干旱胁迫对6种园林灌木光合生理特性的影响
2019-01-19吴雪仪陈红跃黄永芳
吴雪仪 陈红跃 黄永芳
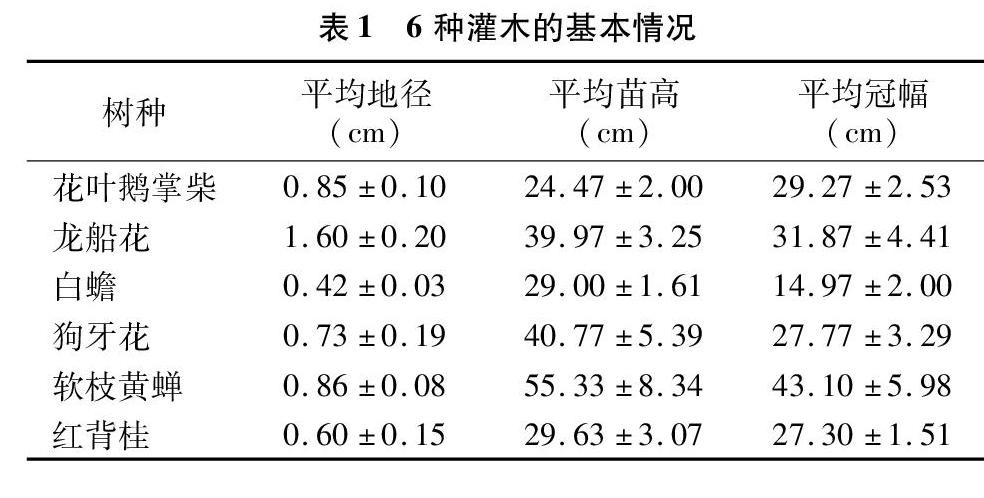

摘要:以花叶鹅掌柴、龙船花、白蟾、狗牙花、软枝黄蝉和红背桂6种园林灌木为材料,研究人工模拟干旱胁迫环境对植物光合指标和叶绿素荧光动力参数的影响,结合主成分分析法综合评价其抗旱能力。结果表明,(1)干旱胁迫处理期间,6种灌木的净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)、和蒸腾速率(Tr)持续下降,复水后软枝黄蝉的Ci未能得到有效恢复,其余植物各光合指标均有回升。(2)光合系统Ⅱ(PSⅡ)的有效光化学量子产量(Fv′/Fm′)、实际光化学量子效率[Y(Ⅱ)]、表观光合电子传递速率(ETR)、光化学淬灭(qP)随着胁迫强度加重而降低,复水后各项指标均有所恢复,其中花叶鹅掌柴的Y(Ⅱ)、ETR和qP、毛杜鹃的Fv′/Fm′和Y(Ⅱ)、龙船花和红背桂的Fv′/Fm′、狗牙花的ETR均恢复至对照水平。6种灌木的非光化学淬灭(NPQ)持续上升,复水后白蟾有所下降,但仍显著高于对照,其余植物均恢复到对照水平。(3)主成分分析结果表明,6种园林灌木的抗旱性强弱顺序为花叶鹅掌柴>白蟾>狗牙花>红背桂>软枝黄蝉>龙船花。
关键词:园林灌木;干旱胁迫;光合指标;叶绿素荧光动力参数;主成分分析
中图分类号: S684.01 文献标志码: A 文章编号:1002-1302(2019)22-0142-06
随着城市化快速发展进程,生态环境质量呈下降趋势。同时,由于城市自然空间的过度开发破坏了自然水循环过程[1],城市有限的水资源在面对日益增长的城市园林绿化面积时变得更为紧缺。另外,城市特有的环境特征衍生出了多种新型绿化类型,如雨水花园、立体绿化和边坡绿化等,它们的生态环境决定了植物经常处于水分亏缺的状态[2-4]。光合系统Ⅱ(PSⅡ)对干旱胁迫的响应非常敏感[5]。干旱胁迫会引起植物气孔部分或全部关闭,进而导致蒸腾速率以及CO2吸收量、光合速率下降[6-7],严重时外界CO2不能进入植物细胞内,继续照光引起植物体内光能过剩,导致光合器官受到破坏,进而影响光合系统Ⅱ的活性、结构和功能。而具有抗旱性的植物会启动自身保护机制,通过光呼吸和热耗散等非光化学过程消耗过剩光能[8-10]。
本试验以华南地区常见的园林灌木花叶鹅掌柴(Schefflera odorata cv. variegata)、龙船花(Ixora chinensis)、白蟾(Gardenia jasminoides var. fortuniana)、狗牙花(Ervatamia divaricata)、软枝黄蝉(Allemanda cathartica)和红背桂(Excoecaria cochinchinensis)为材料,通过研究干旱胁迫对6种灌木的光合特性和叶绿素荧光参数的影响,揭示干旱胁迫对其光合生理的影响机制,为筛选耐旱性强的园林绿化植物与节水灌溉提供理论依据,对于城市绿化和城市可持续发展研究具有重大意义。
1 材料与方法
1.1 试验材料
试验在华南农业大学板栗园温室大棚内进行,试验苗为花叶鹅掌柴、龙船花、白蟾、狗牙花、软枝黄蝉和红背桂的实生容器苗,营养袋直径15 cm,高15 cm,基质中荷兰土与黄土为1 ∶ 1。苗木生长基本情况见表1。
1.2 试验方法
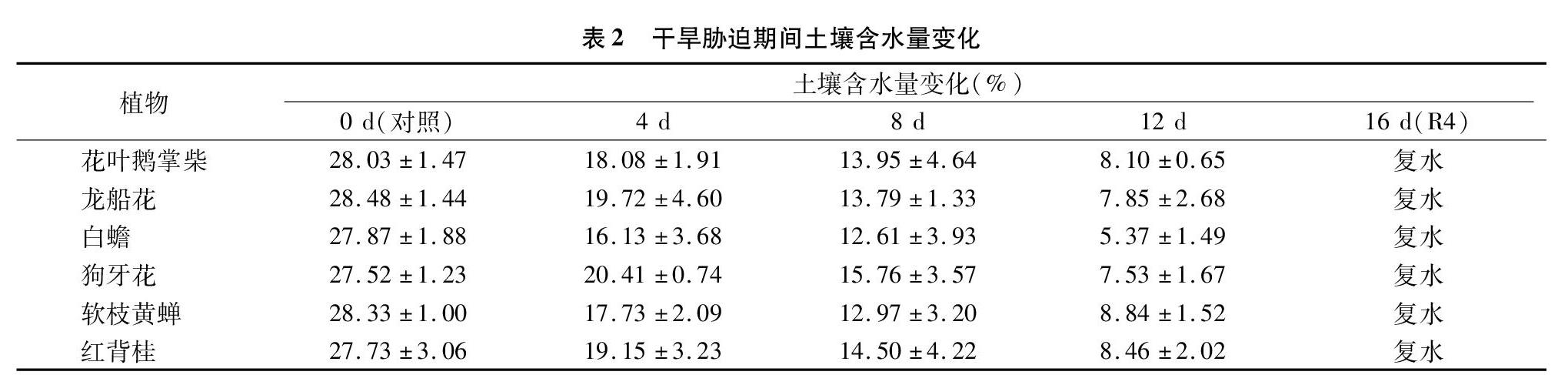
1.2.1 试验设计 2017年10月采用人工模拟自然干旱胁迫的方法,试验前1 d浇水至饱和状态,往后11 d内不再浇水,直至12 d时复水(即浇水至饱和状态)。在停水后0(对照)、4、8、12 d及复水后4 d测定各项光合指标和荧光指标。干旱胁迫期间6种灌木营养袋中土壤含水量变化见表2。
1.2.2 测定方法 选取每株的第3张至第8张成熟功能叶挂牌标记,每次测定时间为09:00—11:00,此期间光线充足且变化不大。光合指标的测定仪器为LI-6400便携式光合仪(LI-Cor,Inc,美国),采用开放式气路,内置光源光照度为 1 200 μmol/(m2·s),空气流速500 L/min,叶温30 ℃。试验测定的光合指标有净光合速率(Pn)、蒸騰速率(Tr)、气孔导度(Gs)和胞间CO2浓度(Ci)等参数,所有参数9次重复。
采用PAM-2500调制式叶绿素荧光仪(WALZ,德国),于09:00—11:00,在光适应下测定最大荧光(Fm′)、最小荧光(Fo′)、 PSⅡ实际光化学量子效率Y(Ⅱ)、表观光合电子传递速率(ETR)。19:30—21:30(天黑后)测定叶片暗适应下的初始荧光(Fo)和最大荧光(Fm)。根据公式计算出以下荧光动力参数:PSⅡ有效光化学量子产量(Fv′/Fm′)=(Fm′-Fo′)/Fm′;光化学淬灭系数(qP)=(Fm′-F′)/(Fm′-Fo′);非光化学淬灭系数(NPQ)=Fm/Fm′-1。
1.3 数据处理
采用Microsoft Excel和IBM SPSS Statistics 19.0软件对试验数据进行统计分析。对原始数据标准化后进行主成分分析,根据特征值>1的提取原则确定主成分个数[11]。各主成分得分是其加权值,权重是主成分所对应的贡献率,根据主成分的特征向量计算出各抗旱指标的得分,按得分评定各树种的抗旱能力。
2 结果与分析
2.1 干旱胁迫对6种灌木光合指标的影响
2.1.1 干旱胁迫对植物叶片净光合速率的影响 干旱胁迫处理期间,6种植物的叶片净光合速率(Pn)随干旱胁迫时间的延长急剧下降,均在胁迫4、8、12 d显著低于对照,并在胁迫12 d时降到最低值(图1)。其中,以龙船花的下降幅度最大,降幅为84.21%,其次是软枝黄蝉、白蟾和狗牙花,分别为82.39%、70.42%和67.64%。经过4 d复水处理后,6种灌木叶片的Pn均显著增加,但仍显著低于对照。
Pn是反映植物光合生理对逆境响应能力的重要指标[14],干旱胁迫下所有植物的Pn均会下降,且抗旱性强的植物Pn的降低程度低于抗旱性弱的植物[15]。6种灌木的Pn均随着干旱胁迫的加剧而逐渐下降,其中花叶鹅掌柴降幅最小,说明其抗旱性较强,而龙船花降幅最大,说明其抗旱性较差。干旱胁迫下Pn下降可能是因为干旱胁迫导致气孔关闭,使CO2进入叶片受阻,即气孔限制(stomatal limitations)[16-17];也可能是植物叶肉细胞光合活性下降所造成的非气孔限制(non-stomatal limitations)[18]。根据Farquhar等提出的观点[19],当叶片的Ci和Gs变化趋势均为下降,则Pn下降为气孔限制所致,Ci降低而Gs升高或不变的情况下为非气孔限制所致。本实验结果表明,干旱胁迫对6种植物叶片的Gs、Ci与对Pn的影响基本一致,说明Pn的下降是由于气孔限制所导致的。Gs在干旱胁迫下的降低有利于减少水分蒸腾[20],所以随着6种灌木Gs的不断下降,气孔开放程度持续降低,6种灌木的Tr也随之下降。
复水后,6种灌木植物叶片的Pn、Tr、Gs均有所恢复,除了软枝黄蝉,其余植物叶片的Ci也有回升,说明随着干旱胁迫的解除,植物叶片的气孔开放,CO2进入叶片的量增加,植物光合作用开始恢复。虽然软枝黄蝉的Gs有回升,但是叶片的Ci恢复较慢,可能是其光系统受害严重和植物自身恢复能力有限所致。
3.2 植物抗旱性与叶绿素荧光动力参数的关系
干旱胁迫下,PSⅡ会主动调节光化学反應效率和ETR以适应植物CO2同化能力下降的情况,从而减少过剩光能对光合系统的伤害[21]。PSⅡ有效光化学量子产量(Fv′/Fm′)代表植物光合反应中心PSⅡ的光合效率,反映了开放的PSⅡ反应中心原初光能捕获效率[22-23]。PSⅡ实际光化学量子产量 Y(Ⅱ) 反映了PSⅡ反应中心在部分关闭下所具有的实际原初光能捕获效率,也反映着PSⅡ反应中心的开放程度[24]。表观光合电子传递速率(ETR)值可以反映植物光合能量的传递速率,也可以表示植物光合能力[25]。化学淬灭(qP)代表了光合能量用于暗反应固定能量的部分[26]。干旱胁迫下,NPQ的提高是光合器官在干旱胁迫下的自我保护机制,通过热耗散防御光抑制对PSⅡ反应的破坏[27-28]。
在本试验中,6种灌木的Fv′/Fm′、Y(Ⅱ)、ETR以及qP均随着干旱胁迫的增强而持续降低,并且同时期NPQ值持续上升,表明6种灌木PSⅡ反应中心的开放程度、光能转化和利用效率均受到干旱胁迫的影响,其碳同化能力和激发能的捕捉效能也因此下降。qP的下降反映了6种灌木PSⅡ天线色素吸收的光能用于光合化学反应的比例降低,而NPQ的提高则表明6种植物均启动光破坏防御机制,通过增加热耗散防御过剩光能的破坏,保护光合器官。复水后,6种植物各项荧光指标均有所恢复,其中花叶鹅掌柴叶片的Y(Ⅱ)、ETR和qP、龙船花和红背桂的Fv′/Fm′、狗牙花的ETR均恢复至对照,除了白蟾外其他5种植物的NPQ均恢复至对照,表明12 d的干旱胁迫未对6种灌木的光合细胞造成不可逆转的损伤,且进一步表明花叶鹅掌柴在本试验中表现了较强的抗旱能力。
3.3 6种灌木抗旱性综合评价
综上所述,6种灌木的光合指标和叶绿素荧光动力参数均对干旱胁迫作出明显的响应。利用主成分分析法,根据对各个指标与植物抗旱性关系的分析得出,6种灌木的抗旱能力由强到弱依次为花叶鹅掌柴>白蟾>狗牙花>红背桂>软枝黄蝉>龙船花。对各抗旱指标的分析表明,光合指标中的Pn、Gs、Tr及叶绿素荧光动力参数中的Y(Ⅱ)、ETR、qP等指标在一定程度上可以反映植物受干旱的影响程度,可以作为比较6种灌木抗旱性的指标。
参考文献:
[1]俞孔坚,李迪华,袁 弘,等. “海绵城市”理论与实践[J]. 城市规划,2015,39(6):26-36.
[2]宋国萍,宋 川. 边坡生态防护用灌木植物耐旱性试验研究与应用[J]. 四川环境,2010,29(1):1-8.
[3]周 媛,徐冬云,董艳芳,等. 9种轻型屋顶绿化景天属植物的抗旱性研究[J]. 中国农学通报,2012,28(25):294-301.
[4]单 进,戴子云. 雨水花园植物选择与搭配研究[J]. 山西建筑,2017,43(1):207-209.
[5]李志军,罗青红,伍维模,等. 干旱胁迫对胡杨和灰叶胡杨光合作用及叶绿素荧光特性的影响[J]. 干旱区研究,2009,26(1):45-52.
[6]Lawlor D W,Cornic G. Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plants[J]. Plant,Cell & Environment,2002,25(2):275-294.
[7]周宇飞,王德权,陆樟镳,等. 干旱胁迫对持绿性高粱光合特性和内源激素ABA、CTK含量的影响[J]. 中国农业科学,2014,47(4):655-663.
[8]马瑞丽,胥生荣,陈 垣,等. 干旱胁迫对黄芪叶片光合特性和叶绿素荧光参数的影响[J]. 中药材,2018,41(3):531-534.
[9]胡宏远,王振平. 干旱胁迫对赤霞珠葡萄叶片水分及叶绿素荧光参数的影响[J]. 干旱区资源与环境,2017,31(4):124-130.
[10]Ahmed S,Nawata E,Hosokawa M,et al. Alterations in photosynthesis and some antioxidant enzymatic activities of mungbean subjected to waterlogging[J]. Plant Science,2002,163(1):117-123.
[11]林 丽,李以康,张法伟,等. 青藏高原高寒矮嵩草草甸退化演替主成分分析[J]. 中国草地学报,2012,34(1):24-30.
[12]马 新,李 铭,朱耀军,等. 干旱胁迫下文冠果光合指标的变化研究[J]. 河南农业科学,2017,46(1):122-126.
[13]韩瑞宏,卢欣石,高桂娟,等. 紫花苜蓿抗旱性主成分及隶属函数分析[J]. 草地学报,2006,14(2):142-146.
[14]刘泽彬,程瑞梅,肖文发,等. 水淹胁迫对植物光合生理生态的影响[J]. 世界林业研究,2013,26(3):33-38.
[15]Ao H,Zhang Y. Effects of water stress on photosynthetic characteristics of spruce[J]. Bulletin of Botanical Research,2007,27(4):445-451.
[16]魏清江,馮芳芳,马张正,等. 干旱复水对柑橘幼苗叶片光合、叶绿素荧光和根系构型的影响[J]. 应用生态学报,2018,29(8):2485-2492.
[17]谭晓风,卢 锟. 干旱胁迫对两种油桐幼苗生长、气体交换及叶绿素荧光参数的影响[J]. 生态学报,2017,37(5):1515-1524.
[18]张林春,郝 扬,张仁和,等. 干旱及复水对不同抗旱性玉米光合特性的影响[J]. 西北农业学报,2010,19(5):76-80.
[19]Farquhar G D,Sharkey T D. Stomatal conductance and photosynthesis[J]. Annual Review of Plant Physiology and Plant Molecular Biology,1982,33(3):317-345.
[20]卢广超,许建新,薛 立,等. 干旱胁迫下4种常用植物幼苗的光合和荧光特性综合评价[J]. 生态学报,2013,33(24):7872-7881.
[21]朱 弘,温国胜. 不同竹龄毛竹冠层叶片PS Ⅱ叶绿素荧光特性的比较[J]. 中南林业科技大学学报,2017,37(1):12-19.
[22]张仁和,郑友军,马国胜,等. 干旱胁迫对玉米苗期叶片光合作用和保护酶的影响[J]. 生态学报,2011,31(5):1303-1311.
[23]尤 鑫,龚吉蕊. 叶绿素荧光动力学参数的意义及实例辨析[J]. 西部林业科学,2012,41(5):90-94.
[24]鲁 松. 叶绿素荧光动力学在植物抗逆性研究中的应用[J]. 四川林业科技,2013,34(4):69-71.
[25]陈建明,俞晓平,程家安. 叶绿素荧光动力学及其在植物抗逆生理研究中的应用[J]. 浙江农业学报,2006,18(1):51-55.
[26]刘 艳,黄乔乔,马博英,等. 高温干旱胁迫下香根草光合特性等生理指标的变化[J]. 林业科学研究,2006,19(5):638-642.
[27]黄承建,魏 刚,徐建俊,等. 干旱胁迫对苎麻叶绿素荧光特性的影响[J]. 中国麻业科学,2014,36(1):41-45.
[28]孙景宽,张文辉,陆兆华,等. 干旱胁迫下沙枣和孩儿拳头叶绿素荧光特性研究[J]. 植物研究,2009,29(2):216-223.
