厚竹高生长期茎秆节部对基本组织细胞壁发育形成的影响
2019-01-14陈阿丽
夏 敏,陈阿丽,于 芬
(江西农业大学林学院/江西省竹子种质资源与利用重点实验室,江西南昌330045)
高生长是竹类植物生长发育的关键过程,与生物量密切相关。一些竹种在竹笋出土初期每天仅长高几厘米,以后逐渐加快,每天几十厘米,在高生长旺盛期一昼夜的高生长量可达1 m[1],并且在45~60 d内即可完成秆型的生长,最大生长高度可达15~20 m[2]。而细胞壁是植物细胞的重要结构,其结构成分和状态与植物细胞的伸长密切相关,植物细胞的伸长直接影响植物的个体生长。因此探究细胞壁的发育规律对揭示竹秆节间快速高生长的机制有重要意义。

图1 厚竹茎秆正常节和异常节外部形态Fig.1 External morphological maps of normal and abnormal sections of thick bamboo stems
厚竹(Phyllostachysedulis‘Pachyloen’)是毛竹(Phyllostachysedulis)的一种优良的变异品种,其不仅具有秆壁特厚、上部近实心、生物量大等特点,同时因其优良的竹材冲击特性和纤维性状等特征,使其具有重要的种质研究意义和综合开发利用价值[3-6]。在对厚竹进行研究的过程中发现部分厚竹具有节部异常的现象,异常节和正常节相比,在外部形态上主要表现为节间变短(图1)。竹秆的迅速高生长主要依赖于节部的居间分生组织。竹秆节部在竹子的生长发育过程中不仅具有机械支撑作用,而且物质的横向交流也主要发生在节部。竹子的高生长也与竹秆的节部密切相关。节间快速伸长主要是由细胞伸长引起的,细胞伸长与细胞壁结构和物质的变化息息相关。因此猜想,节部异常会不会通过影响细胞伸长过程中细胞壁结构和物质的变化进而影响节间的伸长,从而在外部形态上表现为节间缩短呢?目前,有关厚竹茎秆快速高生长过程中基本组织细胞壁的构建机制及节部对茎秆快速高生长的影响机理还不明确。因此,本文拟通过对比研究节部异常和节部正常的竹子高生长过程中细胞壁的发育对厚竹茎秆快速高生长过程中异常节和正常节节间中部基本组织细胞进行比较发育解剖学研究,了解基本组织细胞壁构建的细胞学基础,探究节部对高生长的影响,为进一步揭示茎秆快速高生长提供理论依据。
1 材料与方法
1.1 试验材料
1.1.1 光学材料 实验材料取自于江西农业大学竹类植物种质园厚竹实验基地。分别于2016年4月12日、2016年4月16日、2016年4月18日采取不同发育阶段节部正常和异常的厚竹笋5株,笋高分别为0.78、1.10、1.30、1.37、1.69 m。要求生长状态良好、生境相同、无病虫害。节数从基部开始算起,连续截取各个节间中部和节部的材料。
1.1.2 电镜材料 实验材料厚竹于2016年4月18日采自于江西农业大学竹类植物种质园厚竹实验基地。选取3株高1 m左右的竹笋作为样株,采取笋尖,然后从基部往上开始的各节间中部取1 mm3左右的小块。
1.2 方法
1.2.1 光学制片 材料选取后立即投入FAA固定液中固定,成熟材料用70%FAA,幼嫩材料用50%FAA,且幼嫩材料用酒精梯度进行脱水,二甲苯透明,常规石蜡切片法制作并于旋转切片机上做连续厚约8~10 μm的横、纵切片,番红-固绿对染,二甲苯透明,中性树胶封片,Zeiss AX10荧光显微镜下拍照。
1.2.2 透射电镜制片 材料选取后立即用2.5%的戊二醛和4%的多聚甲醛初固定,1%的锇酸后固定,磷酸缓冲液冲洗,酒精梯度脱水,spurr树脂包埋,于LKB-V型超薄切片机切片,H-7650型透射电镜观察拍照。
2 结果与分析
根据在光镜下观察的组织结构特征,将基本组织细胞的分化与发育过程分为两个时期:初生壁形成期、次生壁形成期。
2.1 初生壁形成期
该时期主要分为两个阶段,初生壁发育形成初期和初生壁发育形成后期。
从显微结构分析,初期,正常节和异常节基本组织并没有明显的差异。该时期主要以细胞分裂为主,体现为细胞数量的增加(图2),细胞长度变化不明显(图3,注:24、15、7是异常节)。从竹笋的纵切面上观察,顶端分生组织具有原套-原体结构(图4,A)和节部与节间分化(图4,B)。细胞排列整齐,细胞核较大且居中,部分细胞正处在有丝分裂状态(图4,C)。部分基本组织细胞和纤维细胞中能观察到双核仁的现象(图4,D)。细胞中含有大量的淀粉粒(图4,E),节部淀粉粒含量较节间多(图4,F)。
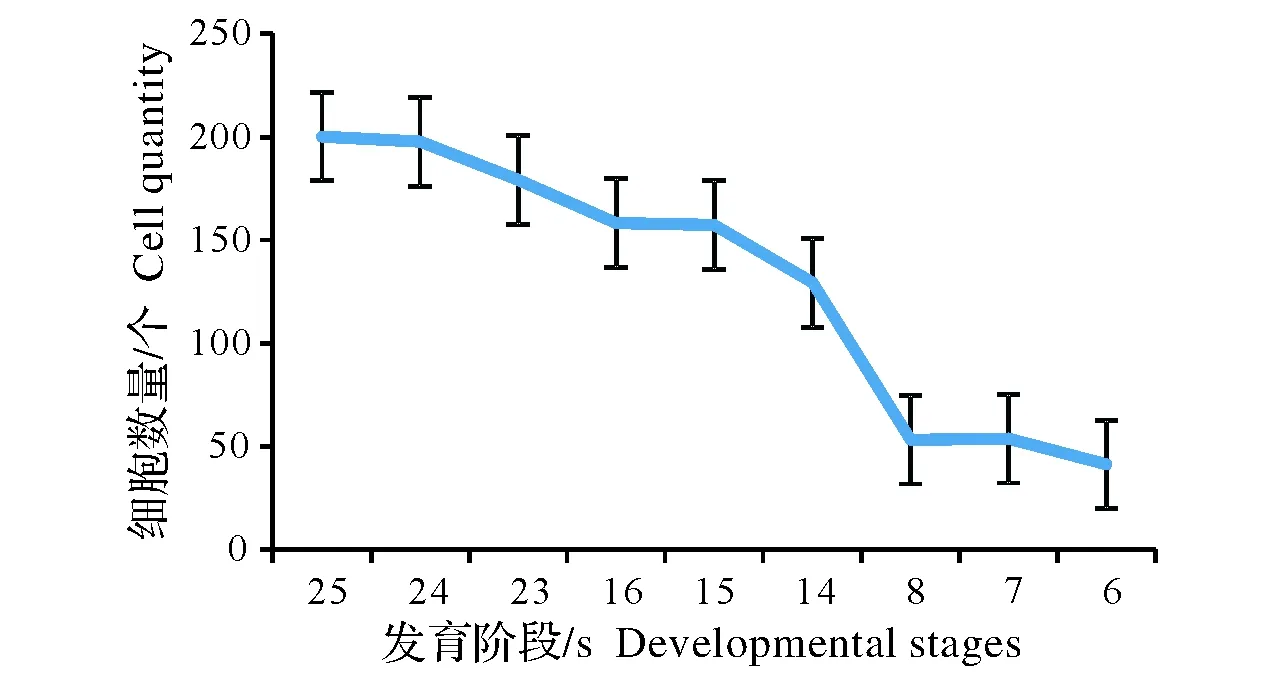
图2 不同发育阶段基本组织细胞数量的变化Fig.2 The varying of ground tissue cell quantity in different developmental stages

图3 不同发育阶段基本组织细胞长度的变化Fig.3 The varying of ground tissue cell length in different developmental stages
后期,主要以细胞伸长为主,节部异常引起细胞数量的增加和细胞长度的减小(图2,图3)。随着组织的发育,正常节长、短细胞的分化较明显(图5,A)。异常节较正常节而言长、短细胞分化不明显(图5,B)。两者细胞壁在荧光显微镜下观察均有增厚的现象(图5,C、D)。淀粉粒含量节部异常节较正常节多(图5,E、F)。
从超微结构分析,初期,基本组织细胞壁较薄,胞间连丝丰富,细胞中含有大量细胞器和运输小泡(图6,A)。节部异常节较正常节线粒体内嵴状结构有降解的现象(图6,B、C)。
后期,随着组织的发育,基本组织细胞壁加厚明显,细胞器丰富(图6,D)。节部正常的节基本组织细胞壁较节部异常节的细胞壁加厚明显,胞间连丝较丰富,物质运输小泡仍较多(图6,D、E、F)。
2.2 次生壁形成期
从显微结构分析,无论是正常节还是节部异常节,都有长、短细胞的分化且分化明显,短细胞细胞核边缘化分布(图7,A),长细胞的内容物多数已降解 (图7,B)。长细胞次生加厚明显(图7,C、D)。只有极少数细胞中还有零星的淀粉粒分布(图7,E、F)。
从超微结构分析,长细胞壁次生加厚明显,能观察到细胞壁不同的壁层,微管沿质膜分布于细胞壁的内侧(图6,G)。节部异常节较正常节而言,胞间层含有电子密度较低的白色物质(图6,H),细胞壁上有白色的凸起物质(图6,I)。
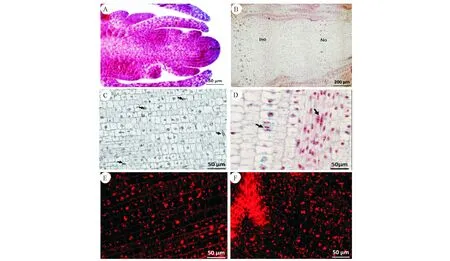
A:顶端分生组织;B:节间与节部基本组织的分化;C:部分基本组织细胞正在进行有丝分裂;D:基本组织和纤维细胞具有双核仁现象;E:基本组织细胞内含有较多的淀粉粒;F:节部淀粉粒的分布A:Apical meristem;B:Differentiation of ground tissue between internodes and nodes;C:Some groung tissue cells were undergoing mitosis;D:The groung tissue and the fiber cells have the double nucleoli;E:More starch grains were found in the ground tissue cells;F:The distribution of starch grains in the node图4 初生壁形成初期基本组织解剖结构Fig.4 Anatomical structure of ground tissue in early stage of primary wall formation

A:长、短细胞分化明显;B:长细胞细胞核边缘化分布;C:基本组织细胞壁加厚;D:节部异常节基本组织细胞壁加厚;E:基本组织细胞内淀粉粒减少;F:节部异常节基本组织细胞内淀粉粒含量较多.注:B、D、F 节部异常节A:The differentiation between long and short parenchyma cells was obvious;B:Marginal distribution of long cell nucleus;C:Thickening of ground tissue Cell walls;D:The cell wall was thickened in the ground tissue cell walls of abnormal node;E:Starch grains decreased in the ground tissue cells;F:The starch grains in the ground tissue cells of abnormal node were higher.Note:B、D、F abnormal node图5 初生壁形成后期基本组织解剖结构Fig.5 Anatomical structure of ground tissue in the late stage of primary wall formation

A:初生壁形成初期基本组织细胞壁较薄;B:胞间连丝丰富,细胞器较多;C:节部异常节线粒体结构;D:正常节胞间连丝丰富;E:初生壁形成后期基本组织细胞壁加厚明显;F:节部异常节运输小泡较少;G:基本组织细胞壁次生加厚;H:基本组织细胞壁上电子密度较低的白色凸起物质(如箭头所示);I:基本组织细胞壁胞间层电子密度较低的白色物质(如箭头所示).C、F、I 节部异常节;A、B、C 初生壁形成初期;D、E、F 初生壁形成后期;G、H、I 次生壁形成期A:The cell walls were thin in the early stage of primary wall formation;B:Rich plasmodesmatas and organelles;C:The mitochondrial structure of the abnormal node;D:The plasmodesmatas in normal node were rich;E:The cell walls were thickened obviously in the late stage of primary wall formation;F:The transport vesicles in abnormal node decreased;G:The long parenchyma cell walls were secondary thickening;H.White protruding material with low electron density on the cell walls of ground tissue (as shown in the arrow);I:A white substance with low electron density in the cell wall of the cell wall (as indicated by the arrow).C,F,I Abnormal node;A,B,C In the early of primary wall formation;D,E,F In the late stage of primary wall formation;G,H,I The secondary wall formation图6 基本组织细胞壁的超微结构Fig.6 Ultrastructure of the cell walls of the ground tissue
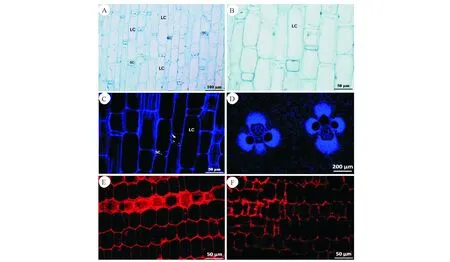
A:短细胞细胞核边缘化分布;B:长细胞的内容物多数已降解;C:基本组织长细胞壁次生加厚;D:示维管束横切面;E:基本组织细胞仍有零星淀粉粒分布;F:示节部异常节淀粉粒的分布.注:F 节部异常节A:Short cell nucleus marginalization distribution;B:Most of the contents of long parenchyma cells has been degraded;C:The long parenchyma cell walls were secondary thickening;D:Showing the transverse section of the vascular bundle;E:Ground tissue cells still have sporadic distribution of starch grains;F:The starch grains distribution of abnormal node.Note:F abnormal node图7 次生壁形成期基本组织解剖结构Fig.7 Anatomical structure of ground tissue in secondary wall formation
3 讨论与结论
3.1 高生长期基本组织细胞壁的发育形成
细胞壁是植物所特有的结构,其结构、成分和状态与植物细胞的伸长密切相关。
基本组织细胞壁在发育形成的过程中,细胞内含有大量的线粒体、内质网和高尔基体等细胞器。在细胞壁的边缘分布大量的运输小泡。内质网是分泌蛋白等物质合成、储存、加工和运输的场所,在新壁形成过程中,细胞壁物质经内质网和高尔基体等细胞器的合成加工,分泌运输小泡,再通过运输小泡和质膜融合通过胞吐的方式将物质释放出,参与细胞壁的形成[7]。
植物在生长发育的过程中,当可溶性糖的含量超过自身消耗所需时便会以淀粉的形式储存。在细胞活跃的时期,便会逐渐的降解为可溶性糖,为植物生长发育提供物质能源。厚竹初生壁发育形成后期,异常节较正常节淀粉粒含量较多,说明该时期异常节细胞代谢缓慢。在次生壁发育形成过程中,部分细胞中还含有极少数的淀粉粒,该时期淀粉粒通过降解为可溶性糖的形式为细胞壁的合成提供物质能量来源。在茎秆生长发育过程中,基本组织细胞始终存在丰富的胞间连丝,胞间隙也逐渐的变大且部分胞间隙能观察到有降解物的存在,有研究表明,胞间隙可作为质外体运输的有效途径[8]。说明基本组织细胞降解的产物通过胞间连丝和胞间隙两种途径来实现物质的胞间转移。贺新强和崔克明[9]通过对植物次生壁形成的研究认为,次生壁是细胞停止生长后在初生壁和质膜之间形成的具有木质素或其他次生代谢产物沉积的具有多层结构的细胞壁。基本组织长、短细胞的细胞壁存在很大的差异,长细胞次生壁逐渐加厚,能观察到细胞壁不同的壁层,且颜色加深明显。短细胞仅在与长细胞相邻的地方次生加厚,这和Liese等[10-13]的研究结果一致。以上结果表明:高生长期,基本组织细胞壁形成过程中,丰富的细胞器参与细胞壁物质的合成,如细胞壁物质经内质网和高尔基体等细胞器的合成,加工分泌运输小泡,运输小泡和质膜融合以胞吐的方式将物质释放出,参与细胞壁的形成,微管系统控制细胞壁物质的沉积排列方向。这个过程中,线粒体为其提供能量来源,淀粉粒的降解为其提供主要物质能源。物质的运输主要有两种方式,即共质体运输和质外体运输。
本实验通过对不同发育阶段相同部位细胞长度和细胞数量统计数据分析表明,节间伸长是由细胞分裂和细胞伸长共同决定的,细胞分裂是基础,细胞伸长是导致节间快速伸长的主要原因。本实验和前人的研究结果相符[14-15]。
3.2 节部在厚竹茎秆快速高生长中的作用
竹秆的迅速高生长主要依赖于节部的居间分生组织。竹秆节间维管束被基本组织细胞完全分开而相互游离,物质的横向运输和环流主要在节部进行[16-18]。许婷婷[19]对厚竹异常节和相邻正常节节部进行解剖结构分析发现,与正常节相比,异常节的节部韧皮结数量较多,而韧皮结是节部物质横向运输的重要部位,起到分流和缓冲的作用,故会影响细胞的生长发育。由于节隔的缺失,竹子茎秆节间和节部物质横向运输的渠道减少。通过解剖结构分析发现,在初生壁形成的后期,节部异常节较正常节而言,淀粉粒的含量较多,运输小泡数量较少,细胞壁加厚不显著,长、短细胞分化不明显。节间快速伸长主要是由细胞伸长引起的,而细胞的伸长需要物质和能量的供给。综上,节隔缺失通过引起节间和节部与物质运输有关结构的变化,使节间细胞伸长所需的物质和能量不能及时的供给,进而影响着节间的伸长,在外部形态上表现为节间缩短。
在次生壁发育形成期,节部异常节基本组织细胞壁与正常节相异之处为:胞间层具有电子密度较低的白色物质;细胞壁内侧含有电子密度较低的白色凸起物质。目前,还不能判断这两种物质为何物,将在前期研究基础上,研究两种物质的成分和功能,进一步揭示茎秆迅速伸长生长的细胞学基础。
致谢:在整个实验过程中,许婷婷、胡添翼、苗森浩及竹子资源与利用重点实验室的老师和同学们对研究给予了帮助,谨致谢意!
