自噬在骨稳态与骨质疏松症中的作用及机制研究进展
2019-04-10张欣然林开利
张欣然 林开利,2
骨质疏松症(osteoporosis,OP)是以骨量减少、骨脆性及骨折倾向增加为特征的渐进性、系统性骨代谢疾病,随着人口老龄化的加剧,其发病率逐年增高。OP的一个重要临床特征为病人早期的症状和体征缺乏或不明显,这通常会导致骨质疏松性骨折(osteoporotic fracture,OF)、骨折后病人活动受限及其他复杂并发症,严重影响病患的日常起居、自理能力,使个人与家庭蒙受生理和心理的双重重担。基础研究表明,骨髓间充质干细胞(bone marrow mesenchymal stem cells,BMSCs)、成骨细胞(osteoblasts,OB)及破骨细胞(osteoclasts,OC)等共同参与了OP的发生发展。自噬是真核细胞的高度保守行为,其与细胞稳态和应激、损伤修复和增殖、分化等密切相关。近年来研究表明自噬在骨代谢以及BMSCs、OB、OC等的增殖、分化、代谢与生理功能行使中有重要的调控作用,但其具体机制尚未明确。因此,我们拟通过文献综述,并从自噬在间充质干细胞(mesenchymal stem cells,MSCs)中的作用,自噬在成骨细胞和骨细胞中的作用,自噬在破骨细胞中的作用等几个方面去探索自噬与骨代谢的关系及其在OP发病机制中的作用。
一、文献检索策略
本文通过中文检索词“自噬”、“骨质疏松”、“间充质干细胞”、“成骨细胞”、“破骨细胞”以及英文检索词“autophagy”、“osteoporosis”、“mesenchymal stem cells”、“osteoblast”、“osteo⁃clast”在中国知网、万方数据库、维普数据库、PubMed、Web of Science等平台进行检索,筛选发表时间为近十年内,共检索到文献723篇。应用EndNote删除重复及无法获得全文文献115篇,最终纳入文献36篇,其中中文文献为3篇,英文文献为33篇(图1)。文献纳入标准为:①已正式发表的期刊文献;②文献内容与自噬、骨质疏松、骨稳态维持的相关细胞密切相关;③同类研究中质量、证据等级较高的文献;④语言为中文及英文的文献。文献排除标准为:①文献质量、证据等级较低的文献;②学位论文、会议类文献;③非中文及英文的文献;④无法获得全文的文献。
二、细胞自噬概述
自噬由Ashford等[1]于1962年通过电镜首次发现。当细胞面临外界环境压力(如营养物质缺乏、缺氧等)或者在病理状态下,细胞通过特殊包膜结构(例如单层膜或者双层膜)包裹长寿蛋白、有害物质或受损的细胞器等形成自噬体,随后通过溶酶体依赖途径进行分解代谢,而降解的产物用以实现胞内成分的循环再利用[2]。目前,根据相关物质从胞质运输到溶酶体过程中功能和途径的不同,将自噬分为了以下几类:巨自噬,小自噬和分子伴侣介导自噬。其中,巨自噬是自噬的主要类型,也是目前研究最多的一种类型,本文以下的自噬指巨自噬[3]。
通过基因筛选的相关研究,学者们发现了30多种自噬相关基因(autophagy⁃related gene,ATG),在自噬体的形成、调控过程中发挥重要作用[4]。这些基因编码的自噬相关蛋白在自噬早期募集到起始位点从而组成自噬的重要结构——双层隔离膜,随后内质网、线粒体、高尔基复合体等细胞器被隔离膜包裹,在这一过程中,Ⅲ类磷酸肌苷3激酶(classⅢphosphoinositide 3⁃kinase,classⅢ PI3K)/空泡分拣蛋白(vacu⁃olar protein sorting,Vps34)复合体被激活,然后与Beclin⁃1、ATG14、Vps15等蛋白一起形成多蛋白复合体,使双层膜结构不断延伸,并进一步招募ATG蛋白[5]。
ATG5、ATG7、ATG10和ATG12在扩展、延伸双层膜结构以形成完整的自噬体的过程中起重要作用。首先,ATG5和ATG12在ATG7和ATG10的介导下形成ATG5⁃ATG12复合体,随后结合ATG16L1,生成 ATG5⁃ATG12⁃ATG16L1 复合体[6]。微管相关蛋白轻链⁃3(microtubule⁃associated protein 1 light chain 3,LC3)前体首先由半胱氨酸蛋白酶ATG4切割,形成LC3⁃Ⅰ,然后通过ATG7、ATG10和ATG5⁃ATG12⁃ATG16L1复合体介导,LC3⁃Ⅰ与磷脂酰乙醇胺(Phosphatidylethanol⁃amine,PE)结合,生成LC3⁃Ⅱ[7]。LC3⁃Ⅱ是检测自噬形成的可靠蛋白质标记物,包括了合成过程中的3种重要的形式:未处理形式(the unprocessed form,pro⁃LC3)、蛋白水解形式(the proteolytically,LC3⁃Ⅰ)和PE-共轭形式(the PE⁃conjugat⁃ed form,LC3⁃Ⅱ)。最后,在自噬溶酶体形成后,自噬体膜以及包封在自噬体中的物质等被溶酶体内的蛋白酶降解,并释放至胞浆中以重新利用[8](图2)。
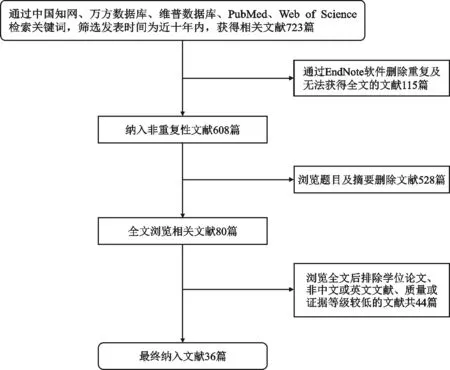
图1 文献筛选流程图
双层膜介导的胞内溶酶体降解这一过程并不复杂,自噬作为细胞内的“管家”,在维持细胞内环境稳定性和确保细胞正常生理功能方面扮演着不可或缺的角色[9]。然而,在某些病理因素或者疾病发生过程中,自噬的功能可能会发生改变或者被异常激活,甚至发生自噬过度的情况而引起细胞凋亡或者死亡。生理情况下,细胞自噬可以将受损或者衰老的细胞器清除、更新来维持细胞稳态。当细胞处于营养物质缺乏,或者应激状态(如药物作用、氧化应激、内质网应激等)下,细胞内自噬水平会升高,各种刺激引起的细胞器损伤及时被消除,细胞的正常新陈代谢和多种生理功能得以维持[10]。然而,自噬的这种代偿作用有一定限度,如果外界的刺激或者应激状态不能及时改善或者去除,一旦自噬的消耗超过了细胞的耐受阈值,可能导致一种不同于凋亡的程序性细胞死亡(Ⅱ型程序性细胞死亡),乃至诱发一些疾病[11]。
三、自噬在骨稳态中的调节作用

图2 自噬的发生过程(绘图作者:张欣然)
骨骼能够为躯体提供支撑和保护作用,但是骨功能作用的发挥需要骨形态、骨量以及骨质等的长期稳定。骨骼处于骨形成以及骨吸收的动态平衡(即骨稳态)中,维持骨骼稳态需要多种类型细胞(OB、OC、骨细胞等)、激素、因子等密切配合。其中,OB是主要发挥合成骨基质作用的细胞,其由MSCs分化而来;OC是具有骨吸收作用的高度分化的多核巨细胞,源自造血干细胞;骨细胞是构成骨质的重要细胞,骨细胞能够通过感知应力和激素水平等的变化来调节OB及OC,其包封在矿化的骨基质中、由OB终末期分化而来(图3)。越来越多的研究表明:自噬与骨稳态具有相关性,参与调控MSCs、成骨、破骨以及骨细胞[5]。
(一)自噬在MSCs中的作用
MSCs具有自我更新和多潜能分化能力,可以增殖、分化成多种类型细胞,例如OB、成软骨细胞和成脂肪细胞,BMSCs是一种存在于骨髓中的MSCs。骨组织发育、骨代谢、骨改建、骨修复和再生过程中所需要的成骨前体细胞,主要来源于 BMSCs[12]。2014年,Song等[13]研究表明H2O2介导的BMSCs的氧化应激能够造成细胞的大量死亡,此时细胞内自噬水平的提高可以促进BMSCs的成活。Nuschke等[14]观察到在未分化的BMSCs中有大量未降解的自噬空泡聚集或自噬体聚集;同时在BMSCs早期成骨分化过程中,BMSCs的胞体内也有大量自噬体聚集,这说明自噬体可以作为能量快速产生的来源。自噬还参与维持来源于脐带血的MSCs干性[15]。Pantovix等[16]研究表明单磷酸腺苷激活蛋白激酶(Ad⁃enosine 5′⁃monophosphate⁃activated protein kinase,AMPK)通路通过早期抑制哺乳动物类雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)和晚期激活蛋白激酶B(protein ki⁃nase B,AKT)/mTOR调节自噬,从而调控MSCs成骨分化。Liu等[17]通过应用自噬抑制剂[如3-甲基腺嘌呤(3⁃methylade⁃nine,3⁃MA)等]进一步研究了自噬在BMSCs中的作用:通过在BMSCs的培养过程中加入自噬抑制剂3-MA及氯喹,可见抑制剂组碱性磷酸酶(alkaline phosphatase,ALP)阳性细胞数量及大小均下降;在加入抑制剂21 d后,茜素红染色结果显示细胞矿化能力也下降。这些结果说明自噬抑制剂会影响BM⁃SCs的成骨分化及矿化能力。关于人牙髓间充质干细胞(hu⁃man dental pulp mesenchymal stem cells,hDP⁃MSC)的研究结果也印证了这个结论:自噬抑制剂巴弗洛霉素A1(bafilomy⁃cin A1,BA1)、氯喹以及氯化铵可以抑制hDP⁃MSC的分化[16]。
(二)自噬在OB和骨细胞中的作用
OB由MSCs分化而来,是特殊化的MSC细胞,负责骨形成。在骨形成过程中,一些OB陷于骨基质中分化为骨细胞,而基础研究证实自噬可以促进OB的增殖、分化。Ozeki等[18]的研究表明ATG7和wingless/int1(Wnt)通路参与了骨形态生成蛋白-2(bone morphogenetic Protein⁃2,BMP⁃2)诱导的OB早期生成过程,BMP⁃2促进了ATG7的表达,但对LC3、Be⁃clin1和ATG5等无作用。通过小干扰RNA沉默ATG7和Wnt16后,显著抑制了BMP⁃2介导的人骨骼肌干细胞(hu⁃man skeletal muscle stem cell,hSMSC)向成骨细胞分化;而自噬激动剂雷帕霉素(rapamycin)提高了hSMSC的成骨分化,这说明自噬参与促进早期成骨细胞分化。这与Nollet等[19]关于OB在骨稳态和骨骼矿化过程中的研究结果一致:大鼠成骨细胞系的矿化能力随自噬相关基因ATG7和Beclin⁃1被siRNA技术沉默而显著下降。此外,Jung等[20]和Zhu等[21]的研究还表明增强自噬可以减轻细胞氧化损伤并抑制细胞凋亡:实验结果显示NO和H2O2等会引起OB的凋亡及功能异常,而通过AMPK通路活化自噬可以减轻相应的损伤。进一步,王雅雯等[22]通过CoCl2制备人hFOB1.19成骨细胞系的缺氧模型,结果显示缺氧环境可激活细胞自噬,并且OB的自噬水平与缺氧作用时间呈正相关,成骨细胞自噬活性的不断增强,对细胞起到一定的保护作用。
与此同时,也有部分研究表明自噬对OB起到负调控作用。Chen等[23]的研究中,在颅骨来源的OB中敲除Raptor(mTORC1的一个重要组成部分)以降低OB相应的mTORC1功能,提高细胞自噬水平,导致OB的基质合成与矿化能力均下降;相应的体内实验中,与对照组相比,Raptor敲除小鼠骨质减少。Yeong等[24]研究表明,在微重力引发的骨量丧失或者骨质疏松中,应用褪黑素可以降低MC3T3⁃E1中的自噬水平、抑制细胞的内质网应激和氧化损伤,从而提高细胞的存活率,其机制与磷酸化ERK/Akt/mTOR通路相关。
(三)自噬在OC中的作用
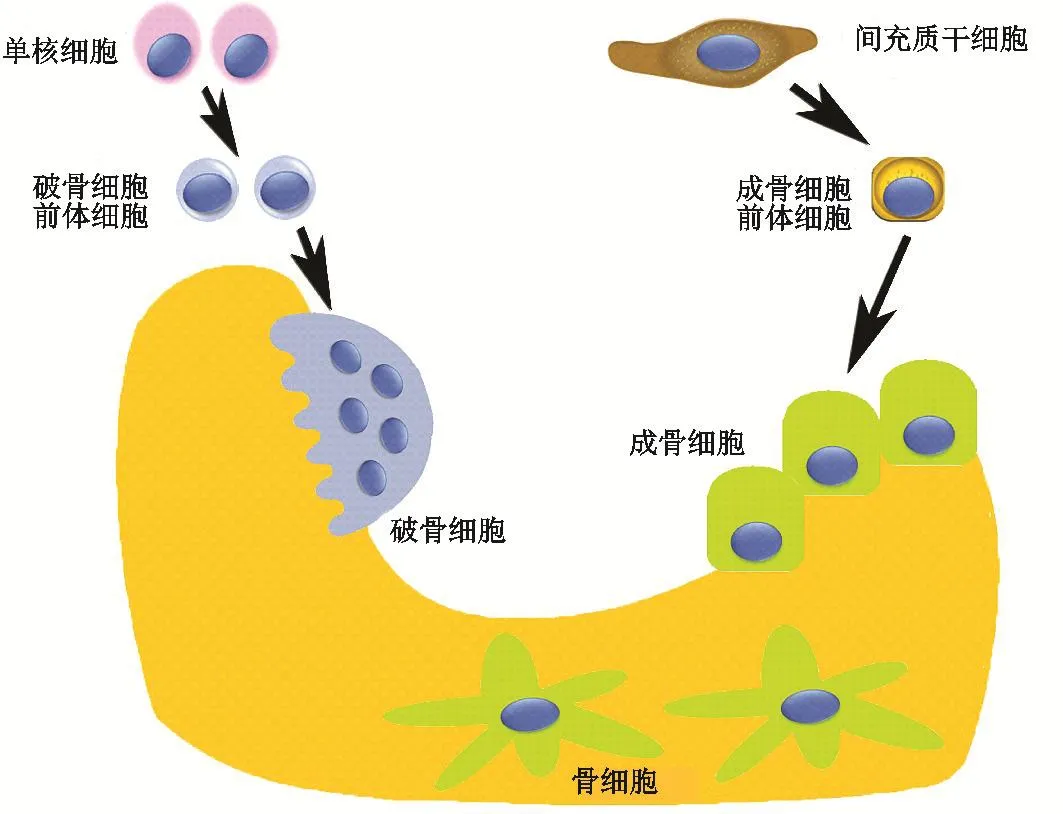
图3 各种细胞在骨稳态维持中的作用(绘图作者:张欣然)
OC是一种来源于骨髓的高度分化的多核巨细胞,OC不仅参与骨质的吸收,还是一种重要的免疫细胞。OC与OB的成骨作用相互协调配合,相辅相成,同时也参与免疫应答过程[25]。研究表明OC的形态可能与自噬密切相关,例如ATG5、ATG7、ATG4B、LC3等自噬相关基因的缺陷会引起OC形态变化、功能障碍[26]。Lin等[27]也发现在卵巢切除术或应用糖皮质激素的小鼠中,采取自噬抑制剂氯喹或抑制单核细胞ATG7可以抑制破骨相关基因的表达和OC的分化,从而缓解模型所诱发的骨质流失,印证了细胞自噬可能具有促进OC分化的作用。然而也有一部分研究得出相反的结论,指出细胞自噬可能抑制OC分化,例如在体外实验中使用自噬激活剂Rapamycin可以减少OC的形成,从而减少骨质流失[28]。
四、自噬与骨质疏松
在正常生理情况下,破骨-成骨的动态平衡使骨代谢保持正常状态,当这种平衡被打破时,就可能导致骨稳态的异常,最终发生OP。对于老年或绝经后妇女OP病人,往往并发OF等多种并发症[29]。有研究发现,与正常个体相比,体外培养OP来源的BMSCs,可见OP来源的BMSCs向成骨细胞分化的数目减少,更倾向于向脂肪细胞分化[30]。对OP病人外周血检测结果显示,与正常人相比,OP病人ALP、mTOR以及Runx相关转录因子2(runt⁃related transcription factor 2,Runx⁃2)的表达量均降低,相关性分析结果显示Runx⁃2与mTOR的表达呈正相关,并且骨密度与Runx⁃2的表达也存在正相关,提示OP病人骨密度的下降可能与自噬相关基因mTOR存在着一定关联性[31]。
关于细胞自噬和OP的相关性研究,目前已经在细胞和动物实验中广泛开展。研究发现老年大鼠的骨细胞自噬关键因子LC3⁃Ⅱ和Beclin⁃1表达降低、细胞自噬水平降低,而sequestosome⁃1(p62/SQSTM1)和骨细胞凋亡水平升高[32]。由于OP表现出一定的年龄相关性,因此我们推测不仅自噬与OP可能存在相互调控的关系,年龄因素、自噬与OP三者的相互关系同样不容忽视。除了年龄相关性骨质疏松症,自噬在遗传性骨质疏松症中同样有着重要的作用。常染色体隐形遗传性骨质疏松症(autosomal recessive osteopetrosis,ARO)是一种恶性的先天性骨质疏松症,研究表明在ARO模型小鼠中,调控细胞自噬早期阶段的V⁃ATP酶的编码基因发生突变,导致p62表达降低、LC3⁃Ⅱ水平升高,使自噬水平异常,从而影响了OC的发育成熟[33]。此外,OP也与雌激素水平密切相关,研究发现在小鼠卵巢切除模型中,雌激素水平的降低提高了自噬活性,小鼠胫骨近端骨矿物质密度下降;然而雌激素的应用能够促进自噬水平恢复正常,并抑制了骨的流失[34]。但是通过建立双侧卵巢切除大鼠骨质疏松模型,Flor⁃enci等[35]提出了相反的观点:雌激素缺乏可以导致自噬水平降低、细胞凋亡增多,而雌激素替代治疗可以逆转这一趋势。综上,近年来相关研究表明雌激素可以抑制骨细胞凋亡,自噬可能参与了这个过程,但是具体的正负调控作用仍不明确。此外,骨质疏松大鼠接受药物干预后,大鼠成骨标志物BMP⁃2、Runx⁃2和自噬标志物 Beclin⁃1、ATG5和LC3的表达均显著增加,微计算机断层扫描(micro computed tomog⁃raphy,micro⁃CT)检测发现大鼠脊椎骨的骨密度、骨量、骨小梁的数量及厚度均显著增加,这表明骨质疏松的改善与自噬水平的增加有关[36](图3)。
迄今,关于自噬与OP的关系的研究多聚焦于动物模型与离体实验,临床研究所提供的信息较少。Zhang等[37]的一项基于984位白人研究对象的人类基因研究表明,自噬通路调控基因是唯一与桡骨远端骨密度变化直接相关的基因,这为自噬在OP发生发展中的作用提供了直接的人体试验证据。
五、展望
细胞自噬在骨稳态的维持中起着重要的作用,骨稳态平衡的打破会导致OP的发生,然而自噬与骨稳态的调控及OP的发生、发展究竟为正相关还是负相关,是自噬在骨代谢领域中的研究焦点之一。一些研究提示提高细胞自噬水平可以促进破骨作用和骨吸收,自噬与OP呈正相关,因而采取抑制细胞自噬可能成为治疗或预防OP的新方法。然而,亦有一部分研究得出了相反的结论,指出自噬与OP呈负相关,对OP的发生发展有抑制作用,因此自噬水平的降低可能导致OP的发生。我们推测,过度或低下的自噬水平均可能不利于骨代谢和骨稳态,调控自噬水平使之保持在适度范围内可能成为治疗OP的新靶点,而这需要在进一步研究中加以阐释。此外,对于OP的全身性治疗中,近年来基础研究表明一些中药及其主要成分在治疗OP中具有一定的作用和价值,而自噬在中药干预OP中的作用和机制尚不明确,因此明确自噬与中药的关系能够为OP的治疗提供新靶点,为发挥中医药的特色诊疗提供新方法;由于OP发病率高,影响人群较大,对于OP病人局部骨缺损的修复也值得进一步研究,自噬是细胞成骨分化不可缺少的一个过程,骨缺损修复材料的改性是否会诱发成骨相关细胞自噬的改变从而来影响OP病人局部骨缺损修复的效果,也值得进行深入研究从而为指导骨修复材料的改性提供理论支持。
综上所述,通过深入研究细胞自噬在OP中的作用及机制,并通过调控自噬促进骨代谢的稳态平衡,可以为全身性延缓或控制OP的发生与发展以及局部促进OP病人骨缺损的修复提供新的思路。
