长江源区沙化高寒草地植被群落特征及其与地形因子的关系
2019-03-22屈建军徐先英唐进年韩庆杰谢胜波赖俊华
赵 鹏,屈建军,徐先英,唐进年,韩庆杰,谢胜波,王 涛,赖俊华
1 中国科学院西北生态环境资源研究院,沙漠与沙漠化重点实验室,兰州 730010 2 甘肃省治沙研究所,荒漠化与风沙灾害防治国家重点实验室培育基地,兰州 730070 3 中国科学院西北生态环境资源研究院,敦煌戈壁荒漠与环境研究站,敦煌 736200 4 中国科学院大学,北京 100049
青藏高原占我国国土面积的四分之一,平均海拔在4000 m以上,属于典型的高寒区。由于受地理位置、地形地貌、气候特征以及土壤类型等因素影响,青藏高原是我国乃至世界上高海拔地区生物多样性的重要区域之一。高寒草地占全国草地总面积的38%,地带性草地由东南向西北依次为高寒草甸—高寒草甸草原—高寒荒漠草原—高寒荒漠更替分布。受全球气候变化和人类活动影响,近几十年来青藏高原已成为沙漠化发生发展的重点区域[1- 3]。2010年风蚀退化土地面积374785 km2,占高原总面积的15%[4]。沙化造成高寒草地持续退化和生产力不断下降,严重威胁着青藏高原的生态安全,引起各方的高度关注[5- 6]。
沙化草地的土壤和植被退化相互作用,并具有负反馈效应[7]。随着沙漠化的发展,草地的植被盖度、高度、地上地下生物量和凋落物量急剧下降,物种丰富度、多样性指数、均匀度指数和植物密度呈波动式下降。多年生优良牧草在群落中的作用下降,一年生杂类草在群落中的作用增强,草地由多年生禾本科植物占优势的群落向一年生禾本科、藜科杂类草为优势种的群落演替[8]。土壤粗化、松散度增加、有机质和养分含量降低,生境异质性增大,灌丛化明显,群落结构趋于简单化[9]。气候、土壤一定的小尺度条件下,地形要素(坡度、坡向、坡位)通过对光照、积温、土壤水分、养分的再分配间接影响植物生长动态及物种分布[10-11]。草原区微地貌变化对生境异质性的控制作用明显[12]。沙丘形态造成其不同部位风沙活动及土壤水分、pH值和全盐含量的差异是抛物线形沙丘和白刺灌丛沙丘上植物群落相异的重要影响因素。沙丘固定导致丘间低地植物种间关系更加紧密[13]。沙质地表基质的流动性和风沙活动通过重塑微地貌形态进而影响着沙地植被的演替[14]。江河源区自然地理条件独特,以往针对沙化草地群落特征及其与微地形因子关系的研究尚未见报道。为此,本文以长江源区七渡口沙化高寒草地为研究对象,通过植物-土壤系统及地貌因子野外调查与室内分析,运用数量生态学方法,研究沙化草地的群落特征及其与微地形因子的关系,以期揭示沙化高寒草地演替规律及其地形驱动机制,为沙化草地恢复提供科学依据。
1 材料和方法
1.1 研究区概况
三江源生态保护与建设工程实施以来,区域生态环境发生了明显的改善,草地退化趋势基本得到控制。但黄河源和长江源草地退化形势依然比较严峻,其中玛多县、曲麻莱县等草地退化最明显[15]。曲麻莱县位于青海省西南部,是我国南北两大水系的主要水源涵养地。县域内平均海拔4500 m以上,年均气温-3.3℃,年均降水量380—470 mm,属高寒大陆性气候。曲麻莱县沙化危害严重,高寒草地退化明显,依托青海省林业厅沙化土地治理试验示范项目,2016年在七渡口高寒草地沙化区域进行了工程+生物综合恢复试验:依据试验区风沙活动规律,铺设HDPE沙障固定流沙,春季在沙障区播撒早熟禾(Poaannua)、老芒麦(Elymussibiricus)、披碱草(Elymusdahuricus),以期人工恢复沙化草地,遏制高寒草地沙化的趋势。试验示范区位于曲麻河乡昂拉村境内的长江通天河七渡口,距曲麻河乡政府所在地37 km,属于三江源国家级自然保护区索加-楚玛尔河野生动物核心区,草地类型以针茅草原和高寒草甸为主,优势种为高山嵩草(Kobresiapygmaea)、短花针茅(Stipabreviflora)、紫花针茅(Stipapurpurea)、青海固沙草(Orinuskokonorica),伴生种有二裂委陵菜(Potentillabifurca)、早熟禾、苔草(Carextristachya)、矮火绒草(Leontopodiumnanum)、黄花棘豆(Oxytropisochrocephala)[16]。地貌类型以冰川地貌、高山地貌、高原丘陵地貌为主,其间有强烈侵蚀中山地貌、高原低山丘陵地貌、河谷地貌。主要土壤类型有高山寒漠土、高山草甸土、高山草原土、山地草甸土、栗钙土、灰褐土、沼泽土、风沙土等类型,以高山草甸土分布最多。草原、草甸、灌丛等退化后形成的大面积裸地和“秃斑地”。土壤解冻期4月下旬,5月初,9月中下旬封冻,无冻期144 d。
1.2 试验设计
在沙化草地典型微地貌缓坡地、河漫滩、山麓地、沟谷地分别设置1个样地(表1),采用样线和样方相结合的方法进行植被群落调查,每个典型微地貌区样地设置3条重复样线,样线长度根据微地貌单元大小确定,每条样线隔5 m设置1 m×1 m样方,共调查60个样方,记录样方内植物的种类组成、高度、多度、盖度,各样线上取其重复样方相关参数的算术平均值进行计算分析。现场用罗盘记录样地坡度、坡向、坡位等微地貌因子,同时采用GPS全球定位系统测定样地的经纬度及海拔。

表1 调查样地基本情况
1.3 数据处理
在样带尺度上采用重要值(importance value, IV)作为各种植物在群落中的优势度指标,表示不同植物在群落中的功能地位,具体计算公式为:灌草重要值=(相对密度+相对频度+ 相对盖度)/3,各样带重要值分别为调查样方的均值。频度界定为样带内植物种出现的样方数/总样方数,仅出现于1个或2个样方的种界定为稀有种。参考丹麦学者C.Raunkiaer的研究方法绘制物种标准频度分布图,依次将标准频度在1%—20%的物种归入A级,21%—40%者归入B级,41%—60%者为C级,61%—80%者为D级,81%—100%者为E级[17]。
地形因子包括海拔、坡度、坡向和坡位,建立60×4的样方环境因子矩阵。坡位赋值方法为:1代表河谷平滩;2代表下坡位;3代表中坡位;4代表上坡位;坡向赋值方法为:1—8分别代表北坡、东北坡、东坡、东南坡、南坡、西南坡、西坡和西北坡。坡度直接用罗盘测定。
采用双向指示种分析(Two-way indicator species analysis,TWINSPAN)对沙化草地群落进行群丛类型划分,分类软件采用WinTWINS 2.3。DCA及CCA排序分析采用国际标准生态学软件Canoco 5,排序分析方法如下:先对物种重要值—样方数据进行除趋势对应分析(Detrend Correspondence Analysis,DCA),根据DCA排序轴的梯度长度选择适宜的排序方法。一般排序轴梯度长度< 3时,宜采用线性模型排序;梯度长度> 4时,宜采用单峰模型排序;梯度长度介于3和4之间,两种模型都适合。排序前对所有环境因子进行RDA筛选,若某环境因子的变异膨胀因子(Inflation factor)>20,表明与其他环境因子有多重共线性,对模型的贡献很小可以舍去。
2 结果
2.1 植物群落特征
60个调查样方中出现29个植物种,隶属16科,27属,其中以禾本科丰富度最高为5个种,菊科、玄参科各3个种,藜科、蓼科、十字花科、毛茛科、罂粟科各2个种,豆科、忍冬科、紫草科、唇形科、莎草科、蔷薇科、百合科、大戟科各1种。不同微地形沙化草地物种组成各异,整体上群落结构简单,根据物种重要值群落命名规则,缓坡地、河漫滩、山麓地、沟谷地群丛类型分别是赖草+绳虫实+披碱草,赖草+披碱草+细叶亚菊,赖草+披碱草+肉果草,赖草+早熟禾+绳虫实(表2)。赖草是不同地形沙化草地群落的优势种,缓坡地包含植物种最多为18,山麓地最少为14种。
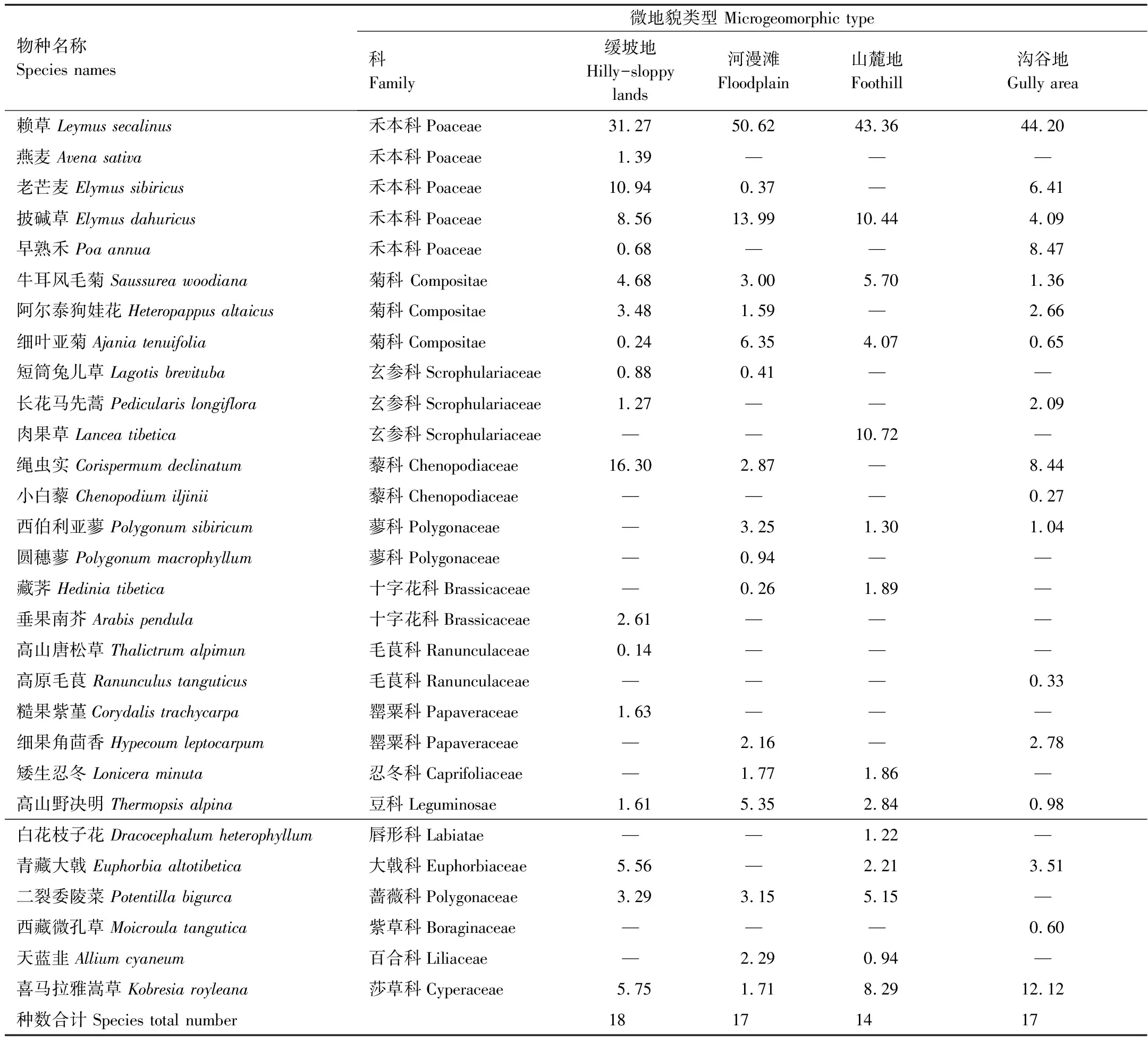
表2 不同地形沙化高寒草地物种重要值

图1 根据C.Raunkiaer方法绘制的物种标准频度分布图 Fig.1 The distribution of standard frequency of species referring to the Raunkiaer′s law参考丹麦学者C.Raunkiaer的研究方法绘制物种标准频度分布图,依次将标准频度在1%—20%的物种归入A级,21%—40%者归入B级,41%—60%者为C级,61%—80%者为D级,81%—100%者为E级
根据C.Raunkiaer方法绘制的物种标准频度分布见图1,缓坡地,各级别物种数表现为A>B>D,无E级物种,而E级植物通常是群落中的优势种和建群种,暗示群落缺乏优势种正处于高度演替阶段。河漫滩结果为 A>C>B=D=E,跟缓坡地相比,优势种比例增加。山麓地,A>B=C>E,优势种比例进一步增加。沟谷地,A>B>C=D=E,此结果与Raunkiaer的标准频度定律比较接近。从物种组成看,优势种比例低于稀有种。有2种植物出现在C、D、E级,即平均频度为40%以上,分别是赖草、披碱草。平均频度为20%—40%之间的B级植物有4种,分别是青藏大戟、喜马拉雅嵩草、绳虫实、细叶亚菊。稀有种植物有14种,占总物种数的48.23%。
2.2 植物群落数量分类
TWINSPAN 的划分结果如图2所示,可以看出各个植物种的观测值集中分布在矩阵对角线附近,生态型相似的植物种聚集分布。依据群落生境特征的指示种和优势种命名群丛类型,最终得到沙化高寒草甸植物群落7个群丛如下:

图2 沙化高寒草地群落的双向指示种(TWINSPAN) 分类结果Fig.2 Two-way indicator species analysis classification results of sandy alpine grassland communityPoa:早熟禾,Poa annua;Eld:披碱草,Elymus dahuricus;Les:赖草,Leymus secalinus;Els:老芒麦,Elymus sibiricus;Avs:燕麦,Avena sativa;Ajt:细叶亚菊,Ajania tenuifolia;Hea:阿尔泰狗娃花,Heteropappus altaicus;Cod:绳虫实,Corispermum declinatum;Chi:小白藜,Chenopodium iljinii;Pos:西伯利亚蓼,Polygonum sibiricum;Saw:牛耳风毛菊,Saussurea woodiana;Pom:圆穗蓼,Polygonum macrophyllum;Hyl:细果角茴香,Hypecoum leptocarpum;Arp:垂果南芥,Arabis pendula;Het:藏荠,Hedinia tibetica; Pob:二裂委陵菜,Potentilla bifurca;Eua:青藏大戟,Euphorbia altotibetica;Hin:矮生忍冬,Lonicera minuta; Alc:天蓝韭,Allium cyaneum;Tha:高山唐松草,Thalictrum alpinum; Kor:喜马拉雅嵩草,Kobresia royleana; Ths:高山野决明,Thermopsis alpina;Mot:西藏微孔草,Microula tibetica;Drh:白花枝子花,Dracocephalum heterophyllum;Pel:长花马先蒿,Pedicularis longiflora;Lat:肉果草,Lancea tibetica;Cot:糙果紫堇,Corydalis trachycarpa;Lab:短筒兔耳草,Lagotis brevituba;Rat:高原毛茛,Ranunculus tanguticus
群丛Ⅰ(计算机编码000)为赖草+阿尔泰狗娃花+牛耳风毛菊(Ass.Leymussecalinus+Heteropappusaltaicus+Saussureawoodiana),包括样方4、5、1、2、3,伴生种有青藏大戟、二裂委陵菜、早熟禾,分布在沙化缓坡地坡底,群丛盖度为3.94%。
群丛Ⅱ(计算机编码001)为赖草+喜马拉雅嵩草+绳虫实(Ass.Leymussecalinus+Kobresiaroyleana+Corispermumdeclinatum),包括样方49、57、11、48、50、53、6、7、8、9、42、51、10、52、43、44。伴生种有青藏大戟、二裂委陵菜、肉果草,分布在沙化缓坡地,群丛盖度为4.35%。
群丛Ⅲ(计算机编码010):赖草+披碱草+细叶亚菊(Ass.Leymussecalinus+Elymusdahuricus+Ajaniatenuifolia),包括样方31、46、24、25、26、45、60、23、32、33、30、41、47、21、35、36、37、38、54、28、29、34。伴生种绳虫实、天蓝韭、西伯利亚蓼、牛耳凤毛菊、喜马拉雅嵩草,主要分布在河漫滩沙地,群丛盖度6.83%。
群丛Ⅳ(计算机编码011)为赖草+高山野决明(Ass.Leymussecalinus+Thermopsisalpina),包括样方22、27、39、40,伴生种有绳虫实、长花马先蒿,主要分布在沙化缓坡地顶部,群丛盖度4%。
群丛Ⅴ(计算机编码100)为老芒麦+早熟禾(Ass.Elymussibiricus+Poaannua),包括样方55、56、58、59,伴生种有赖草、披碱草、绳虫实、喜马拉雅嵩草,主要分布在山麓地,群丛盖度2%。
群丛Ⅵ(计算机编码101)为老芒麦+披碱草(Ass.Elymussibiricus+Elymusdahuricus),包括样方16、17、18、20、19。伴生种有绳虫实、糙果紫堇、短筒兔耳草,主要分布在沙化严重的山麓地,群丛盖度3.23%。
群丛Ⅶ(计算机编码11)为绳虫实(Ass.Corispermumdeclinatum),包括样方12、13、14、15,主要分布在沙化沟谷地,群丛盖度2.12%。
2.3 植物群落排序
60个样方—重要值的DCA分析结果表明,4个排序轴的特征值分别为0.56、0.28、0.23、0.17,因其前两个排序轴的特征值较大,显示出重要的生态意义,所以采用前2个排序轴的数据分别作二维排序图(图3)。各群丛类型在排序图的分布呈现一定的规律性: 7个群丛由左到右依次变化,I、Ⅱ和VI、Ⅶ分布在排序图的两端,III和Ⅳ分布在排序图的中央。绳虫实是流动沙地常见的一年生草本植物,在沙地恢复过程中起着非常重要的作用。其群丛Ⅶ与其他群丛相距较远,代表沙化严重的生境。TWINSPAN分类所得各群丛类型与DCA排序结果基本吻合。
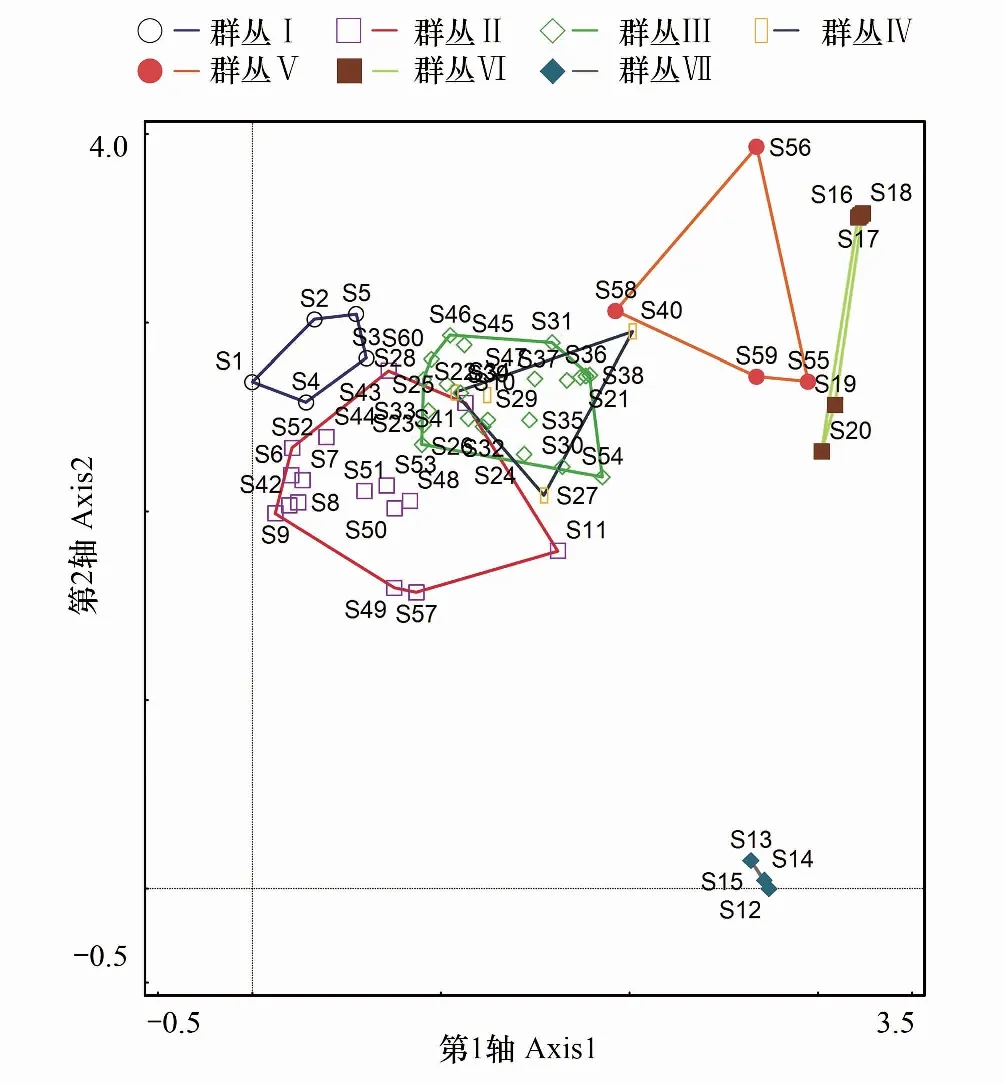
图3 沙化高寒草地群落样方的DCA排序图 Fig.3 DCA two-dimensional ordination diagram for samples of sandy alpine grassland community群丛I:赖草+阿尔泰狗娃花+牛耳风毛菊,Leymus secalinus+Heteropappus altaicus+ Saussurea woodiana;群丛II:赖草+喜马拉雅嵩草+绳虫实,Leymus secalinus+Kobresia royleana+Corispermum declinatum;群丛Ⅲ:赖草+披碱草+细叶亚菊,Leymus secalinus+Elymus dahuricus+Ajania tenuifolia;群丛Ⅳ:赖草+高山野决明,Leymus secalinus+Thermopsis alpina;群丛Ⅴ:老芒麦+早熟禾,Elymus sibiricus+Poa annua;群丛Ⅵ:老芒麦+披碱草,Elymus sibiricus+Elymus dahuricus;群丛VII:绳虫实,Corispermum declinatum;S1—S60代表样方编号
沙化高寒草地群落物种重要值—样方矩阵的DCA排序所有轴的梯度长度最大为3.93(表3),故选择单峰模型(CCA)。蒙特卡罗检验结果显示,所有典范排序轴通过统计检验(F=1.85,P=0.002),说明地形因子对七渡口沙化高寒草地植物种分布具有显著影响。CCA排序前4轴代表不同的地形因子梯度变化,根据地形因子与植物排序轴的相关性大小,海拔、坡度、坡向、坡位与第一轴呈显著正相关关系,相关性大小排序为坡度>坡位>海拔>坡向,表明第一轴主要反映坡度的梯度变化。坡向、海拔、坡度、坡位与第二轴呈显著正相关关系,相关性大小排序为坡向>海拔>坡度>坡位,表明第二轴主要反映坡向的梯度变化(表4)。前向选择与Monte Carlo检验结果表明,坡度是沙化高寒草甸群落植物种分布的最重要地形因子,其次是坡向、海拔、坡位(表5)。
物种-环境因子CCA排序直观的表明了沙化草地物种分布与地形因子的关系(图4),CCA排序第一轴代表坡度地形梯度,从左往右坡度逐渐升高。第二轴代表坡向地形梯度,从上往下代表物种分布对坡向的依赖减小。垂果南芥、青藏大戟、喜马拉雅嵩草、长花马先蒿分布在CCA排序图的右上侧,受坡度、坡向影响较大。白花枝子花、二裂委陵菜、肉果草分布在CCA排序图第一轴左端,绳虫实、短筒兔耳草分布在CCA排序图右下侧,受坡度影响较小。早熟禾、西藏微孔草、天蓝韭、细叶亚菊、矮生忍冬位于CCA排序图的左下方,受坡度、坡向影响较小,主要分布在平坦的河漫滩。

图4 沙化高寒草地群落物种与环境因子的CCA排序图Fig.4 CCA two-dimensional ordination diagram for species and environmental variables of alpine grassland communitiesAsp:坡向,Aspect;Alt:海拔,Altitude;Pos:坡位,Slope position;Slo:坡度,Slope;Poa:早熟禾,Poa annua;Eld:披碱草,Elymus dahuricus;Les:赖草,Leymus secalinus;Els:老芒麦,Elymus sibiricus;Avs:燕麦,Avena sativa;Ajt:细叶亚菊,Ajania tenuifolia; Hea:阿尔泰狗娃花,Heteropappus altaicus;Cod:绳虫实,Corispermum declinatum;Chi:小白藜,Chenopodium iljinii;Pos:西伯利亚蓼,Polygonum sibiricum;Saw:牛耳风毛菊,Saussurea woodiana;Pom:圆穗蓼,Polygonum macrophyllum; Hyl:细果角茴香,Hypecoum leptocarpum;Arp:垂果南芥,Arabis pendula;Het:藏荠,Hedinia tibetica;Pob:二裂委陵菜,Potentilla bifurca;Eua:青藏大戟,Euphorbia altotibetica;Hin:矮生忍冬,Lonicera minuta;Alc:天蓝韭,Allium cyaneum;Tha:高山唐松草,Thalictrum alpinum;Kor:喜马拉雅嵩草,Kobresia royleana;Ths:高山野决明,Thermopsis alpina;Mot:西藏微孔草,Microula tibetica;Drh:白花枝子花,Dracocephalum heterophyllum;Pel:长花马先蒿,Pedicularis longiflora;Lat:肉果草,Lancea tibetica;Cot:糙果紫堇,Corydalis trachycarpa;Lab:短筒兔耳草,Lagotis brevituba;Rat:高原毛茛,Ranunculus tanguticus
3 讨论
3.1 沙化高寒草地的群落特征
植物是沙漠化发展各阶段中的最敏感和直观的标志,沙漠化的发展与植被的变化紧密相连、同步发展[18]。植物群落的逆行、进展演替实质上对应着沙漠化的正、逆过程[19]。随着沙化程度的加剧,土壤种子库密度下降,群落多样性明显降低,草地植被表现出对寒冷、干旱、放牧等胁迫的生态适应性[7, 20]。群落结构趋向简单,典型高寒草甸优势种和伴生种逐渐退出群落,而适应沙质荒漠的物种逐渐占据群落中的优势地位,灌丛化明显[9,21],群落出现湿生—中生—旱生的演替过渡格局[22]。随着草地沙化程度的加剧,植被组成发生变化,多年生草本植物呈逐渐减少,一年生草本植物呈逐渐增加的趋势。中度沙化阶段是草地植物群落特征变化的重要转折时期[23]。长江源区七渡口沙化高寒草地群落同样表现出类似的规律:29个植物种隶属16科,27属,单科植物占27.59%。赖草、细叶亚菊、绳虫实等耐旱植物种逐渐代替了牛耳风毛菊、二裂委陵菜、长花马先蒿等地带性草地植物,总体上优势种比例低于稀有种且以一年生草本为主,植被群落处于高度演替阶段。禾本科、莎草科等可食牧草逐渐减少和杂类草盖度急剧增加的趋势反映了高寒草甸退化演替过程植被变化的基本特征[24]。七渡口沙化草地优势种赖草大量分布,这与若尔盖高寒草地沙化演替植被分异规律相似[25]。草场沙化导致半固定沙地面积不断扩大,呈斑块状分布。作为典型的克隆植物,赖草是沙化生境里具有代表性且分布广泛的根茎型禾草,以风蚀形成的裸沙斑块作为根茎扩展的资源。随着草地退化过程加剧,赖草种群以聚集格局为主,体现了种内相互协作的个体关系[26]。赖草的扩张,毒杂草细叶亚菊、沙生植物绳虫实等指示种的出现预示七渡口高寒草地沙化处于进展之中,植被群落向旱生方向演替。
3.2 沙化高寒草地物种演替与风沙微地貌的关系
高寒草地沙化指出现以风沙活动和沙漠景观为主要特征的草地退化过程,同时也是一个土壤质地粗粒化、干早化和贫瘠化过程,并伴随着土壤自身固碳潜力降低[27]。风沙活动形成的各种洼地、沙丘等风蚀、风积地貌是沙生植被生存发展的重要载体,沙质地表基质的流动性和风沙活动伴随着沙地植被演替的各阶段[28],在强风沙活动时期,细颗粒物质被搬运、侵蚀,地表粗化,固定和半固定沙丘向半流动和流动沙丘演变,风蚀带走植被生长所需营养的细颗粒物质[29]。在弱风沙活动时期,细颗粒物质缓慢堆积于植被周围,其携带的养分促进了植被的发育[30]。风蚀和沙埋通过叶水势、叶片含水量及气孔导度调节光合速率和蒸腾速率,最终影响骆驼刺的水分利用效率[31]。随着沙面固定程度的增加,虫实倾向于将更多的资源和能量用于繁殖生长[32]。风沙地貌过程与土壤、植被演替变化在不同时空尺度密切联系[33],形成一个复杂动态的生物地貌互馈系统[34]。

表3 沙化高寒草地群落物种变异的DCA排序

表4 沙化高寒草地群落物种变异的CCA排序
**P<0.01; *P<0.05

表5 环境因子的重要性排序和显著性检验结果
*P<0.05,**P<0.01
土壤种子库在荒漠草原区群落演替、更新以及生态恢复中有着非常重要的作用。相同气候区,不同地形对地上植被及土壤种子库有着直接影响,最终导致植被格局的变化。顶坡到底坡地上土壤种子库储量整体上均呈现出增加的趋势[35]。随着沙丘的固定,土壤种子库的密度逐渐增加,物种多样性增加,物种分布逐渐均匀。不同沙丘类型不同坡位土壤种子库密度各异[36]。种子库对沙丘生态系统植被恢复贡献率因沙丘类型而异,沙丘生态系统自然物理过程与种子库的耦联关系是未来重要的研究方向[37]。由此可见,风沙地貌过程通过影响生境土壤养分、叶片水分生理、个体生存策略及种子库密度调控着草地植被群落物种分布。本研究发现,物种标准频度分布在不同微地形表现各异,总体上优势种比例低于稀有种,缓坡地植物最多为18种,山麓地最少为14种,反映了微地貌在沙化草地植被群落物种分布中的重要作用。
3.3 沙化草地微地形与环境因子的关系
一般来讲,地形因子包括海拔、坡度、坡向、坡位等通过物质和能量的再分配,共同影响物种的分布格局[38],同时也是风沙地貌过程的重要影响因子。坡度是水平方向上水分和土壤养分流的驱动力,对土壤厚度、理化性质有显著的影响。坡度增大导致风对土壤表面的直接冲击力增大,同时也使得土壤颗粒沿坡面向下的分力增加,从而影响到坡面土壤风蚀量[39]。风蚀速率随坡度的增大而增加,固定沙丘迎风坡中部的植被空间异质性一般都小于其下部和上部,与迎风坡各部位坡度、蚀积强度变化以及坡面气流场特性对植物繁殖体传播的影响密切相关[40]。地形对澜沧江源区高寒草甸植物丰富度及其分布格局的影响的研究结论与本文一致[41],即坡度是长江源区沙化高寒草甸群落植物种分布的最重要微地形因子。
坡向是重要地形因子之一,通过影响光照、温度及土壤养分等进而影响群落植物种的组成和分布[42]。背风坡草本植被的数量特征和多样性指数均大于迎风坡,沙丘坡向、坡位、高度均对土壤肥力显著影响,浑善达克沙丘微地形使沙地植被具有超地带性群落性质[43]。古尔班通古特沙漠沙丘坡向和坡位对草本植物α多样性和β多样性时空格局均产生显著影响[44]。丘间地的土壤水分条件最好,丘中次之,丘上最低[45]。丘间低地的降雨补给量最多,由于微地形变化的影响,沙丘下部较顶部、上部的降雨补给量最小[46]。本研究坡度、坡向对沙化草地物种分布有显著的影响。在青藏高原高寒草甸植物群落中,不同地形下的土壤理化性质差异显著,且土壤理化性质对植物群落功能性状及功能多样性有着显著的影响[47]。综上所述,因研究尺度及区域自然地理条件的差异造成海拔、坡度、坡向、坡位地形因子对植物物种分布作用大小各异。
沙化高寒草地的人工恢复重建首先应重视乡土优势植物种的选择利用,其次充分挖掘土壤种子库在草地恢复中的潜力,最后应考虑风沙地貌过程与植被演替的关系,借助有利自然生境条件,因势利导,通过工程与生物相结合的综合措施实现草地植被恢复与生态修复目的。
4 结论
(1)沙化草地群落结构简单,不同微地形沙化草地物种组成各异,稀有种植物有14种,占总物种数的48.23%,优势种比例低于稀有种,暗示群落正处于高度演替阶段。赖草、细叶亚菊、绳虫实等植物种的出现指示七渡口高寒草地沙化处于进展之中,植被群落向旱生方向演替。
(2)TWINSPAN将沙化高寒草地划分为7个群丛类型:Ass.I赖草+阿尔泰狗哇花+牛耳凤毛菊Leymussecalinus+Heteropappusaltaicus+Saussureawoodiana,Ass.Ⅱ赖草+喜马拉雅山嵩草+绳虫实Leymussecalinus+Kobresiaroyleana+Corispermumdeclinatum,Ass. Ⅲ赖草+披碱草+细叶亚菊Leymussecalinus+Elymusdahuricus+Ajaniatenuifolia,Ass.Ⅳ赖草+高山野决明Leymussecalinus+Thermopsisalpina,Ass.Ⅴ老芒麦+早熟禾Elymussibiricus+Poaannua,Ass.Ⅵ老芒麦+批碱草Elymussibiricus+Elymusdahuricus,Ass.Ⅶ绳虫实Corispermumdeclinatum。7个群丛在DCA排序图上的分布呈现一定的规律性。
(3)CCA排序结果表明,坡度和坡向是沙化高寒草地物种分布微地貌条件中的主导因子。
