稻虾共作水体浮游植物群落结构特征分析
2019-03-11宋庆洋米武娟王斌梁黄宇波程咸立马达文毕永红
宋庆洋 米武娟 王斌梁 黄宇波 程咸立 马达文 毕永红
(1. 中国科学院水生生物研究所, 淡水生态与生物技术国家重点实验室, 武汉 430072; 2. 中国科学院大学, 北京 100049;3. 湖北省水产技术推广总站, 武汉 430070)
克氏原螯虾(Procamarus clarkia)俗称小龙虾,具有生长快、抗逆性强, 对环境要求较低, 杂食性且繁殖力强的特点, 成为近年来重要的水产养殖品种[1]。目前我国克氏原螯虾的养殖模式主要有稻虾共作、池塘高密度精养、虾蟹混合养殖等[2]。稻虾共作是一种新兴的农业生产模式, 将种植业和养殖业有机结合起来, 充分利用稻田空间, 光、热、水及生物资源, 达到水稻和克氏原螯虾双丰收的目的[3]。浮游植物是稻田生态系统中的重要生物类群之一, 稻田浮游植物具有吸收磷固定氮、增加土壤有机质含量、改善土壤、促进水稻生长的作用[4]。浮游植物的生长繁殖易受到环境因子如温度、光照、营养盐浓度的影响, 除此之外人类活动和水生动物也会对浮游植物的生物量和群落组成造成影响[5—7]。稻虾综合种养作为一种复合生产模式, 共存于稻田生态系统中, 作为对水环境和小龙虾养殖具有重要影响的浮游植物, 其在稻田生态系统中的动态演变规律受到关注和重视; 但目前关于稻田种养水体浮游植物的研究较少。为了认识稻虾种养水体浮游植物群落结构组成及其变化规律, 掌握种养结合模式下浮游植物的基本情况, 为藻相调控和水质管理提供指导, 特开展本研究, 获取具体数据资料, 为稻田种养的健康发展提供理论依据。
1 材料与方法
1.1 实验材料
于2016年4月至12月逐月对4处稻虾共作水体采集水样并分析。如图 1所示, JYK采样地位于潜江市积玉口镇(112.57°E, 30.46°N), 大小为1.7×104m2; XK01和XK02采样地位于潜江市熊口镇(112.79°E, 30.24°N), 大小分别为1.8×104和2.1×104m2; CH采样地位于洪湖市汊河镇(113.49°E,19.95°N), 大小为2×104m2。

图 1 采样地分布Fig. 1 Sketch map of sample stations in the Jianghan Plain
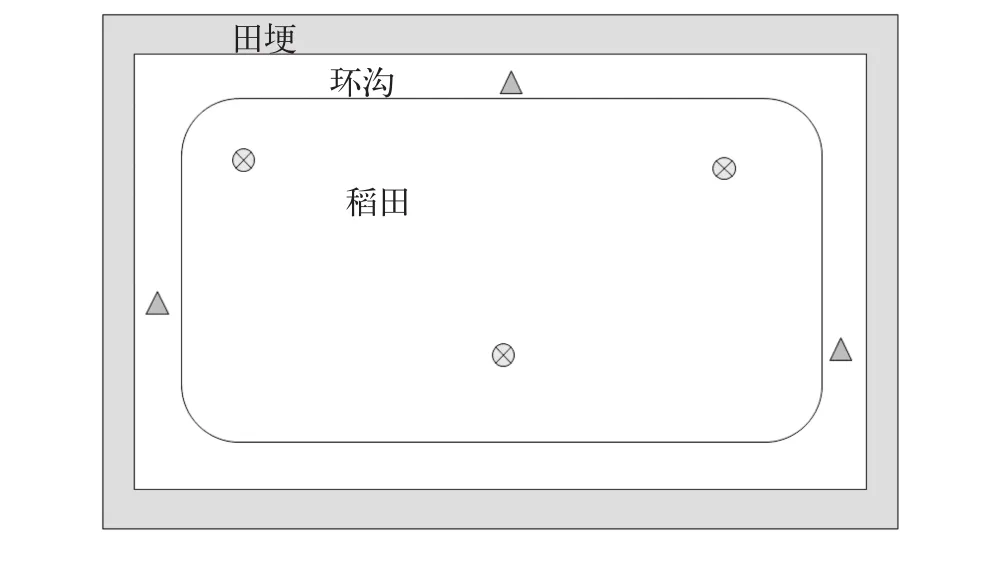
图 2 稻虾共作监测采样点示意图Fig. 2 Schematic illustration of paddy field and sample stations in the IRCSF system
稻田工程如图 2所示, 种稻面积约2×104m2, 环沟面积约0.33×104m2, 环沟深度为1.0—1.5 m, 宽度约3.0—4.0 m。环沟内种植水花生、水葫芦、轮叶黑藻等水生植物, 田埂种植芝麻、蚕豆等农作物。各采样稻田均为“稻虾共作”的种养模式,是指在稻田里养殖二季小龙虾并种植一季中稻。即在11至次年5月只养殖小龙虾,6至10月份种植中稻同时养殖小龙虾。其中6—10月水稻生长季节稻田和环沟水深有差异, 环沟水深约1.0 m, 小龙虾主要在环沟内活动; 非水稻生长季节环沟水深约1.5 m,田面水深约0.5 m, 小龙虾在环沟和稻田均有分布;但稻田和环沟水体相互连通, 并不是完全分割相互不交换的。水稻种植密度为低密度, 18株/m2, 小龙虾的放养密度9—12尾/m2, 饵料投喂只集中在4—6月, 各采样地均以投喂小龙虾专用颗粒饲料为主, 日投饵量约为存虾总重量的1%—4%, 投喂时全池均匀泼洒。
1.2 实验方法
水样采集参照《淡水浮游生物研究方法》[8]并结合稻虾共作水体环境特征进行, 采样点设置如图 2所示, 每个稻田设置6个采样点用来代表该样本的具体情况。测定的主要水质指标包括水温、总磷、正磷酸盐、总氮、铵态氮、硝态氮等。使用25号浮游生物网采集定性样品; 定量样品使用5 L有机玻璃采水器采集表层水, 取1 L混合水样置于聚乙烯水样瓶中, 加入10 mL鲁哥氏液静置48h以上后, 除去上清液, 浓缩到50 mL后取0.1 mL置于浮游植物计数框计数。
浮游植物样品定性分析鉴定参照《中国淡水藻类》[9]等资料进行。参照《淡水浮游生物研究方法》对浮游植物样品计数及计算。
多样性分析采用香农威纳(Shannon-Weiner)指数H′进行表示:
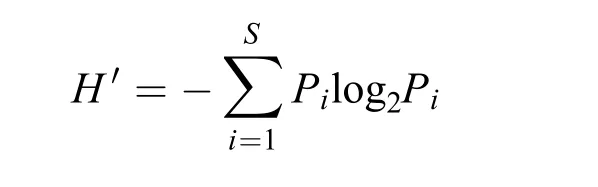
优势度计算公式:Y=ni/N·fi。式中Pi=ni/N,ni为物种i的个体数,N为群落样本个体总数,Pi为第i种个体数占总个体数的比例,S为群落中物种总数,fi为该种在该地区出现的频率, 在本研究中fi默认为1。本研究中把Y>0.1的种类定为优势种,Y>0.4的种类为强优势种, 0.01<Y<0.1的种类定为常见种。
数据分析、处理以及图表制作使用软件SPSS 19.0和微软office软件Excel 2010。单因素方差分析用于分析各样本间的时空差异。
2 结果
2.1 种类组成
共鉴定出浮游植物7门124种, 其中绿藻78种、蓝藻16种、硅藻15种、裸藻3种、隐藻2种、甲藻7种、金藻3种。各样点的藻类种类数逐月呈现先增加后减少的趋势, 但种类数最多的月份不同(图 3)。CH样点种类数最多的月份为6月, 为50种;JYK和XK01样点种类数最大值均出现在9月, 分别为52种和37种; XK02样点种类数最大值出现在8月,为38种。各月份蓝藻门种类数量最多, 其次为绿藻门和硅藻门的种类。金藻门的种类只在春、秋两季出现, 其他种类各月份均有出现。
图 4为调查期间各样地的浮游植物种类组成情况, 浮游植物种类数最多的是JYK, 为100种, 种类数最少的是XK01, 为83种。研究期间各样点共有种类数为56种, 其中CH和JYK两点共有种类最多, 为80种, XK01和XK02两点共有种类最少, 为68种。
2.2 细胞密度

图 3 稻虾共作水体浮游植物种类数Fig. 3 Richness of phytoplankton in the integrated rice-crayfish symbiosis farming system

图 4 稻虾共作水体浮游植物种类组成相似性Fig. 4 Similarity of phytoplankton species inthe integrated ricecrayfish symbiosis farming system
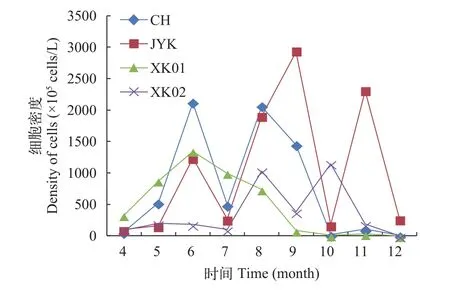
图 5 稻虾共作水体浮游植物细胞密度逐月变化Fig. 5 Monthly variation of cell density for phytoplankton monthly in the integrated rice-crayfish symbiosis farming system
如图 5所示, 细胞密度变化范围在1.37× 105—2.93×108cells/L, 细胞密度最大值出现在6—9月份,最小值出现在12月份和4月份。CH样地6月份细胞密度最大, 达到2.06×108cells/L, 12月份藻类细胞密度最低, 为1.14×106cells/L; JYK样地9月份细胞密度最大, 达到2.93×108cells/L, 4月份细胞密度最小,为1.03×107cells/L; XK01样地6月份细胞密度最大,达到1.34×108cells/L, 12月份细胞密度最小, 为2.17×107cells/L; XK02样地8月份细胞密度最大, 达到1.03×108cells/L, 12月份细胞密度最小, 为1.37×105cells/L。图 6显示在研究期间, 蓝藻和绿藻门藻细胞密度占据较高的比例, 其他门藻细胞密度占有比例相对较低; 各样地各门浮植物游细胞密度百分比波动变化较大, 同1月份各样地各门浮游植物细胞密度存在较大差异。
2.3 叶绿素a含量
如图 7所示, 叶绿素变化范围为0.15—208.60 μg/L。叶绿素a含量分别在6月和8月份出现2个峰值,在4—6月份和7—9月份呈现先升后降的趋势,9—12月份除JYK以外其他样地叶绿素a含量均呈现下降的趋势。
2.4 优势种组成
如表 1所示, 研究期间各样地出现的优势种有蓝藻门的泽丝藻、颤藻、蓝纤维藻; 绿藻门的小球藻、栅藻、实球藻, 硅藻门的小环藻、菱形藻等。4个采样地中均出现的优势种有: 泽丝藻、颤藻、栅藻和隐藻, 各月份出现的优势种差异较大。颤藻在7月和8月份成为CH和XK01采样点优势度较高的的单一优势种, 而JYK和XK02样点出现的优势种较多, 优势度相对较低。
2.5 多样性指数
如图 8所示, 研究期间各样地Shannon-Wiener多样性指数变化范围在0.64—6.3, CH、XKO1、XKO2样地的多样性指数在6—10月间均表现出先降低后升高的趋势, 在10—12月间先降低后升高。根据生物多样性阈值的分级评价标准[10], 研究期间所有样地的多样性等级在Ⅰ级以上。JYK样地多样性在4—10月份间处于Ⅳ级以上, 其他样地在Ⅱ—Ⅴ级之间波动。

图 6 不同月份各样地浮游植物细胞密度百分比Fig. 6 Aboundance of phytoplankton ineach sample station
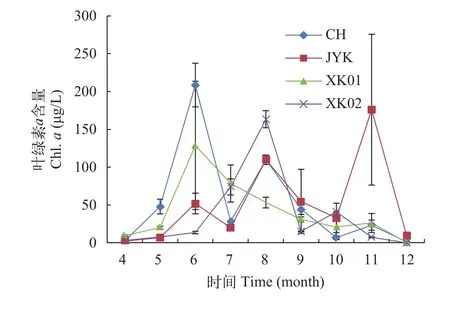
图 7 稻虾轮作水体中浮游植物叶绿素a含量的逐月变化Fig. 7 Chlorophyll-a content in the integrated rice-crayfish symbiosis farming system
2.6 浮游植物与环境因子的相关性
如表 2所示, 各样地水温最大值为33.9, 最小值为10.09; 总磷最大值为1.10, 最小值为0.02; 正磷酸盐最大值为0.59, 最小值为0; 总氮最大值为11.01,最小为0.26; 铵态氮最大值为8.71, 最小值为0; 硝态氮最大值为0.42, 最小值为0.01。
表 3所示, 对稻虾共作水体浮游植物和环境因子的相关性分析表明, 各稻田水体浮游植物种类数和温度具有显著的正相关关系, 细胞密度和总氮具有显著的正相关关系, 叶绿素a含量和总磷具有显著的正相关关系, CH采样点多样性指数和温度呈显著的负相关, JYK和XK02采样点多样性指数和温度呈显著的正相关关系。
3 讨论
3.1 浮游植物种类组成及优势种
稻虾共作水体中浮游植物种类较为丰富, 研究期间共鉴定出浮游植物7门124种, 少于郑敏[11]对稻鸭共育水体的研究中共鉴定6门230种, 多于程慧俊[2]对克氏原螯虾稻田养殖生态学研究中鉴定出7门86种, 也多于李岩[5]对稻蟹共生水体研究中鉴定出8门61种。不同水体浮游植物种类数的差异可能跟研究水域及其时空跨度等有关。本研究中各样地的浮游植物种类数在4—12月呈现先增后降的趋势, 不同季节浮游植物种类数呈现夏季>春季>秋季>冬季的规律; 该结果与程慧俊[2]在5—10月份对养虾稻田的研究结果相同, 与郝俊等[12]对河蟹生态养殖水体的研究相接近。大多数浮游植物最适生长温度在18—25℃[13], 江汉平原夏季稻田水温正好处于该范围内, 适宜各种浮游植物的生长, 因此, 物种数在夏季最高。优势种是藻类群落中在营养盐、光线和其他环境因子上具有一定竞争优势的种类[14, 15],自然湖泊中低温时一般绿藻、硅藻占据优势地位,高温时蓝藻快速生长并成为优势种类[16, 17], 本研究中硅藻只在秋、冬季作为优势种出现, 与程慧俊[2]研究结果一致, 蓝藻和绿藻在大多数月份成为优势种, 这可能跟稻田水体较浅春季升温较快有关。以上结果表明, 稻虾共作水体浮游植物种类丰富, 不同类型稻田种养水体所鉴定出浮游植物种类数存在较大差别, 浮游植物种类数总体为夏季种类数最多, 春季次之, 秋冬季最少的规律, 大多数月份的优势种由蓝藻和绿藻门的种类组成, 稻田的浅水环境对于浮游植物群落具有显著影响。
浮游植物Shannon-Wiener多样性指数变化范围在0.64—6.3, 不同季节间存在明显的波动变化,总体呈现春、秋季较高, 夏季较低的规律, 与武秀国等[18]、李瑞娇等[19]对于养殖池塘水体浮游植物多样性指数变化规律接近。水体富营养化会导致浮游植物的大量繁殖, 高温季节蓝藻的竞争能力强,繁殖速度快, 因而成为优势度较高的优势种降低浮游植物的多样性[20]。因此稻虾种养水体浮游植物多样性存在显著的季节差异(P<0.05), 蓝藻生长增殖并成为优势种是造成夏季浮游植物多样性较低的原因。
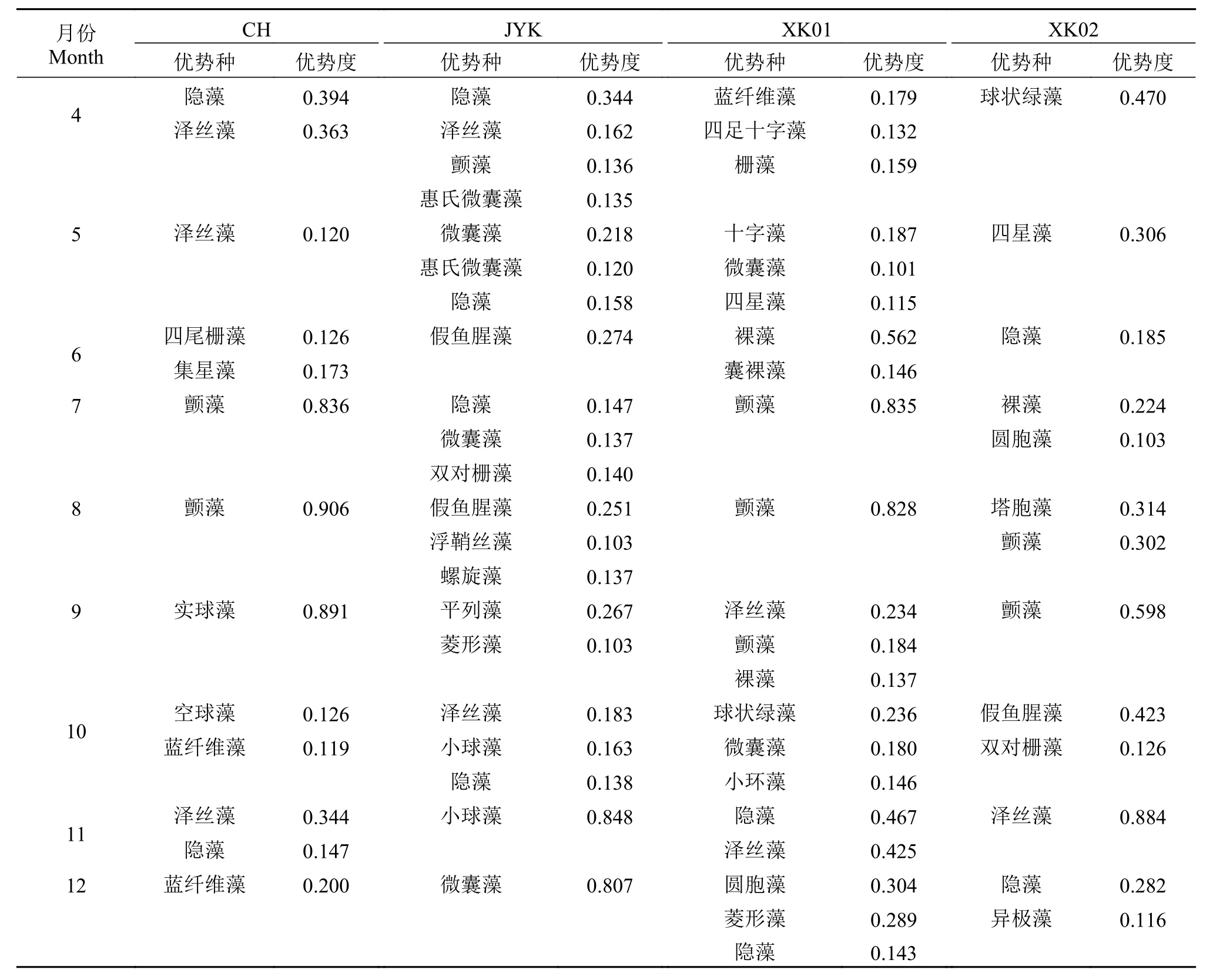
表 1 稻虾共作水体浮游植物优势种的组成Tab. 1 Dominant species of phytoplankton in the integrated rice-crayfish symbiosis farming system
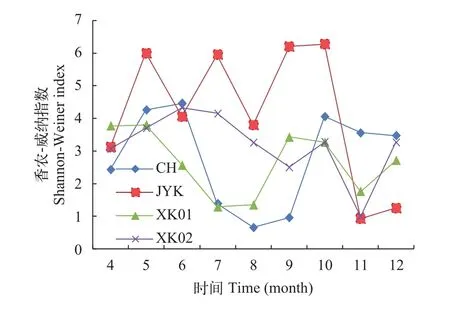
图 8 稻虾共作水体浮游植物多样性指数的逐月变化Fig. 8 Biodiversity index of phytoplankton in the integrated ricecrayfish symbiosis farming system
3.2 浮游植物细胞密度及叶绿素a含量
浮游植物细胞密度和叶绿素a含量在夏季最高、春秋季次之, 冬季最低。 该结果与多数养殖水体浮游植物研究结果[2, 12, 21]相同。与程慧俊[2]研究结果一致, 不同样地的浮游植物细胞密度和叶绿素a含量在6月出现1个峰值, 7月份降低, 8月份回升,9月份及以后逐渐降低。氮、磷等营养盐是浮游植物生长不可缺少的元素[22, 23], 各样地浮游植物细胞密度、叶绿素a含量与总氮、总磷呈显著的正相关关系, 说明稻虾共作水体中氮、磷与浮游植物群落的密切相互关系, 这种相互关系与其他水体中的情况一致, 氮磷营养仍然是稻田生态系统中影响浮游植物群落的关键因子。4—6月份, 随着养殖活动的进行, 向水体中投入大量饵料, 残饵和小龙虾粪便向水体中释放的氮和磷营养促使浮游植物大量繁殖。7月份是水稻插秧的初期, 可能是由于稻田中引入了新的水源造成浮游植物细胞密度和叶绿素a含量的下降[24]。有研究表明高温对能够促进底泥向养殖水体中的营养盐释放[25, 26], 8月份无养殖投饵, 但浮游植物生物量较高, 可能是由于环沟底泥中氮磷等营养元素持续释放导致的。XK01样地在8月份叶绿素a含量显著低于其他样地(*P<0.05), 未出现上升的趋势, 可能与其环沟内大量生长的水花生、水葫芦有关, 多数研究表明浮水植物具有很好的抑藻效果以及营养盐去除能力[27—29]。9月份及以后水温逐渐降低, 温度成为限制浮游植物生长的因素。

表 2 稻虾共作水体理化指标Tab. 2 Physico-chemical parameters in the integrated rice-crayfish symbiosis farming system

表 3 浮游植物各指数与环境因子Pearson相关性系数Tab. 3 Pearson’s correlation coefficient between phytoplankton indexes and environment factors
综上所述, 浮游植物细胞密度和叶绿素a含量在夏季达到最大值, 春、秋季低于夏季, 冬季最低。氮、磷是稻虾共作水体浮游植物生长的限制因子。
4 结论
稻虾共作水体中浮游植物种类较为丰富, 浮游植物种类数总体为夏季种类数最多, 春季次之, 秋冬季最少, 大多数月份优势种为蓝藻和绿藻; 浮游植物多样性存在较大的季节差异。浮游植物生物量在夏季最大, 冬季最低。氮、磷是稻虾共作水体浮游植物生长的限制因子。
