黄土丘陵区典型草地演替中植物群落特征与土壤储水量关系
2019-03-26赵勇钢刘小芳王子龙赵世伟刘新春朱兴菲
李 轲, 赵勇钢, 刘小芳, 王子龙, 赵世伟, 刘新春, 朱兴菲
(1.山西师范大学 生命科学学院, 山西 临汾 041004; 2.西北农林科技大学 水土保持研究所,陕西 杨凌 712100; 3.西北农林科技大学 资源环境学院, 陕西 杨凌 712100)
水分作为干旱、半干旱地区生态系统重要的环境因子,是黄土高原地区植被恢复与重建的主要限制因素之一[1]。在草地植被恢复演替过程中,草地植被与土壤水分相互作用,相互影响。一方面草地植被通过水分利用和消耗以及改变降水分布等方面影响土壤水分,另一方面土壤水分也会对草地植物的个体生长和生存以及群落分布、组成和结构等方面产生影响。在干旱和半干旱地区,土壤水分与植物冠层覆盖、叶面积、蒸腾和群落组成等生理生态因子密切相关,是生态过程的基础,也是生态系统演替和服务功能发挥的重要驱动因素[2]。一些研究表明[3-4],草地植被的生物量、群落结构组成和物种多样性等生态学特征与土壤水分具有显著的相关关系,草地植被在恢复演替和退化过程中的土壤含水量在不同时间阶段及土壤剖面上呈现出明显的变化特征。因此,探明土壤水分和植物群落的动态变化特征及其相互关系,对于阐明植被—土壤的互馈机制具有重要意义。
黄土高原地区水土流失严重、生态环境脆弱,退耕还林(草)是该地区恢复植被、改善生态的重要举措。生态系统形成是植被与环境因子相互作用协同演变的结果[5]。目前的研究主要集中在土壤水分对土壤养分的影响、土壤碳储量和土壤微生物结构方面,而关于黄土高原典型草原区植被群落演替过程中的植被生产力和多样性与土壤水分的关系报道较少,并且缺乏长时间序列的阶段性比较研究。因此,本研究拟基于属黄土高原半干旱区典型草原生态系统的云雾山国家级自然保护区长时间的恢复演替序列的研究样地[6],分析草地植被演替过程中植被生产力、植物多样性等生态学特征与土壤水分之间的相互关系,以期为黄土高原地区退耕地植被恢复提供一定的科学依据。
1 材料与方法
1.1 研究区概况
研究区位于宁夏回族自治区固原市东北部云雾山国家级自然保护区(106°24′—106°28′E, 36°13′—36°19′N),是中国西北黄土高原地区以本氏针茅(Stipabungeana)为优势种的典型地段,也是该地区唯一保存下来的天然草地[7]。保护区总面积7 000 hm2,海拔1 800~2 100 m,坡度15°~40°,属黄土高原长梁缓坡丘陵区。土层深厚,土壤类型为山地灰褐土和黑垆土。气候属中温带半干旱气候区,年均气温5 ℃,年均降水量400~450 mm(1983—2005年),其中7—9月降水量占全年的65%~75%,蒸发量1 330~1 640 mm,年日照时数2 500 h,无霜期112~140 d。保护区主要草本植物有本氏针茅、铁杆蒿(Artemisiavestita)、百里香(Thymusmongolicus)和厚穗冰草(Aneurolepidiumdasystachys)等,其中丛生禾本科植物本氏针茅分布最广[8-9]。
1.2 样品采集与测定方法
研究采用植被空间序列代替时间序列的方法进行调查取样分析[9]。1982年建立保护区时,建立了几个不同植被演替阶段的长期定点观测样方[10],加之保护区在不同年限土地退耕恢复形成的群落样地,已形成一个从演替初期到本氏针茅群落的长时间序列样地。
野外调查采样于2016年8月进行,在研究区选择有代表性的具有不同恢复年限的典型样地6个,包括恢复3 a(R3)、恢复8 a(R8)、恢复13 a(R13)、恢复46 a(百里香群落,R46)、恢复66 a(铁杆蒿群落,R66)和恢复89 a(本氏针茅群落,R89)。每个样地沿坡面水平设置3个10×50 m的样带,每个样带选取3个1 m×1 m样方,每个样方间距不低于10 m,每个样地共调查9个样方,共计54个样方。调查每个样方内所有草本植物的物种数、总盖度、高度和株数等。用收获法收集植被地上部分以及地表凋落物,带回室内在65 ℃下烘干后称量干重,分别计算地上生物量和凋落物生物量。由于研究区草地根系主要分布在0—1 m土层内[11],因此本研究把0—1 m土层内的根系生物量作为草地总地下生物量。在样方内用直径为9 cm的根钻按照10 cm进行分层取样,置于孔径为0.4 mm纱袋内带回室内漂洗干净,在65 ℃下烘至恒重后称量干重,计算地下生物量。每个样方挖1 m深的土壤剖面,去除枯枝落叶层后,按照0—5,5—10,10—20,20—40,40—70,70—100 cm土壤层次分别采集混合样、原状土样和环刀样,以测定土壤基本理化性质。土壤容重采用环刀法测定。用直径40 mm的土钻采集相应层次的土样装入铝盒,称取土壤鲜重后放入105℃烘箱烘12 h至恒重,计算土壤含水量。
土壤储水量的计算采用如下公式:
W=0.1×h×ρ×θ
(1)
式中:W——土壤储水量(mm);h——土层厚度(cm);ρ——土壤容重(g/cm3);θ——土壤含水量(质量含水量;%)。下同。
群落多样性特征用以下指标表示:
物种丰富度指数:
R=S
(2)
式中:S——物种数目。下同。
物种多样性用Shannon-Wiener指数表示:
(3)
式中:H——物种多样性指数;Pi——属于种i个体在全部个体中的比例。下同。
物种均匀度指数:
J=H/lnS
(4)
1.3 数据处理
各样方的根系生物量换算成单位面积的地下生物量,在此基础上取平均值,得到每个样地的地下生物量。总生物量=地上生物量+地下生物量+凋落物质量。对土壤含水量与生物量和多样性指标进行Pearson相关性分析,各变量之间的显著性差异及多重比较使用最小显著性差异法(LSD方法)。在SPSS 17.0软件上进行相关统计处理,采用Origin 9.0进行图形处理。
2 结果与分析
2.1 群落生物量和多样性指标
方差分析结果表明,在群落结构水平上,草地的盖度存在显著的差异(表1)。在生产力水平上,草地的地上生物量、凋落物质量、地下生物量、总生物量和根冠比存在显著的差异。在物种多样性水平上,各样地间群落的物种多样性、丰富度和均匀度有显著差异。
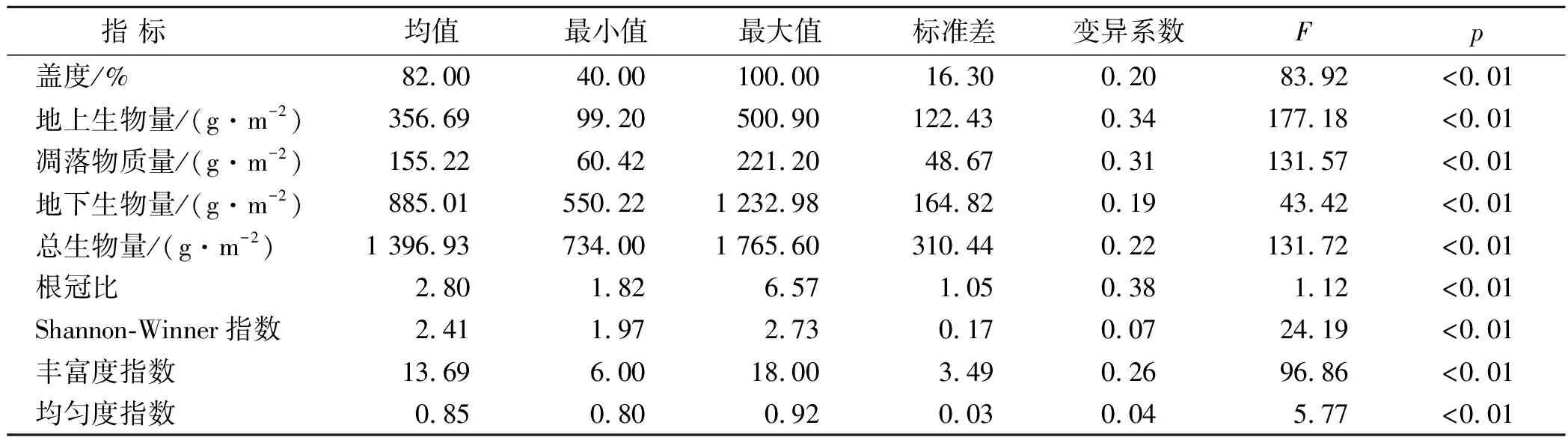
表1 黄土丘陵区草地群落演替植物生物量与多样性指标统计分析(n=54)
2.2 群落演替中生物量和物种多样性变化
随着草地植被演替的进行,群落盖度和生物量随着恢复年限的增加呈现增加趋势(表2)。退耕3 a群落盖度、地上生物量、凋落物生物量和总生物量显著低于其他5个植物群落(p<0.05)。草地恢复13 a后群落总生物量无明显差异(p>0.05),而根冠比显著高于其他5个群落(p<0.05)。从同一群落生物量的分配比来看,地下生物量占总生物量的58%以上。丰富度指数、Shannon-Wiener指数随着演替的进行变化趋势呈现为增加-减小-增大的趋势,最小值出现在恢复3 a,丰富度指数最大值出现在恢复13 a,Shannon-Wiener指数最大值出现在恢复46 a。均匀度指数则呈现减小的趋势,最大值出现在恢复3 a,最小值出现在恢复89 a。
总体来看,随着群落演替的进行,除均匀度外的其他指标在退耕13 a之前大多变化显著,之后逐渐趋于稳定,恢复89 a的本氏针茅群落稳定性达到最好,基本处于顶级群落。
2.3 群落演替中土壤水分变化
草地植被演替对0—40 cm土层的含水量有显著影响(p<0.05),但对40 cm以下土层的含水量没有显著(p>0.05)影响,其值在10%左右(图1)。随着植被演替的进行,土壤含水量呈现增加的趋势。在0—10 cm土层恢复46 a和89年的含水量最高,显著(p<0.05)高于恢复3 a样地。在10—40 cm土层恢复89 a的含水量最高,显著(p<0.05)高于恢复8 a和恢复3 a样地。随着土层的加深,土层含水量有逐渐增加的趋势。

表2 黄土丘陵区不同恢复年限样地的草地生物量与多样性指标
注:同列不同小写字母表示差异显著(p<0.05)。

注:图中误差棒为标准误。图1 黄土丘陵区草地演替过程中土壤含水量在土壤剖面的分布
由表3可知,同一土层中,植被演替过程对0—40 cm土层土壤储水量没有显著(p>0.05)影响,但对40—70,70—100 cm土层具有显著(p<0.05)影响,恢复89 a相比较恢复46 a分别降低了9.96%和7.56%。
说明植被演替过程对表层土壤储水量影响不大,但对深层已产生明显影响,尤其在演替后期土壤储水量有显著下降。同一个演替群落中,土壤储水量随着土层深度的增加而增大,土壤储水量在0—10 cm土层最小,在10—40 cm土层有显著的增加,在70—100 cm土层达到最大值。

表3 黄土丘陵区不同草地植被演替阶段的土壤储水量分布 mm
注:同行不同小写字母表示相同土层不同恢复年限土壤储水量的差异显著(p<0.05)。
2.4 土壤储水量与植被群落指标的相关性
由表4可以看出,土壤储水量与均匀度指数和Shannon-Winner指数在所有土层均无显著的相关关系(p>0.05),在0—5 cm与丰富度指数显著相关(p<0.05)。在0—10 cm与地下根系生物量和总生物量显著相关,在5—10 cm与地上部生物量有显著相关性。20—40 cm土层储水量与地上部植物生物量和地下根系生物量显著相关,40—70 cm土层与地上部植物生物量显著相关,而10—20,70—100 cm各土层储水量与所有因素均没有显著相关性。在0—10 cm土层储水量与地上生物量、地下生物量、总生物量显著相关(p<0.05)。地下生物量与0—40 cm土层土壤储水量显著相关(p<0.05),根冠比与0—70 cm土层显著相关。
3 讨 论
本研究中随着草地演替的进行,群落的盖度及生物量在恢复13 a前增加迅速,之后逐渐变缓;物种多样性呈波动式增加,这与李中林等[12]和郝红敏等[13]的研究结果相似。原因可能是草地演替过程中,初期一年生的草本植物逐渐被多年生禾本科植物所竞争淘汰,形成相对较稳定的群落结构。此外,植物群落的形成除了受植物个体相互间的竞争影响外,还可能受生境、干扰程度及遗传特性等的影响。

表4 土壤储水量与植物群落指标间的相关性(n=54)
注:*和**分别表示在0.05,0.01水平上显著相关。
本研究发现,土壤储水量不仅影响植物生物量,还影响地下生物量,这一观察结果与前人的研究结果一致[14]。同时,研究还发现,0—10 cm土壤储水量与生物量显著相关,这与张永旺[15]的研究结果一致。物种丰富度是群落的一个基本特征,在调节生态系统的结构和功能方面发挥着重要作用[16]。本研究结果表明,黄土高原的干旱和半干旱地区的表层土壤水分与植物丰富度之间呈正相关,与Deng等[2]人的研究结果一致。这是由于草地恢复过程中,主要优势物种由一年生草本植物逐渐变成多年生草本植物,根系生物量逐渐增大,根系生长也由浅层向深层进行,根系的更多耗水需要更多的土壤水分以维持植物自身生长[17],这也进一步说明土壤储水能力的提升对群落生产力及多样性的重要性。土壤水分在半干旱草原生态系统中调节植物多样性方面起着重要作用,一般来说,植物—土壤反馈效应可以改变植物群落结构和土壤性质[18]。
草地演替对0—40 cm土层容重和含水量影响较大,对40—100 cm影响较小(图1)。随着草地演替的进行,土壤含水量呈增加趋势,土壤储水量呈减小趋势,这与前人的研究结果一致[9,17]。董杰等[19]和苏永中等[20]对退化草地围栏封育恢复的研究表明,随着围封年限的增加,有机碳含量显著增加,土壤容重逐渐下降,土壤结构得到改善。在草地植被恢复过程中,植物群落结构与组成发生变化[21],枯枝落叶返还及根系周转会促进土壤有机质含量增加,促使土壤大团聚体形成[9],改变土壤结构[20,22-23],从而影响土壤持水和供水能力[9]。同时,植被群落的变化,生物量的增加会影响土壤水分的利用和消耗[24]。因此,土壤含水量和储水量的变化是植被与土壤相互作用的结果,是植被耗水与土壤保水和供水能力的平衡。本研究草地演替过程中,植被生物量、多样性均呈增加趋势,而0—40 cm土壤储水量没有显著变化(表3),这表明在植被演替过程中,土壤供水能力或者土壤水分状况没有因此减弱,这是植被能够继续演替的重要条件。但草地演替对40 cm以下土层土壤储水量有明显影响(表3),地下根系生物量的增加致使其对深层土壤水分的消耗增加可能是主要原因。
4 结 论
(1) 随着草地植被恢复年限增加,尤其在恢复13 a之前,植被群落生物量、多样性、盖度增加显著,之后增加趋于平缓。
(2) 随着演替的进行,土壤含水量有增加的趋势。植被群落演替对0—40 cm土层土壤储水量没有显著影响,说明土壤储水量能够支撑群落演替,但演替后期植物群落已明显表现出对40 cm以下土层水分的消耗。
(3) 0—10 cm土层水分变化与植被群落生物量及物种多样性指标有显著的正相关关系。
