饥饿对中华倒刺鲃幼鱼代谢、个性和集群的影响
2019-03-22唐金玉覃英莲曾令清彭姜岚付世建
王 蕾,唐金玉,覃英莲,曾令清,彭姜岚,付世建
重庆师范大学进化生理与行为学实验室 重庆市动物生物学重点实验室, 重庆 401331
季节更替、气候变化和人为活动等因素导致自然界水体食物丰度发生波动或时空分布不均,使得鱼类经常面临食物匮乏甚至饥饿风险[1- 4]。因此,鱼类在长期进化过程中形成了一系列的生理生化和行为策略来应对食物丰度的变动,如增加觅食空间、减少日常活动和降低能量代谢等[5]。标准代谢率(Standard metabolic rate, SMR)是指变温动物在非活动、无食物消化吸收状态的最小机体能量消耗强度,通常以耗氧率表征SMR[6- 9]。研究发现在种内的不同个体间存在稳定的SMR个体差异[10],且这种差异在经体重、年龄、性别等校正后仍可达2—3倍[11-12]。SMR是动物生活史理论重要的能量代谢参数,某些种类的SMR可占个体能量总支出的50%[13]。由于个体在整个生活史中从外界获取的总能量有限,SMR所占比例直接影响机体其他生理功能(如生长和繁殖)的能量分配,进而对个体乃至种群的生活史特征和适合度产生影响[11,14]。因此,食物丰度的改变会显著影响鱼类的SMR[15-16]。
稳定个体差异(Consistent individual difference, CID)普遍存在于自然界中,它是指动物表型(如生理和行为等)在种内水平上表现出来的个体差异,并且这种差异在一段时间内保持稳定[17]。研究者把动物行为的CID称为动物“个性”,通常用勇敢性、探索性和活跃性等指标来衡量。勇敢性是指动物对潜在风险的反应,是评价动物为获取更多食物或繁殖机会而愿意承担风险的个性行为[18]。探索性是指动物在面临新异环境时收集信息并适应该环境的自适应过程[19-20],主要体现在面临新环境时表现出来的离开隐蔽场所以及通过视觉或嗅觉搜集隐蔽场所之外的信息等个性行为[21]。活跃性是指动物不受外部环境刺激仅仅由于自身状态所引起的行为活动[22-23],主要采用自发运动作为指标,与机体觅食、寻找配偶、避敌等行为密切相关[24]。研究发现,鱼类的个性行为常常受到遗传因素、环境条件以及其生活史阶段特征的影响,且与机体的SMR密切关联[25-26];其中,饥饿胁迫导致的个性行为的变化在花鳉(Brachyraphisepiscopi)[27]和欧鲈(Dicentrarchuslabrax)[28]等不同鱼类均得到证实。这些研究发现饥饿过程中鱼类个性行为的变化可能存在种间差异,而特定种类个性行为在食物短缺条件下的适应性调整是否与能量代谢特征相关值得研究者关注[29]。
鱼类成群聚集、协调一致的集体行动被称为集群行为[30],大多数鱼类在其生活史的特定时期甚至整个生活史周期呈现集群行为[31]。鱼类集群行为有降低被捕食风险、增加觅食成功率和改善游泳效率等益处[32];但同时也会加剧食物和配偶的资源竞争[33]。现有研究通常关注个体行为及其交互作用如何影响鱼群的群体特征,其中最主要的群体特征为凝聚力和协调性。凝聚力通常以群体的个体间距离和最近邻距离等来衡量,而协调性主要通过不同个体之间游泳速度的同步性、群体极性、空间分布等来衡量[34-35]。研究发现,鱼类的集群行为与机体营养状况密切关联,可能显著影响鱼类的日常能量消耗[17]。
本研究的目的是考察食物资源短缺下暖水性鲤科鱼类能量代谢、个性与集群行为的应对策略及其可能的内在关联,即三者之间的变化是否存在关联。本研究以喜好流水生活的中华倒刺鲃(Spinibarbussinensis)幼鱼作为实验对象,测定饥饿前后实验鱼的SMR、个性行为(勇敢性、探索性、活跃性)以及集群行为(凝聚力和协调性)等能量代谢和行为学参数。中华倒刺鲃是一种喜好群体生活的鲤科鱼类,在野外或实验室条件下均呈现明显的集群行为[36]。
1 材料与方法
1.1 实验鱼来源及其驯化
中华倒刺鲃幼鱼(3—7 g)购于重庆本地渔场,于实验室自净化循环控温水槽(长1.2 m×宽0.55 m×高0.55 m,约250 L)中驯养两周。驯化水温为(25±0.5)℃,实验用水为曝气除氯自来水,溶氧水平维持接近饱和状态(≥7.0 mg/L),日换水量约为10%,驯化期间的光周期为14 L∶10 D。驯化期间,每日10:00AM用中国通威公司的浮性饲料饱足投喂实验鱼,投喂30 min后,用虹吸管清除剩余饵料和粪便。
1.2 实验设计
驯养结束后,称取鱼体健康、体重相近(5.25±0.16)的实验鱼200尾,使用体内PIT(Passive integrated transponder)方法进行电子标签体内标记以便区分实验鱼,随后进行两周的恢复。恢复结束后随机选取80尾鱼测定其体重、体长及SMR,之后随机均分为饥饿组和对照组(其中对照组1尾实验鱼实验测定过程出现技术故障,该数据未纳入分析),每组40尾,饥饿组实验鱼进行2周饥饿处理,对照组实验鱼每天以维持日粮(根据预实验得到)水平投喂一次。与此同时测定剩余120尾鱼的体重、体长及个性行为(勇敢性、探索性和活跃性)。测定结束后,也将实验鱼随机均分为饥饿组和对照组。两周处理之后,分别测定80尾实验鱼的SMR和120尾实验鱼的个性行为,同时测定实验鱼的体重和体长。最后选取测定SMR的80尾实验鱼,分别以4尾鱼为单位组成饥饿组和对照组鱼群(每处理组n=10),通过视频拍摄的方式记录饥饿和对照鱼群的集群行为。实验过程中溶氧、水温、光照等条件与驯化期间保持一致。
1.3 实验参数的测定
1.3.1 SMR的测定
本研究SMR测定仪器为实验室自制流水式呼吸代谢仪,仪器的原理结构详见已发表论文[37]。将禁食24 h并测定体重的实验鱼放入代谢仪呼吸室,再进行24 h适应。随后采用溶氧仪(HQ30d,美国哈希公司)每小时测定一次每尾实验鱼的耗氧率,每天测定12个点(08:00—20:00),连续测定两天。得到24个耗氧值,以其平均值作为对应实验鱼的SMR。实验鱼SMR(mgO2/h)的计算公式如下:
SMR=ΔO2×v
式中,ΔO2是实验鱼所在呼吸室出水口与空白对照溶氧的差值(mg/L),v是呼吸室的流速(L/h)。
1.3.2 个性行为的测定

图1 鱼类勇敢性、探索性和自发运动测定装置平面图 Fig.1 The structure of the device used to determine boldness, exploration and activity in fish speciesr:遮蔽区, refuge; d:可拆卸隔板小门, sliding door; oz:开阔区, open zone; ct:连接, connecting tube
个性指标采用实验室自制的鱼类行为测定装置(图1)测定,该装置主体为非透明有机玻璃制作的方形水槽(长70 cm×宽35 cm×高35 cm)。用有机玻璃隔板将玻璃缸分为大小不等两个区域,即开阔区(Open zone:长55 cm×宽35 cm×高35 cm)和遮蔽区(Refuge:长15 cm×宽35 cm×高35 cm)。有机玻璃隔板底部中央有一正方形开口(10 cm×10 cm)连通遮蔽区和开阔区,并附带有可拆卸的隔板小门(Sliding door)。开阔区正上方安置与计算机连接的高清摄像头(Logitech Webcam C930e)。拍摄时测定装置加入曝气自来水,水深12 cm,水温控制在(25±0.5)℃。
行为测定时首先将实验鱼放置在遮蔽区适应30 min,随后轻轻提起隔板小门,开始持续20 min的视频拍摄(15帧s-1),通过实验鱼在遮蔽区和开阔区的行为进行勇敢性分析。待勇敢性观测结束后,将实验鱼引入遮蔽区,闭合小门,放置一个内径为4 cm蓝色的PVC直角连接管于开阔区远离遮蔽区一端(该陌生物体距离边缘15 cm中心处,开口背向遮蔽区)。待实验鱼恢复20 min后,再次提起小门并开始20 min 的视频拍摄,通过实验鱼对陌生物体的侦查行为进行探索性分析。探索性观测结束后,立即取出陌生物体,确保实验鱼处于开阔区并闭合小门,待其恢复10 min后进行10 min的自发游泳行为的拍摄,用于活跃性分析。
勇敢性参数采用游出遮蔽区的次数和在开阔区停留时间(min),由人工观测计数得到结果;探索性参数采用距离陌生物体的平均距离(cm)和进入陌生物体的次数。大部分实验鱼在20 min的观测期内会离开遮蔽区接近陌生物体在进行探索,其中37尾实验鱼观测过程一直停留于遮蔽区内,无法进行侦查行为的评估。活跃性参数采用实验鱼在开阔区移动的总距离(m)和移动时间(min)。探索性和活跃性视频皆由Noldus动物运动轨迹跟踪软件(EthoVision XT 9.0, Noldus, Netherlands)软件分析;其中,探索性测定过程中,若实验鱼未游出遮蔽区(r)进入开阔区探索则不计入统计。活跃性测定过程中5尾鱼视频出现技术故障无法进行视频解析未计入结果。
实验鱼的个性指标均在第二天进行重复测定,以2次测定的平均值进行后续的统计分析[29]。
1.3.3 群体行为的测定
已有研究表明鲤科鱼类4尾鱼聚集时会出现典型的群体现象[38],在本研究集群行为测定过程中将以每4尾鱼作为一个群体。群体行为由直径50 cm的圆形鱼类集群行为测定装置测定,该装置上方放置连接计算机的摄像头。测定时从对照组或饥饿组随机选4尾实验鱼放入该装置适应10 min后,打开摄像头进行20 min的拍摄。拍摄视频(15帧/s)采用idTracker图像软件分析,获取每尾实验鱼每一帧的运动轨迹及其坐标点。随后根据坐标点进行实验鱼的速度和空间分布特征的计算。测定指标包括游泳速度(Vt, cm/s)、鱼体间的平均距离(Dt, cm)、最近邻距离(cm)和排列极性,其计算公式如下:
式中,Vt表示游泳速度(cm/s),xt和yt分别代表实验鱼在t时刻的横纵坐标点,dt为每两帧图像间的时间间隔。
式中,Dt表示鱼体间距离(cm),xt和yt分别代表实验鱼在t时刻的横纵坐标点。
1.4 数据统计和分析
实验参数用统计软件SPSS 17.0进行统计分析。实验前后个性行为和SMR的可重复性,即处理前后各指标的相关性均采用Pearson相关分析。以鱼的编号为随机因子,处理和测定时间为固定因子,分别对形态参数、SMR和个性指标进行线性混合模型进行分析;若存在显著差异,则分别采用独立和配对t检验分析实验组和测定时间的统计差异。实验数据用Excel进行常规计算,所有数据均以平均值±标准误(Mean±SE)表示,显著性水平为P<0.05。实验处理过程中个性行为饥饿组实验鱼丢失三尾,未纳入数据统计中。
2 结果
2.1 饥饿对实验鱼体长、体重的影响
饥饿组实验鱼在实验处理后体重显著下降8.1%(P<0.05),而体长无显著变化(表1)。对照组实验鱼实验处理后体长和体重均无显著变化。因此,实验结束后对照组实验鱼体重显著大于饥饿组(P<0.05)。
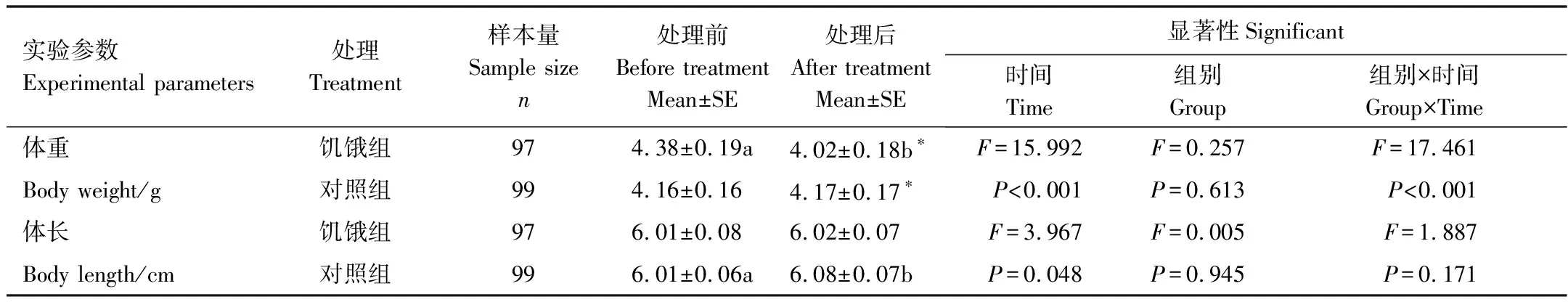
表1 饥饿胁迫对中华倒刺鲃体重和体长的影响
不同字母表示测定前后存在差异, *表示饥饿组和对照组的数据存在差异
2.2 饥饿对SMR的影响
饥饿组实验鱼的SMR处理前后存在重复性(P=0.004),而对照组SMR不具重复性(图2)。实验处理后,饥饿组实验鱼和对照组实验鱼个体耗氧率分别显著下降24.8%和13.1%(P<0.05)(图2)。

图2 不同实验处理组SMR的可重复性及饥饿对中华倒刺鲃SMR的影响(对照组N=40, 饥饿组 N=39)Fig.2 Repeatability of SMR in different experimental treatments and effects of starvation on the SMR of qingboSMR:标准代谢率, standard metabolic rate
2.3 饥饿对个性行为的影响
处理前饥饿组和对照组的各个性指标均无显著差异;经过两周实验处理,勇敢性参数游出次数和在开阔区的时间均显著增加(P<0.001),且实验处理后饥饿组实验鱼游出次数和开阔区时间显著大于对照组(表2,图3);经过两周实验处理,探索性参数距陌生物体平均距离显著缩短、进入陌生物体频次显著增加(P<0.001)(表2),且实验处理后饥饿组实验鱼进入陌生物体的频次显著高于对照组(P<0.05);经过两周实验处理,活跃性参数移动总距离和移动时间均显著增加(P<0.001)(表2),但饥饿组和对照组间无显著差异。

表2 饥饿对中华倒刺鲃勇敢性、探索性和活跃性的影响

图3 饥饿胁迫对中华倒刺鲃勇敢性、探索性和活跃性的影响(平均值±标准误,样本量见表2)Fig.3 Effect of starvation on the boldness, exploration and activity of the qingbo ((Mean±S.E.,see Table 2 for sample size)不同字母者表示测定前后存在差异,* 表示饥饿组和对照组的数据存在差异
2.4 饥饿对集群的影响
与对照组相比,饥饿组实验鱼个体间距离显著缩短9.4%(P<0.05)(图4)、平均速度显著上升11.3%(P<0.05)(图4),而对照组和饥饿组的最近邻距离和排列极性没有显著差异。
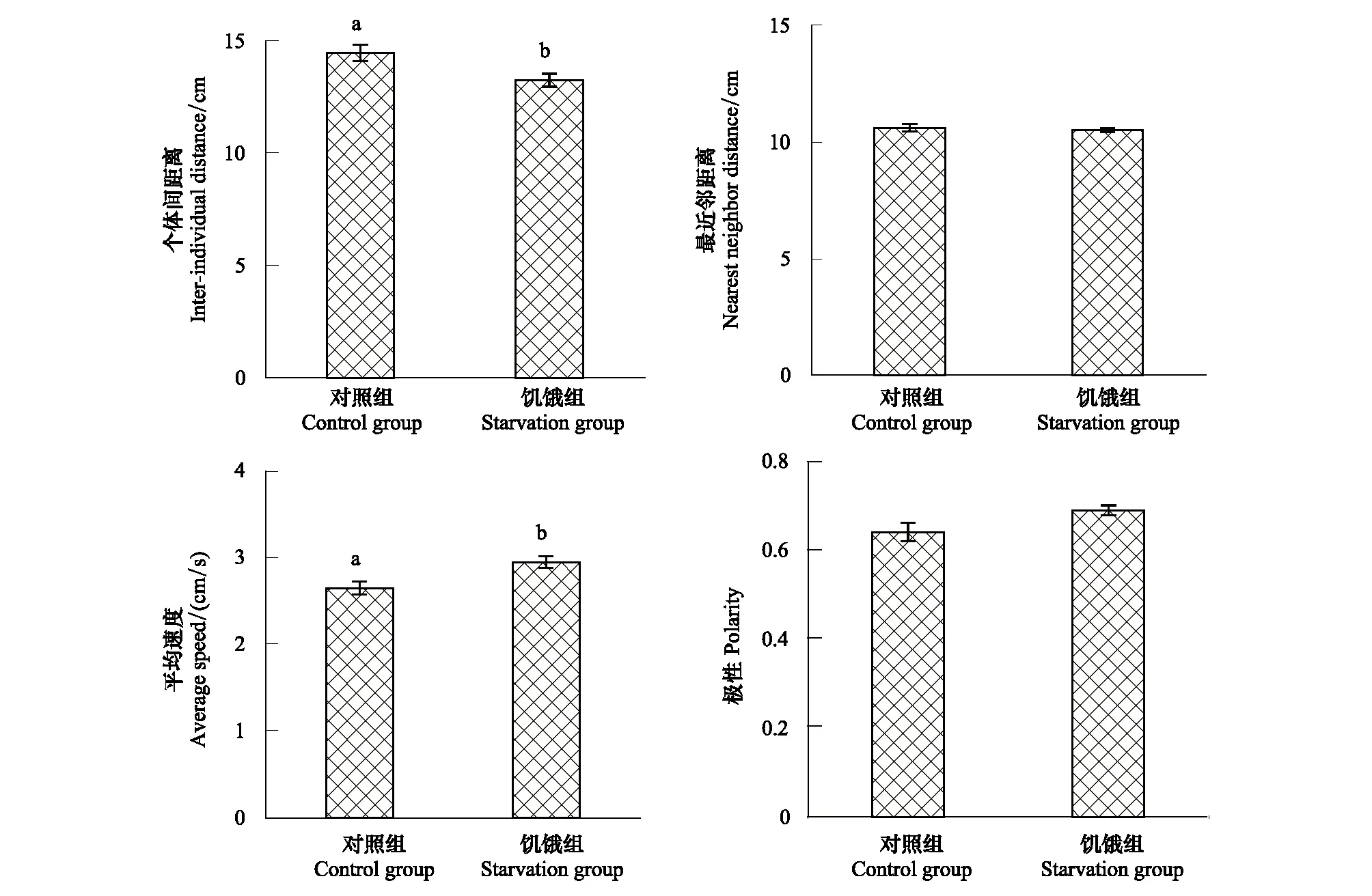
图4 饥饿胁迫经历对中华倒刺鲃群体行为的影响(平均值±标准误,N=10)Fig.4 Effects of starvation on the schooling behavior of qingbo (Mean±S.E., N=10)
3 讨论
3.1 饥饿对中华倒刺鲃SMR的影响
SMR是动物最为重要的生理功能参数之一,表征个体的维持能量消耗强度。长期饥饿状态下鱼类对其身体贮能的利用存在着两个相悖的适应需求:一方面通过降低代谢水平可以减少能量消耗,使得有限的身体贮能维持更长的生存时间;;另一方面尽可能将代谢保持在相对较高的水平,以保证在重新获得食物供应或面临其他环境胁迫时能产生适当的应激反应[39]。研究发现大多数鱼类,如南方鲇(Silurusmeridionalis)[40]、鲇(Silurusasotus)、褐鳟(Salmotrutta)[41]和银大麻哈鱼(Onchorhynchuskisutch)[2]等均会依据食物资源的丰度相应调节消化道功能,并逐渐改变代谢强度。本研究中对照组和饥饿组中华倒刺鲃SMR在实验过程中均出现了显著的下降,其中对照组SMR的下降可能是因为实验鱼在实验室单调驯养环境下的“囚禁效应”(captivity effect)所致[42],该现象在实验室以往的鲤科鱼类研究中也多次被发现[43]。可能与本实验采用的呼吸室体积狭小、饲养环境稳定单一,致使实验鱼生理功能下调等因素有关。与我们预期不同,经过2周的处理,饥饿处理组实验鱼的SMR和对照组没有显著差异,这可能是由于实验鱼偏小,实验过程的自发活动造成实验过程的实验误差偏大,掩盖了饥饿对SMR的影响。然而饥饿组实验鱼实验过程下降24.8%,远大于对照组的13.1%。这说明与其他鱼类一样,中华倒刺鲃在饥饿过程中也会相应下调SMR,以节约能量消耗。
一般认为,鱼类SMR的个体差异具有一定的稳定性,即在一段时间内可以重复。然而本研究发现对照组SMR试验前后没有重复性,这可能是实验误差所致。有趣的是,饥饿组实验鱼的SMR具有较高的重复性,可能因为饥饿条件下鱼类降低了不必要的生理活动,实验误差降低所致。
3.2 饥饿对中华倒刺鲃个性行为的影响
鱼类的个性受遗传、生活史、环境等因素的影响,具有较高的可塑性。当鱼类所受内外环境发生改变时,鱼类个性也将随之改变[44]。研究发现鱼类的个性特征主要与其经历的捕食强度与饥饿程度有关[23]。本研究发现,无论饥饿组还是对照组,实验结束时勇敢性、探索性和活跃性均显著上升。原因可能涉及上述提到的“囚禁效应”的影响;另外,这也可能说明实验前期的驯化时间偏短,正式实验前实验鱼尚未完全适应实验室条件,在首次实验测定过程中表现出较强的胁迫,抑制了鱼类的个性行为活动。
本研究发现,与对照组相比,饥饿处理后的中华倒刺鲃呈现出更高的勇敢性和更强的探索性。这可能是因为饥饿组实验鱼急需补充能量和营养物质,因此更为急切的获取食物资源信息。这和以往的研究结果相似,如有关鲫(Carassiusauratus)和欧鲈(Dicentrarchuslabrax)的研究均发现饥饿后个体表现出更高的勇敢性[27,45]。然而这种勇敢性和探索性的提高,可能导致被捕食风险的上升。因此,一些鱼类在饥饿后会表现出截然不同的个性变化,如饥饿导致狭鳕(Theragrachalcogramma)的勇敢性降低[46]和翡翠凤凰鱼(Pelvicachromistaeniatus)探索行为的减少[47]。这说明,鱼类在面临饥饿时个性变化因种类而异,可能存在不同的适应对策,而这种对策的差异可能与机体反捕食能力或环境的捕食压力高低有关。本研究中,中华倒刺鲃幼鱼面临饥饿选择提升自身勇敢性和探索性以期待寻找到更多的食物。一般认为,鱼类个性特征与SMR可能存在正相关,而本研究中实验鱼饥饿逆境下二者截然相反的变化趋势值得进一步研究。可能原因是饥饿个体生理功能下调诱导的代谢下降掩盖了个性行为增加导致的能量消耗的上升[29]。
有关梭鱼(Lizahaematocheila)[48]、牙鲆(Paralichthysolivaceus)[49]等研究发现经历饥饿的鱼类为降低能量消耗会减少自发活动;但有关鲤(Cyprinuscarpio)幼鱼[24]和斑马鱼(Daniorerio)[50]的研究显示,饥饿会增加的其活跃性,这可能与饥饿条件下觅食行为的增加有关。本研究发现饥饿后中华倒刺鲃活跃性没有明显的变化,这可能是觅食行为需求和节约能量需求相互抵消所致。
3.3 饥饿对中华倒刺鲃集群行为的影响
以往研究发现,饥饿会影响群体中个体的空间分布及其行为,这些行为的变化体现在个体和同类的相互作用,并且不同的鱼类可能采取不同的策略保持群体运动的协调性[51]。遭受饥饿胁迫的鱼群在空间分布过程中,通常会增加个体间距离,扩大搜寻水体空间并可以减少个体间的食物竞争,但这种集群行为的改变可能会增加鱼群边缘个体的被捕食风险[52];另有研究显示,拟鲤(Rutilusrutilus)个体间距离越大的群体能量消耗也越多[51],表明个体间距离的大小影响了的群体游泳效率。而本研究发现,饥饿组实验鱼个体间距离显著小于对照组实验鱼,这可能是因为饥饿组实验鱼在饥饿过程中消耗较多能量,为降低能量消耗,饥饿组实验鱼在群体行为中减少个体间距离,增强凝聚力。本研究还发现,饥饿组的群体速度和速度同步性显著高于对照组,即具有更好的协调性。这种现象在前期研究中也有类似报道,如食蚊鱼(Gambusiaholbrooki)[53]、大西洋鲱(Clupeaharengus)[54]等的研究均表明在饥饿环境中群体的空间分布及其行为调整策略可能有利于提高群体生存能力。
综上所述,饥饿导致中华倒刺鲃SMR有所下降,而饥饿后增强的勇敢性和探索性可能有利于提高个体的搜寻食物的成功率。另外,饥饿迫使中华倒刺鲃群体结构发生明显调整,其凝聚力和协调性的提高可能有助于群体生存能力。然而,中华倒刺鲃个性行为和集群结构的适应性变化是否由能量代谢介导的结果有待进一步研究。
