极高的水温均明显抑制了浮游植物和光合细菌的固碳作用, 并使水体呈负代谢平衡, 易引起水体溶解氧含量下降, 呈贫氧状态, 尤其是夏季效应最为明显。这种水体贫氧化还可能进一步影响海洋生物生长, 对生物资源产出产生负面作用, 导致生态系统功能下降(Song et al, 2015)。浮游生物GPP/CR比值是衡量系统发育程度和成熟度的重要指标, 成熟的生态系统中, 该比值逐渐接近于1, 说明没有多余的生产量可供系统再利用(徐姗楠等, 2008; Chen et al, 2011)。模拟实验发现升温与营养盐输入会造成GPP/CR比值的较大波动, 且GPP/CR常明显偏离理论上的平衡值1, 这也说明了升温效应以及营养盐输入均能通过影响浮游生物群落代谢来影响水体生态系统的稳定性, 造成海湾浮游生态系统的稳定性下降。此外, 升温和富营养化等环境作用还可能改变浮游生态系统的食物网结构与食物链传递途径(Culler, 2013; Lewandowskaet al, 2014; Song et al, 2015), 这些综合的生态效应均可能进一步对生态系统功能及生物资源产出造成负面影响。
4 结论
夏季和冬季大亚湾浮游生物群落代谢均随水温上升而增加趋势, 特别是30~32℃左右温度有显著促进作用, GPP值和CR值具有较高水平, 但在极高温环境下(36~38℃)热效应抑制作用明显, 浮游生物群落代谢和初级生产水平下降或呈异养代谢状态。
升温和营养盐输入均能促进浮游生物的GPP值和CR值, 浮游生物初级生产速率的增长整体高于呼吸代谢速率。升温和营养盐输入对大亚湾浮游生物群落代谢水平的影响力存在季节性差异: 夏季整体上受营养盐的影响作用则显著于温度; 冬季受升温的影响显著于营养盐输入的影响。这主要是由自然水体温度、营养盐含量等环境背景的季节性差异决定的。
升温效应和营养盐输入均可通过影响浮游群落代谢平衡状况, 导致海湾生态系统的稳定性下降, 脆弱性增加。
蔡泽富, 2011. 温排水对象山港浮游生态系统的影响及其围隔实验研究[D]. 上海: 上海海洋大学. CAI ZEFU, 2011. The study of affects of thermal effluent on pelagic ecosystem by mesocosm experiments in Xiang Shan Bay[D]. Shanghai: Shanghai Ocean University (in Chinese with English abstract).
丘耀文, 王肇鼎, 朱良生, 2005. 大亚湾海域营养盐与叶绿素含量的变化趋势及其对生态环境的影响[J]. 台湾海峡, 24(2): 131–139. QIU YAOWEN, WANG ZHAODING, ZHU LIANGSHENG, 2005. Variation trend of nutrient and chlorophyll contents and their effects on ecological environment in Daya Bay[J]. Journal of Oceanography in Taiwan Strait, 24(2): 131–139 (in Chinese with English abstract).
王娜, 2014. 南海及福建近岸水体中群落与细菌呼吸的研究[D]. 厦门: 厦门大学. WANG NA, 2014. Community and bacterial respiration in the South China Sea and Fujian coastal waters[D]. Xiamen: Xiamen University (in Chinese with English abstract).
谢福武, 刘华雪, 黄洪辉, 等, 2018. 大亚湾浮游植物粒级结构对温排水和营养盐输入的响应[J]. 热带海洋学报, 37(3): 55–64. XIE FUWU, LIU HUAXUE, HUANG HONGHUI, et al, 2018. Effects of thermal discharge and nutrients input on size structure of phytoplankton in Daya Bay[J]. Journal of Tropical Oceanography, 37(3): 55–64 (in Chinese with English abstract).
徐姗楠, 陈作志, 何培民, 2008. 杭州湾北岸大型围隔海域人工生态系统的能量流动和网络分析[J]. 生态学报, 28(5): 2065–2072. XU SHANNAN, CHEN ZUOZHI, HE PEIMIN, 2008. Energy flux and network analysis for an artificial ecosystem of a large enclosed sea area in North Hangzhou Bay[J]. Acta Ecologica Sinica, 28(5): 2065–2072 (in Chinese with English abstract).
余立华, 2006. 秋季长江口不同辐照和氮、磷浓度水平下浮游植物营养盐吸收动力学及生长变化研究[D]. 上海: 华东师范大学. YU LIHUA, 2006. Nutrient absorbing kinetics and growth changes of phytoplankton controlled by different irradiance, Nitrate and phosphate concentration levels on autumn in Yangtze River estuary[D]. Shanghai: East China Normal University (in Chinese with English abstract).
CHEN ZUOZHI, QIU YONGSONG, XU SHANNAN, 2011. Changes in trophic flows and ecosystem properties of the Beibu Gulf ecosystem before and after the collapse of fish stocks[J]. Ocean & Coastal Management, 54(8): 601–611.
CULLER L E, 2013. Temperature effects on consumer-resource species interactions: Integrating thermal physiology and community ecology[D]. Dartmouth: Dartmouth College.
DUCKLOW H W, MCCALLISTER S L, 2005. The biogeochemistry of carbon dioxide in the coastal oceans[M]//ROBINSON A R, BRINK K H. The Sea. Cambridge, MA: Harvard University Press, 13: 269–315.
EL-SABAAWI R, HARRISON P J, 2006. Interactive effects of irradiance and temperature on the photosynthetic physiology of the pennate diatom(bacillariophyceae) from the Northeast subarctic Pacific[J]. Journal of Phycology, 42(4): 778–785.
FU MINGZHU, WANG ZONGLING, LI YAN, et al, 2016. Phytoplankton biomass size structure and its regulation in the Southern Yellow Sea (China): seasonal variability[J]. Continental Shelf Research, 29(18): 2178–2194.
GARCÍA-MARTÍN E E, SERRET P, LEAKEY R J G, 2014. Plankton community and bacterial metabolism in Arctic sea ice leads during summer 2010[J]. Deep Sea Research Part I: Oceanographic Research Papers, 92: 152–161.
JIANG ZHAOYU, WANG YOUSHAO, CHENG HAO, et al, 2015. Spatial variation of phytoplankton community structure in Daya Bay, China[J]. Ecotoxicology, 24(7–8): 1450–1458.
KIM Y H, AHN J K, YOON H D, et al, 2007. Effects of heated effluents on the intertidal macroalgal community near Gori Nuclear Power Plant[J]. Algae, 22(4): 297–304.
KIRKWOOD D S, AMINOT A, CARLBERG S R, 1996. The 1994 quasimeme laboratory performance study: nutrients in seawater and standard solutions[J]. Marine Pollution Bulletin, 32(8–9): 640–645.
LEWANDOWSKA A M, BOYCE D G, HOFMANN M, et al, 2014. Effects of sea surface warming on marine plankton[J]. Ecology Letters, 17(5): 614–623.
LIU HUAXUE, SONG XINGYU, HUANG LIANGMIN, et al, 2011. Diurnal variation of phytoplankton community in a high frequency area of HABs: Daya Bay, China[J]. Chinese Journal of Oceanology and Limnology, 29(4): 800–806.
NGUYEN D, MARANGER R, TREMBLAY J É, et al, 2012. Respiration and bacterial carbon dynamics in the Amundsen Gulf, western Canadian Arctic[J]. Journal of Geophysical Research: Oceans, 117(C9): C00G16.
PARSONS T R, MAITA Y, LALLI C M, 1984. A manual of chemical and biological methods for seawater analysis[M]. Oxford: Pergamon Press: 158–161.
REGAUDIE‐DE‐GIOUX A, DUARTE C M, 2012. Temperature dependence of planktonic metabolism in the ocean[J]. Global Biogeochemical Cycles, 26(1): GB1015.
SONG XINGYU, HUANG LIANGMIN, ZHANG JIANLIN, et al, 2009. Harmful algal blooms (HABs) in Daya Bay, China: An in situ study of primary production and environmental impacts[J]. Marine Pollution Bulletin, 58(9): 1310–1318.
SONG XINGYU, LIU HUAXUE, ZHONG YU, et al, 2015. Bacterial growth efficiency in a partly eutrophicated bay of South China Sea: Implication for anthropogenic impacts and potential hypoxia events[J]. Ecotoxicology, 24(7–8): 1529–1539.
TEIXEIRA T P, NEVES L M, ARAÚJO F G, 2012. Thermal impact of a nuclear power plant in a coastal area in Southeastern Brazil: effects of heating and physical structure on benthic cover and fish communities[J]. Hydrobiologia, 684(1): 161–175.
VAQUER-SUNYER R, DUARTE C M, 2013. Experimental evaluation of the response of coastal mediterranean planktonic and benthic metabolism to warming[J]. Estuaries and Coasts, 36(4): 697–707.
VAQUER-SUNYER R, CONLEY D J, MUTHUSAMY S, et al, 2015. Dissolved organic nitrogen inputs from wastewater treatment plant effluents increase responses of planktonic metabolic rates to warming[J]. Environmental Science & Technology, 49(19): 11411–11420.
VAQUER-SUNYER R, READER H E, MUTHUSAMY S, et al, 2016. Effects of wastewater treatment plant effluent inputs on planktonic metabolic rates and microbial community composition in the Baltic Sea[J]. Biogeosciences, 13(16): 4751–4765.
WOHLERS J, ENGEL A, ZÖLLNER E, et al, 2009. Changes in biogenic carbon flow in response to sea surface warming[J]. Proceedings of the National Academy of Sciences of the United States of America, 106(17): 7067–7072.
Impact of simulated warming and nutrients input on plankton community metabolism in Daya Bay*
XIE Fuwu1, 2, SONG Xingyu3, TAN Yehui3, TAN Meiting3, HUANG Yadong3, LIU Huaxue1
1. Guangdong Provincial Key Laboratory of Fishery Ecological Environment, South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, China; 2. Hainan Academy of Ocean and Fisheries Sciences (Hainan Provincial Marine Development Plan and Design Research Institute), Haikou, 571126, China; 3. Key Laboratory of Tropical Marine Bio-resources and Ecology, South China Sea Institute of Oceanology, Chinese Academy of Sciences, Guangzhou 510301, China
Characters of plankton community metabolism of Daya Bay were studied using data collected in August 2016 and January 2017. The possible effects of nutrients input were discussed to understand the response mechanism of plankton to warming and eutrophication based on simulation experiments. The results indicated that both GPP (gross primary production) and CR (community respiration) were influenced by warming and nutrients input, and response of GPP to environment was seriously sensitive. The impact of GPP on nutrients input was more remarkably than that of temperature, except for being significantly inhibited in early cultured stage (24 h) with extremely high temperature conditions. In winter, both GPP and CR were significantly affected by the warming effect (<0.05). Overall, both warming effect and nutrients input could affect carbon metabolism of plankton community which would lead to the declining of marine ecosystem stability, thereby affecting marine ecosystem stability and biodiversity and quality of marine ecological environment and fishery resource output.
plankton; community metabolism; nutrients input; Daya Bay
2018-07-25;
2018-09-10. Editor: YIN Bo
National Key Research and Development Program of China (2017YFC0506302); National Basic Research Program (2015CB452904); National Natural Science Foundation of China (41276161)
P735.12; P735.53
A
1009-5470(2019)02-0048-10
10.11978/2018075
2018-07-25;
2018-09-10。殷波编辑
国家重点研发计划(2017YFC0506302); 国家重点基础研究发展计划(2015CB452904); 国家自然科学基金(41276161)
谢福武(1992—), 男, 海南省万宁市人, 从事海洋生态学研究。E-mail: xfw1021900977@163.com
宋星宇, 研究员。E-mail: songxy@scsio.ac.cn; 刘华雪, 副研究员。E-mail: liuhuaxue@scsfri.ac.cn
*感谢各位评审专家对文章的指导与修改。
SONG Xingyu. songxy@scsio.ac.cn; LIU Huaxue. liuhuaxue@scsfri.ac.cn
