细鳞斜颌鲴5种组织2种同工酶的电泳分析
2019-03-07张涛甘金华周剑光张林陈建武何力
张涛,甘金华,周剑光,张林,陈建武,何力
(农业部水产品质量安全风险评估实验室(武汉),农业部淡水鱼类种质监督检验测试中心,中国水产科学研究院长江水产研究所,武汉 430223)
鱼类同工酶的遗传学研究始于20世纪60年代初期,其作为一种生化指标,已广泛应用于鱼类的物种和杂交种鉴定、倍性鉴定、亲缘关系比较、系统分类及胚胎发育过程中基因的表达与调控、杂交育种以及种群遗传结构分析等理论研究和实际应用领域[1-2]。
细鳞斜颌鲴(Xenocyprismicrolepis),隶属鲤形目(Cypriniformes)、鲤科(Cyprinidae)、鲴亚科(Xenocyprininae)、斜颌鲴属(Plagiognathops),广泛分布于长江、珠江、黑龙江等流域,为中小型水体中下层鱼类。由于其具有适应性强、食物链短、生长速度快、繁殖力强、营养价值高、不与草鱼等主养品种争食,又能摄食草鱼不能利用的腐殖质和底栖藻类等特点,于20世纪70年代前后成为优良的大水面引进增殖品种之一。随着人工繁殖的成功,如今细鳞斜颌鲴已成为一个优良的养殖品种。目前,关于细鳞斜颌鲴的研究,主要集中在生物学特性和增养殖技术[3-5]、年龄与生长[6]、繁殖特性[7]、细胞遗传特性[8]、分子生物学特性[9]、营养学特征[10]以及生理特性[11-12]等方面。有关细鳞斜颌鲴生化遗传特性的研究,主要集中于从生化遗传学角度探讨鲴亚科鱼类的亲缘关系及进化地位和细鳞斜颌鲴几种组织乳酸脱氢酶(lactatedehydrogenase, LDH)的表达情况。朱蓝菲[13]发现细鳞斜颌鲴和黄尾密鲴的LDH谱形非常相似,细鳞斜颌鲴♀×黄尾密鲴♂所获杂交种不仅成活率高,而且可繁殖后代。曹丽琴等[14]对鲴亚科6种鱼的骨骼系统和同工酶进行了研究,发现其生化分类和经典分类结果完全一致。杨品红等[15]对细鳞斜颌鲴各主要组织的LDH酶带进行了分析,发现酶谱的表达具有明显的组织特异性。酶带共有3个基因座位编码(LDH-a、-b、-c),肌肉和肝脏中分别发现1条和7条带,其余组织均发现3条带。张林等[16]报道了湖北丹江水库细鳞斜颌鲴的肌肉和肾脏中LDH同工酶的表达情况,发现肾脏和肌肉组织中各有5条LDH酶带表达,均由LDH-A和LDH-B两个基因编码。上述研究在一定程度上丰富了细鳞斜颌鲴生化遗传方面的研究内容,但并没有确定其特征生化遗传参数。本研究通过不连续聚丙烯酰胺凝胶电泳技术对细鳞斜颌鲴5种组织的LDH和苹果酸脱氢酶(malate dehydrogenase,MDH)进行了分析,旨在更全面地探索细鳞斜颌鲴的LDHMal和MDH表达特征,同时筛选出细鳞斜颌鲴种质的特征生化遗传参数,从而更方便地从生化遗传角度鉴定细鳞斜颌鲴的种质特性,以期为细鳞斜颌鲴的种质标准制定、遗传育种以及种质资源保护利用提供科学依据和理论基础。
1 材料与方法
1.1 实验材料
实验用细鳞斜颌鲴于2016年采自浙江湖州,共30尾,均为人工养殖的成鱼,体重范围175.66~323.78 g,体长范围20.3~27.31 cm,所有样本鱼均健康无伤病。
1.2 实验方法
1.2.1 组织酶液的制备
组织酶液的制备方法参照贺刚等[17]的方法,依据实验情况酌情考虑变动。首先对所测样本中10尾鱼进行LDH同工酶的普遍筛查,结合初步筛查结果,若确定某种组织LDH同工酶是单态,再对所测样本中另10尾样本鱼的该种组织该同工酶进行验证。若验证样本中,该种组织LDH同工酶同样是单态,则可初步确定该组织LDH同工酶可作为细鳞斜颌鲴种质的特征生化遗传参数。再随机选取剩余样本鱼为试验对象,研究MDH同工酶在不同组织中的表达情况。
剪鳃放血后在鱼尚存活时,冰浴条件下取心脏、晶状体、肌肉、肾脏和肝脏,组织经预冷的生理盐水冲洗干净后用滤纸吸干水分,放入低温冰箱(-80 ℃)保存备用。
样品称重后放入洗净预冷的匀浆器内,按1∶3(g/mL,m/V)加双蒸水,冰浴条件下反复研磨至成浆状,在4 ℃、16 097 g条件下离心3次,每次30 min,分装上清液,-80 ℃保存以备电泳。
1.2.2 电泳方法
同工酶分析采用不连续聚丙烯酰胺凝胶电泳方法,电泳时所有样品上清液稍微解冻后与甘油溴酚蓝混合点样。LDH(酶编号:E.C.1·1·1·27)和MDH(酶编号:E.C.1·1·1·37)。浓缩胶浓度和分离胶浓度分别为4.0%和7.5%。电泳在4 ℃条件下进行,电泳缓冲系统为Tris-Gly系统,pH 8.3,每孔点样量为20 μL。恒压电泳,起始电压为280 V,30 min后将电压调至220 V,待溴酚蓝条带被电泳至胶底停止电泳,电泳时间为9.5 h。
1.2.3 染色及成像
电泳结束后,将凝胶板取下于室温避光染色,LDH和MDH的染色分别参照张涛等[18]和贺刚等[19]的方法。待出现清晰条带后,将凝胶板用去离子水漂洗2~3次。将漂洗后的凝胶板平放在自制灯箱上用尼康数码相机(J5 10-100)拍照,并根据电泳图谱结合相对迁移率绘制酶谱模式图。
1.3 酶谱分析
同工酶的缩写、基因座位和等位基因的命名参照Shaklee等[20]的方法,即以同工酶名称缩写的大写代表酶蛋白,小写代表编码基因。控制同一种酶的不同基因座位据其相对迁移率Rf由大到小依次编号为1~10,酶带分析参考熊全沫[21]的方法。
2 结果与分析
2.1 LDH的表达
从图1A可以看出,10尾样本鱼心脏组织共检测到8条LDH酶带,其中LDH1~LDH6表达活性相对较强,其余两条酶带表达活性相对较弱,LDH1~LDH6为10尾样本鱼的共有酶带,LDH3和LDH4为同一位点的两个等位基因编码。LDH7和LDH8仅在1~4号鱼和7~10号鱼中有检出。LDH1、LDH2和LDH5表达活性相对较强。10尾样本鱼晶状体组织中LDH表达的酶谱相同(图1B),都是单态,且为经典的5条LDH酶带,从阳极向阴极依次对应为LDH1—B4、LDH2—A1B3、LDH3—A2B2、LDH4—A3B1和LDH5—A4。其中LDH1和LDH2表达活性相对较强,其余3条酶带表达活性相对较弱。LDH在10尾样本鱼肌肉组织中表达程度不高,共检测到4条LDH酶带(图1C),除1号鱼仅检测到2条LDH酶带外,其余9尾样本鱼均各检测到4条LDH酶带,而且2~4号样本鱼的LDH1和LDH2表达活性相对较弱,而9号和10号样本鱼LDH1和LDH2表达活性相对较强,所有样本鱼的LDH3和LDH4表达活性相对较强。LDH在10尾样本鱼肾脏组织中共检测到5条LDH酶带(图1D),其中5号鱼未检测到LDH酶带,其余9尾样本鱼LDH酶谱相同,也是经典的5条LDH酶带,其中LDH1和LDH5表达活性最强,其余LDH酶带表达活性相对较弱。LDH在细鳞斜颌鲴肝脏中的表达较为复杂(图1E),10尾样本鱼共检测到8条LDH酶带,而且表现出多态,LDH8由C基因编码[22],2号样本鱼LDH酶带数最多,共检测到6条LDH酶带,LDH1~LDH3为一独立基因编码区。
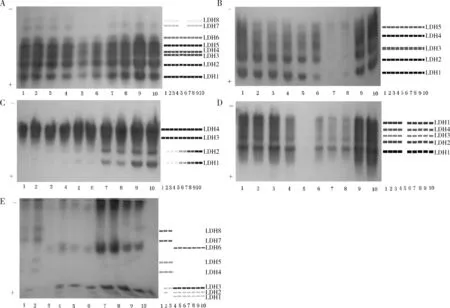
图1 细鳞斜颌鯝LDH电泳图谱A、B、C、D、E分别表示心脏、晶状体、肌肉、肾脏和肝脏的LDH酶谱;1~10泳道分别表示不同个体的酶谱。Fig.1Electrophoretogram of LDH isozymes in Xenocypris microlepisA,B,C,D,E: electrophoretograms of LDH isozymes expressed in heart, eye, muscle, kidney and liver respectively.1~10: the zymograms of different individuals.
2.2 MDH的表达
细鳞斜颌鲴各组织MDH同工酶的表达如图2所示,心脏、肝脏和肾脏组织中MDH酶带可分为上清液型(s-MDH)和线粒体型(m-MDH),晶状体和肌肉组织中未检测到上清液型s-MDH。心脏组织中共检测到8条MDH酶带(图2 H1、H2),m-MDH6在2号鱼有表达,在1号鱼未检出,m-MDH3表达活性最强,m-MDH4次之,其余MDH酶带表达活性相对较弱。晶状体组织中共检测到6条m-MDH酶带(图2 E1、E2),m-MDH3表达活性最强,其余MDH酶带表达活性相对较弱,m-MDH4在2号鱼中有表达,在1号鱼中未检出。肌肉组织中共检测到8条MDH酶带(图2 M1、M2),m-MDH5表达活性较强且在1号鱼中有表达,在2号鱼中未检出,除m-MDH5外,m-MDH3、m-MDH4以及m-MDH10表达活性相对较强,其余酶带表达活性程度相对较弱。肝脏组织中检出MDH酶带数较少(图2 L1、L2),共检测到4条,而且2尾样本鱼MDH酶谱相同,m-MDH3表达活性程度最强,m-MDH6表达活性最弱。肾脏组织中也检测到4条MDH酶带(图2 K1、K2),2号鱼MDH酶带数最少,仅3条;m-MDH4在1号鱼中有检出,在2号鱼中未检出;m-MDH3表达活性最强。
2.3 细鳞斜颌鲴特征生化遗传参数
本研究中,10尾样本鱼的心脏、肌肉、肾脏和肝脏组织中LDH酶带均有多态,表现在不同样本鱼的酶带数不同,或者酶带数相同但表达程度有差异,而所有样本鱼晶状体LDH不仅酶带数相同,而且分离效果相对较好,酶带清晰,可初步确定以晶状体LDH作为从生化遗传特征层面鉴定细鳞斜颌鲴种质的备选对象。随机选择另外10尾样本鱼的晶状体进一步电泳以验证晶状体LDH酶带表达情况(图3),结果全为单态,所以本研究认为可以将晶状体组织LDH作为鉴定细鳞斜颌鲴种质的特征生化遗传参数。
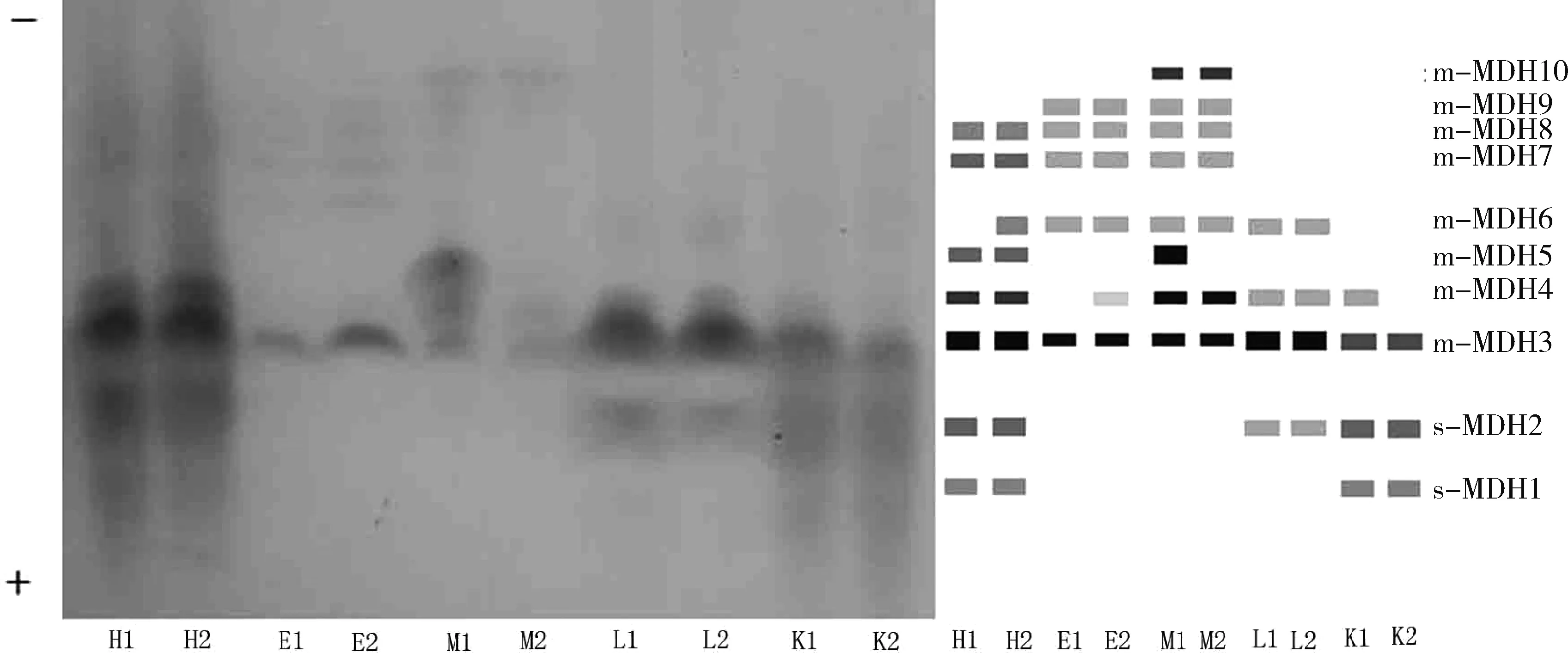
图2 细鳞斜颌鯝MDH电泳图谱H1、H2、E1、E2、M1、M2、L1、L2、K1和K2分别表示1号鱼和2号鱼心脏、晶状体、肌肉、肝脏和肾脏中的MDH酶谱。Fig.2Electrophoretogram of MDH isozymes in Xenocypris microlepisH1, H2, E1, E2, M1, M2, L1, L2, K1, K2: electrophoretograms of MDH isozymes expressed in heart, eye, muscle, liver and kidney from theNo.1 and No.2 specimen respectively.
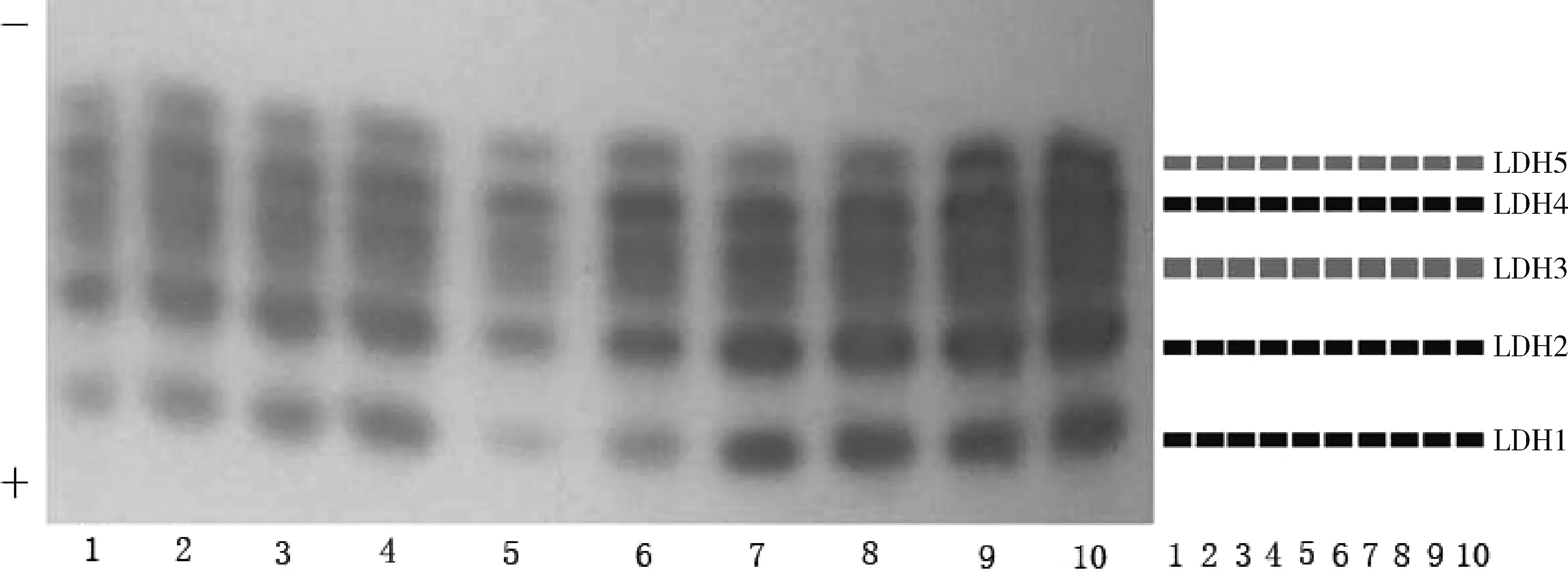
图3 细鳞斜颌鲴晶状体LDH电泳图谱1~10泳道分别代表不同个体的酶谱。Fig.3 Electrophoretogram of LDH isozymes expressed in eyes from Xenocypris microlepis1~10: the zymograms of different individuals.
3 讨论
3.1 同工酶表达具有组织特异性
本研究发现细鳞斜颌鲴的心脏组织、晶状体组织、肌肉组织、肝脏组织和肾脏组织中均能检测到LDH同工酶和MDH同工酶的表达,表明其在细鳞斜颌鲴体内分布较广。从研究结果来看,2种同工酶的表达均有组织特异性,主要表现在:同一种同工酶在同一尾样本鱼不同组织中是否表达、表达的酶带数以及活性程度不同。如心脏中共检测到8条LDH酶带(图1A),而肌肉中共检测到4条LDH酶带(图1C);MDH在心脏中的表达程度比晶状体中要高(图2)。B4、A1B3、A2B2、A3B1和A4这5条经典的LDH同工酶酶带的生理功能不同,尽管都催化乳酸和丙酮酸之间的相互转化,但靠近阴极的A4的主要功能是催化乳酸脱氢后转化为丙酮酸,丙酮酸在线粒体中进行有氧氧化,以产生更多的能量供给组织需要,而靠近阳极的B4主要参与无氧酵解,催化丙酮酸还原成乳酸[23],一般在耗氧量大的器官或组织中A4含量较大,而在对氧气需求量低或无需氧气的器官或组织中B4含量相对较高。本研究中,细鳞斜颌鲴心脏中LDH酶带数超过经典的5条带,可能是由于重复基因的存在和表达,该现象在其他鱼类中也有发生,如在鲑(Samonsala)和部分鲤形目鱼类(Cypriniformes)中发现有LDH的重复位点(A1和A2,B1和B2)[24]。本研究认为,细鳞斜颌鲴心脏组织中靠近阴极的2~3条带是A4重复基因的表达,心脏中LDH酶带数比肌肉中多的原因可能是:心脏作为耗氧量较大的器官,其中A4含量较高,在LDH同工酶酶谱中表现为,心脏组织在靠近阴极的部位出现比肌肉中多的酶带。不同组织同工酶的特异性与其代谢功能密切相关。MDH是一种在糖代谢、三羧酸循环过程中扮演重要角色的酶类,它能使苹果酸脱氢后参与到细胞中糖酵解后的有氧代谢反应中,心脏组织中有氧代谢旺盛[25]。本研究中,细鳞斜颌鲴心脏MDH表达活性较晶状体中强,与两种组织中有氧代谢旺盛与否或者说两者行使的生理功能密切相关。分析认为造成同工酶表达具有组织特异性的原因主要是:同工酶是基因表达的产物,其表达受到温度、压力、激素、氧容量和营养等多种内外因素调控,致使该基因在各组织间的表达时间和强度不相一致,造成酶谱特异性[26]。
鱼类LDH同工酶通常由3个基因编码,其中A、B基因所编码的酶出现在鱼类的多种组织中,而C基因位点具有组织差异性,其编码的蛋白质电荷量与A、B不同[22, 27-28],C基因编码的酶在鲤形目鱼类中只存在于肝脏中[13],有的学者称之为“肝带”[22],在电泳行为上,“肝带”一般表现为向阴极迁移。本研究中细鳞斜颌鲴肝脏LDH8由C基因编码,离阴极端较近,与白甲鱼[29]和草鱼[30]的实验结果类似。
3.2 与已有研究结果的比较分析
关于细鳞斜颌鲴LDH同工酶的研究已有相关报道[13-16],本研究与之相同的地方为都证实了细鳞斜颌鲴LDH同工酶具有组织特异性,而且肝脏中都存在C基因编码的同工酶(本研究认为曹丽琴等[14]发现的阳极侧有特异的F4同工酶存在即C带);不同之处在于各组织中LDH同工酶酶带数存在差异。朱蓝菲[13]的研究表明细鳞斜颌鲴心脏、晶状体和肾脏中LDH同工酶酶带数均为3条,而曹丽琴等[14]发现上述3种组织LDH同工酶酶带数均为经典5条带;张林等[16]发现细鳞斜颌鲴肌肉和肾脏中LDH同工酶酶带数均为5条;杨品红等[15]发现细鳞斜颌鲴心脏、晶状体、肌肉、肝脏和肾脏组织中LDH同工酶酶带数分别为3条、3条、1条、7条和3条。本研究发现细鳞斜颌鲴心脏、晶状体、肌肉、肝脏和肾脏组织中LDH同工酶酶带数分别为6~8条、5条、2~4条、4~6条和5条,与杨品红等[15]的研究结果存在一定的差异。分析造成这一现象的原因,除了试验样本不同外(野生样本或养殖群体),可能还与实验方法不同有关,如凝胶浓度不同,导致凝胶孔径不同,使电泳过程中酶蛋白经过胶孔的分离效果有差异;同时,增大电压或延长电泳时间可能会使一些来不及分离的酶蛋白达到有效分离,如杨品红等[15]所采用的电压和电泳时间分别为150 V、5~6 h,而本研究分别为220 V、9.5 h。当然不排除制样过程中的操作产生的差异,如冰浴匀浆、离心等。如对肝脏的离心,笔者认为应尽可能增加离心次数,以排除干扰物。
除了LDH同工酶,本研究还报道了细鳞斜颌鲴各主要组织的MDH同工酶的表达情况,和大多数硬骨鱼类一样,细鳞斜颌鲴的MDH也存在互相不形成异聚体的移动较快的细胞质型(s-MDH)和移动较慢的线粒体型(m-MDH)两种类型,但仅存在于除了晶状体和肌肉组织外的其他3种组织中。电泳条件下细胞质型(s-MDH)泳动速度要比线粒体型(m-MDH)快。曹丽琴等[14]发现细鳞斜颌鲴心脏、晶状体、肌肉、肝脏和肾脏组织中MDH同工酶酶带数分别为5、4、5、4和6条,而本研究中上述5种组织MDH同工酶酶带数分别为8、6、8、4和4条。分析认为,造成这种差异的主要原因可能是实验方法不同,两者的电泳条件(电压和电泳时间)和电极缓冲液类型及pH均不同,同时,不排除样本来源存在地理种群差异的可能性。MDH是生物糖代谢的关键酶之一,能催化草酰乙酸与苹果酸之间的相互转化[30-31]。本研究中肝脏组织MDH表达活性程度比晶状体组织强(图2中L1、L2的m-MDH3的着色程度较E1、E2的m-MDH3着色深),可能与肝脏行使重要的生理功能有关,杨书婷等[32]在研究雌核发育白鲢(Hypophthalmichthysmolitrix)时也发现类似的现象。
本研究较为系统地报道了细鳞斜颌鲴几种组织中LDH和MDH的表达情况,通过细鳞斜颌鲴两种同工酶酶谱结果分析其生化遗传特征参数,可为制定其种质标准及其杂交育种提供参考依据,目前这一技术已广泛应用于鲴亚科以及其他鱼类中[13,33-34]。
3.3 同工酶表达具有多态性
本研究中,除了晶状体和肾脏外,同种组织在不同个体间的LDH表达酶带数不同,除肝脏组织外,心脏、眼睛、肌肉和肾脏组织中的MDH表达酶带数不同,不同个体同一位点酶带的表达活性也不同,如5、6号鱼和9、10号鱼肌肉组织LDH1和LDH2酶带的表达活性程度不同,表明细鳞斜颌鲴LDH和MDH具有多态性。同工酶的多态性已见不少研究报道[26,32,35],余敏等[35]研究发现,10尾云南高背鲫(Carassiusauratus)样本鱼心脏中EST-5只在部分鱼中表达,而另一部分样本鱼中未表达,存在明显的个体差异。同工酶是基因表达的产物,不同个体同种组织中同工酶的表达与否以及表达程度强弱受到一些因子的调控。同工酶在同一物种、不同个体的相同组织中的表达差异可能与其生长发育阶段[36]、健康状态[37]、所行使生理功能[32]等均有关联,以便于适应环境,更广泛地消化和利用底物[26]。正因为同工酶存在个体差异性,借助同工酶研究物种鉴定与分类、亲缘关系比较以及群体遗传多样性水平时,需对多个样本进行验证分析,避免被因个体差异和样本量不足造成假象而误导。
3.4 晶状体组织LDH为特征生化遗传参数
通过同工酶分析可较深入、直接地了解物种的遗传信息,是分析生物的发育、遗传、系统进化和生理生化过程的有效探针。人们对同工酶的认识是从LDH开始的,LDH也是研究最透彻的同工酶,而MDH是生物糖代谢的关键酶之一,这两种同工酶在机体执行生理功能中扮演着重要的角色。本研究中细鳞斜颌鲴晶状体组织LDH在各实验鱼样本中均为单态,表明LDH在细鳞斜颌鲴晶状体组织中的表达稳定,应符合可遗传性的特点,故选取晶状体组织LDH作为细鳞斜颌鲴的特征生化遗传参数。
尽管同工酶电泳技术是研究物种遗传变异、系统发育以及生理生化过程的重要手段之一,且具有快速有效、花费较少等优点,但由于同工酶是蛋白质水平的标记,是对基因的间接反映,检测结果往往是被修饰的基因产物,而非基因本身[38]。随着分子生物学技术的发展,DNA分子标记内容逐渐丰富,应用领域也越来越广,尽管花费相对昂贵,但它是DNA水平上遗传变异的直接反应。因此,建议在进行物种种质鉴定、生理生化过程、群体遗传变异分析以及系统发育进化等研究时,将同工酶电泳技术与分子生物学技术结合起来,使两者相互印证,取长补短。
4 结论
通过采用不连续丙烯酰胺凝胶垂直板电泳,对细鳞斜颌鲴心脏、晶状体、肌肉、肝脏和肾脏LDH和MDH同工酶进行研究,发现细鳞斜颌鲴LDH和MDH两种同工酶均呈现出组织特异性:所测样本鱼的心脏、晶状体、肌肉、肾脏和肝脏中LDH酶带数分别为6~8、5、2~4、5和4~6条,MDH酶带数分别为7~8、5~6、7~8、3~4和4条。其中,晶状体组织LDH可作为细鳞斜颌鲴特征生化遗传参数。本研究为制定细鳞斜颌鲴种质标准,从生化遗传角度鉴定其种质提供参考依据。
