流速对不同浮游藻类的生长影响研究
2019-01-09周静苟婷张洛红蓝郁马千里梁荣昌赵学敏
周静, 苟婷, 张洛红, 蓝郁, 马千里, 梁荣昌, 赵学敏,*
流速对不同浮游藻类的生长影响研究
周静1,2, 苟婷2, 张洛红1, 蓝郁3, 马千里2, 梁荣昌2, 赵学敏2,*
1. 西安工程大学环境与化学工程学院, 西安 710048 2. 环境保护部华南环境科学研究所, 广州 510655 3. 暨南大学生命科学技术学院, 广州 510632
为了研究流速对不同浮游藻类生长的影响, 于2015年3月采集东江流域广东惠州河段原水, 在环形有机玻璃水槽中分别培养铜绿微囊藻、斜生栅藻和小环藻, 通过调整水体流速, 研究不同流速对不同藻类的细胞密度、叶绿素a浓度、最大比增长率和叶绿素荧光参数Fv/Fm的影响。结果表明, 在不同流速条件下, 不同浮游藻类的生理生化特征有所差异; 铜绿微囊藻表现为在不同流速条件下生长状况都较弱; 斜生栅藻和小环藻在低流速(<0.075 m·s–1)条件下繁殖速度较快, 且随着水体流速的增加, 对藻类生长有促进作用, 而在较高流速(>0.075 m·s–1)条件下随着流速的增加, 藻类的生长受到抑制。可见, 不同持续的流动条件是影响浮游藻类数量和生理指标变化的重要原因。该研究为东江水源的流速流量调节控制富营养化和水华防治提供技术支撑。
东江; 流速; 浮游藻类; 富营养化; 水华
1 前言
由于高强度人类活动影响下大量氮磷营养盐进入水环境, 造成水体富营养化和水华频繁发生已严重损害了水生态系统服务功能[1]。水华的暴发与藻类的生理特点、营养盐、温度、光照、缓慢的水流条件及其诸多环境因素密切相关[2]。其中, 水文水动力的影响尤为显著[3-5], 水利工程的开发导致水体从河流型向水库型改变, 大坝拦截成库造成局部河段流速减缓, 引起营养盐的累积和藻类大量繁殖[6-7]。如长江三峡大坝截流后, 香溪河库湾由天然河流变为人工水库, 引起库区水生态系统变化暴发甲藻水华[8]; 福建省九龙江北溪江自东引水工程运行后, 2009年曾发生甲藻水华事件[9]; 丹江口水库大坝建成后, 导致汉江浮游植物群落组成以及细胞密度均发生显著变化, 1992年初春汉江下游江段发生硅藻水华[10]。
流速作为水动力条件中最基本、最直观的参数, 对浮游植物的生长和繁殖具有十分重要的影响[11]。研究表明[12-14]低流速有利于维持水环境的稳定、增加光照利用率和延长水力停留时间, 促进悬浮藻类的生长, 而过高的流速则抑制悬浮藻类的生长, 并且较大流速能有效地阻止水华的形成与发展, 消除已经形成的水华。因此, 通过流量管理和水力调度等方式控制富营养化和藻类水华已在河流水环境管理策略中得到广泛应用[15-16]。
东江是珠江水系三大河流之一, 是河源、惠州、东莞、深圳等地的主要供水水源, 并承担着向香港地区供水的重要任务[17]。近年来由于流域内经济发展、城市化加速导致水体污染物增加, 加之流域内的梯级开发, 东江部分河段水质下降明显, 干流富营养化程度从上游至下游呈逐渐加重趋势[18-19]。东江干流已开发的梯级电站13座, 最下游的一个梯级为东江水利枢纽(剑潭梯级), 东江下游污染物的迁移扩散主要受该梯级下泄流量的影响[20], 剑潭梯级也是距离下游东深供水取水口最近的一个梯级。因此, 剑潭梯级上下游的水环境质量对东深供水水质的影响较大。研究表明[21]剑潭水库富营养化程度已较蓄水初期有加剧趋势, 已有潜在的藻类水华风险, 且水流流速是影响东江干流浮游植物生长的重要因素[22]。为掌握流速对东江藻类水华发生的影响, 本研究采集东江惠州河段原水开展藻类室内培养模拟试验, 模拟东江水体中浮游藻类生长对流速的响应, 讨论不同流速梯度下浮游藻类的生长特征及其变化规律, 探讨抑制藻类水华暴发的临界流速, 为东江流域水华风险防控提供数据支撑。
2 材料与方法
2.1 实验装置及流速设定
本研究采用5个环形有机玻璃水槽(图1), 水槽长宽高尺寸为1150 mm×650 mm×500 mm。将水槽放置于25 ℃恒温室内, 光照为日光灯照射, 水槽上方光照强度1500 lx。根据2015年3月12日在东江剑潭库区(坝首至库尾)现场实测的流速大小0.077—0.15 m·s–1以及东江的水流特征, 设置5个流速梯度(1#、2#、3#、4#和5#), 分别为0、0.045、0.075、0.135和0.25 m·s–1, 流速大小通过循环水真空泵调节控制。
2.2 实验材料
实验藻种选取蓝藻门的铜绿微囊藻(FACHB-905)、绿藻门的斜生栅藻(FACHB-416)和硅藻门的小环藻sp.(FACHB-1635), 以上藻种均购自中国科学院淡水藻种库。
2.3 实验方法
(1)实验用水取自东江剑潭水库大坝处的原水, 用25#浮游植物网滤去大型浮游动物, 再用0.45 µm醋酸纤维滤膜进行抽滤后, 分别注入5个水槽, 水槽中水深为30 cm, 水量170 L。测定水槽中水体理化指标初始值, 根据水体氮磷营养盐浓度, 向水槽中添加浓度为15 g·L–1的NaNO3和3 g·L–1的KH2PO4, 用于调节各水槽水体中总氮、总磷浓度, 分别达到3.0 mg·L–1和0.19 mg·L–1。
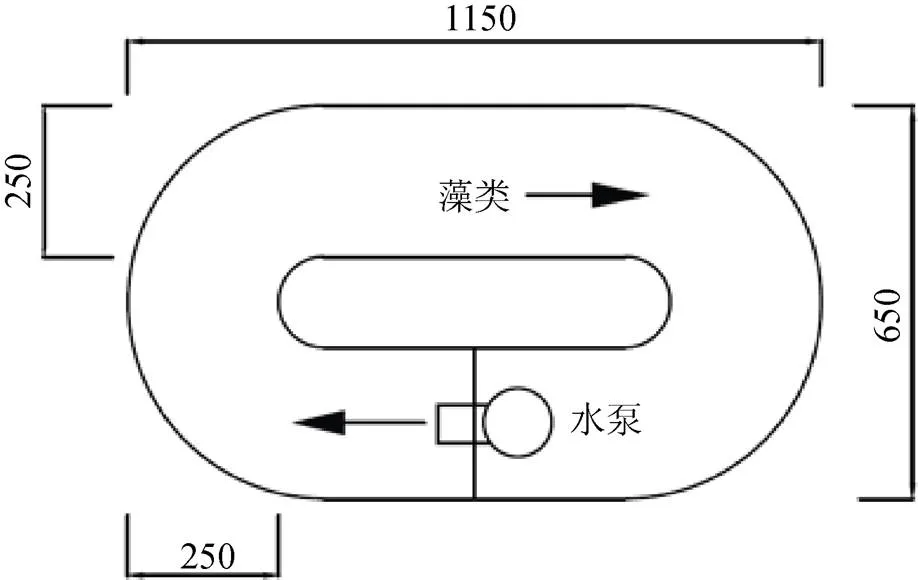
图1 实验装置平面示意图 (单位mm)
(2)将室内培养至对数生长期的藻种, 收集置于50 mL离心管, 离心收获藻液, 用15 mg·L–1NaCO3溶液洗涤后再次离心, 弃去上清液, 重复两次, 以除去附着在藻细胞上的营养盐, 预处理后转接至实验水槽内。
(3)实验期间隔天测量氮、磷营养盐浓度, 保证装置中氮、磷营养盐浓度维持在高富营养水平, 每天早上(9:00)和晚上(21:00)分别监测1次, 记录溶解氧(DO)、pH等, 并测量藻细胞密度、叶绿素a和叶绿素荧光参数(Fm、Fv)直到实验结束。
2.4 样品分析
pH和DO用便携式水质分析仪测定, 总氮(TN)、总磷(TP)和叶绿素a的测定均参考《水和废水监测方法(第4版)》, 显微镜(OLYMPUS CX21)下浮游藻类细胞数量计数方法参照《淡水浮游生物研究方法》[23], 叶绿素荧光参数用浮游植物荧光仪(WATER-PAM)测定, Fv/Fm是可变荧光与最大荧光的比值, 反映藻类PSII的活性。
2.4 数据处理
最大比增长速率()是在某一时间间隔内藻类生长的速率。其计算公式为:
=ln (X/X)/(t-t) (1)
式中:X为某一时间间隔开始时的藻类现存量(cells·mL–1);X为某一时间间隔结束时的藻类现存量(cells·mL–1);t-t: 为某一时间间隔(d)。
使用Origin Lab8.5对试验数据进行图形分析;使用SPSS17.0统计软件进行方差分析(ANOVA), 显著性水平为0.05。
3 结果与分析
3.1 流速对不同藻类生长过程中pH和DO的影响
不同流速条件下铜绿微囊藻、斜生栅藻和小环藻生长过程中pH的变化整体均呈现上升趋势, 其中铜绿微囊藻和斜生栅藻的pH值变化范围较接近, 分别为6.82—7.83、6.66—7.69; 小环藻的pH值相对较高, 变化范围为7.21—8.24(图2)。流速组(2#—5#)铜绿微囊藻生长期内pH均高于静止组(1#); 斜生栅藻和小环藻生长的水体中, 有流速的条件下随着流速增加pH值升高, 而静止组pH波动明显, 在小环藻生长的中后期静止培养组pH值高于流速组。
不同流速条件下铜绿微囊藻生长过程中DO呈先上升后降低的变化趋势, 变化范围为7.45—8.42 mg·L–1, 流速组DO高于静止培养组(1#); 斜生栅藻和小环藻生长过程中DO变化呈上升趋势, 且在生长的中后期静止培养组的DO升高较快, 最高值分别达到9.53 mg·L–1和10.65 mg·L–1, 高于流速组(图2)。
3.2 流速对不同藻类生长速率的影响
不同流速条件下初始接种的铜绿微囊藻细胞密度平均值为6.29×105cells·L–1, 在生长过程中不同流速组铜绿微囊藻细胞密度呈现先上升后降低趋势(图3)。在流速为0.135 m·s–1和0.25 m·s–1条件下培养至第3 d时, 铜绿微囊藻细胞密度数量高于其他各流速组和静止组, 至培养的第4 d, 各流速组和静止组的铜绿微囊藻细胞密度数量均大幅下降。不同流速条件下铜绿微囊藻生长中叶绿素a浓度与藻细胞密度变化趋势较为一致, 至培养的第4 d, 叶绿素a浓度已由第1 d的平均值3.42 µg·L–1下降为1.18 µg·L–1。结合显微镜下观察, 以上结果反映出铜绿微囊藻在培养过程中生长状况较差, 至培养的第4 d藻细胞开始大量死亡。

图2 不同流速下pH、DO变化趋势
不同流速条件下初始接种的斜生栅藻细胞密度平均值为0.72×105cells·L–1, 斜生栅藻在生长过程中静止组的细胞密度和叶绿素a浓度均高于流速组, 最低流速0.045 m·s–1条件下藻细胞密度和叶绿素a浓度一直处于较低水平, 其他各流速组藻细胞密度和叶绿素a浓度波动较大(图3)。
不同流速条件下初始接种的小环藻细胞密度平均值为2.31×105cells·L–1, 在生长的第5 d小环藻生长开始进入稳定期, 且该时期0.135 m·s–1和0.25 m·s–1高流速组的藻细胞密度和叶绿素a浓度低于0.045 m·s–1、0.075 m·s–1流速组和静止培养组(图3)。
3.3 流速对藻类最大比增长速率的影响
不同流速条件下铜绿微囊藻、斜生栅藻和小环藻的最大比增长率如图4所示, 铜绿微囊藻静止组与流速组最大比增长率呈现明显差异(<0.05), 静止组在生长过程中细胞密度很快下降至低于初始接种密度水平, 导致最大比增长速率为负值, 流速组最大比增长率相对较低且呈相同的变化趋势。斜生栅藻在流速(≤0.045)和流速(≥0.075)呈现明显差异(<0.05), 且静止组的最大比增长速率最高, 流速由0.045 m·s–1增加至0.075 m·s–1时, 斜生栅藻最大比增长速率下降幅度较大, 说明流速的增加对斜生栅藻的生长产生一定的抑制作用。小环藻的最大比增长率在不同流速组差异不明显, 且在流速为0.075 m·s–1时, 最大比增长率相对较高, 流速>0.075 m·s–1时最大比增长速率下降, 说明高流速也不利于小环藻的生长。
3.4 流速对不同藻类叶绿素荧光参数Fv/Fm的影响
不同流速下铜绿微囊藻、斜生栅藻、小环藻的Fv/Fm值变化如图5所示。铜绿微囊藻初始Fv/Fm值约为0.3, 在生长期内铜绿微囊藻的Fv/Fm值逐渐降低接近为零, 说明水体中铜绿微囊藻光合系统PSII活性很低。在生长期内不同流速条件下的斜生栅藻Fv/Fm均逐渐降低, 小环藻Fv/Fm则呈现先上升至稳定然后又下降的趋势, 且静止组(1#)斜生栅藻和小环藻的Fv/Fm均高于流速组(2#—5#)。
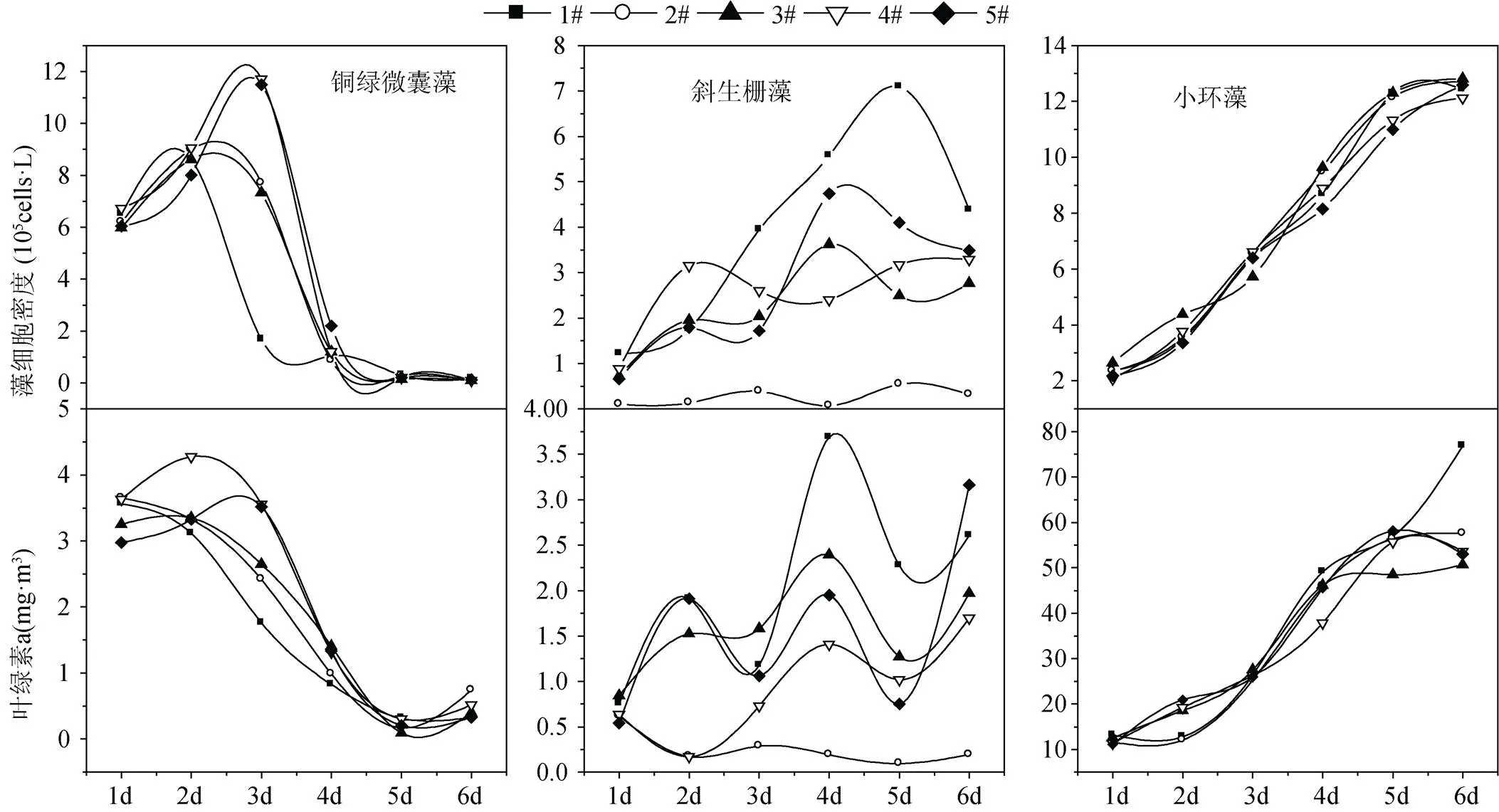
图3 不同流速下藻细胞密度和叶绿素a的变化情况

注:字母a, b标识最大比增长率在不同流速之间显著性差异比较, 采用SPSS17.0进行方差分析, 显著水平0.05。

图5 不同流速下藻类Fv/Fm的变化趋势
4 讨论
4.1 东江水体发生藻类水华的风险分析
东江作为流域内河源、惠州、东莞、深圳、广州等城市以及香港约4000多万人口的主要饮用水源, 其水环境保护一直备受重视, 流域内采取了系列严格的水环境保护措施, 尤其是东江流域上游, 为保护好东江优质水资源在经济发展方面做出了巨大的牺牲[24]。尽管东江干流水质整体优良, 然而由于流域内人口增加和经济的快速发展[25], 干流仍有少数区域达到富营养化水平[18]; 东江干流TN含量均值为1.99 mg·L–1, TP含量变化范围为0.01—0.28 mg·L–1, 且越靠近下游TN、TP含量越高, 水体富营养化程度越高[22], 特别是东江下游剑潭水库内较高的营养盐已为浮游植物大量生长繁殖创造了有利条件, 东江下游潜在的水华风险受到关注[21]。
近年来我国河流藻类水华发生呈增加趋势, 与湖泊水华相比, 水动力条件的改变是河流中浮游藻类优势种变化的最主要影响要素, 也是河流中影响浮游藻类生长的主要限制因素之一[26]。不同藻类的临界流速不同[11], 澳大利亚达令河卷曲鱼腥藻大量暴发的临界流速为 0.05 m·s–1[14]; 运用非恒定二维生态动力学模型研究发现嘉陵江藻类水华发生的临界流速为0.04 m·s–1[27]; 室外试验研究发现小球藻和纤维藻生长的临界流速为0.05 m·s–1和0.01 m·s–1[28]; 室内实验表明低流速(0.05—0.12 m·s–1)有利于铜绿微囊藻生长, 而在静止与高流速条件下 (0.20—0.30 m·s–1)藻类的生长受到抑制[29]。除嫌水藻类外, 中流藻类和喜流水藻类都可以在流水中生长, 由于水体流动使藻类不断得到新的营养物质的供应, 水流对藻类的生长和繁殖是有利的[30]。微囊藻属嫌流水藻类[31], 较高的流速对微囊藻的生长有明显的抑制作用, 但较缓慢的流速在一定程度上有利于微囊藻的生长。本研究发现在静水、小流速和较大流速条件下, 铜绿微囊藻生长速率均较低, 与实验所用的铜绿微囊藻活性较低有关, 实验初始铜绿微囊藻的Fv/Fm仅为0.263—0.321; 其次, 研究发现东江剑潭水库中微囊藻并不是优势种, 其藻细胞密度较低[21];此外, 根据实验氮磷营养盐浓度水平, 各装置在实验第2d氮磷浓度有所降低, 说明微囊藻生长消耗了营养盐, 而从第3d开始, 氮磷浓度基本保持不变, 可见水体中微囊藻已不再消耗营养盐。以上结果表明目前东江干流剑潭水库中并不适宜微囊藻的大量生长, 由此可以判断东江剑潭水库发生微囊藻水华的可能性较低。斜生栅藻作为绿藻的一种, 以小流速下生长状况最好, 而较高的流速不利于其生长。斜生栅藻在流速大于0.02 m·s–1时即产生明显的抑制生长作用[33]。硅藻是形成河流水华的主要种类之一, 多发生在天然河流成库后的淹没回缓流水库湾或支流。小环藻为喜流水藻类, 适宜生活于较低的水流速度, 而对较高的水流速度耐受较弱[31]。近年来汉江暴发硅藻水华(优势种为小环藻)时的河道流速为0.07 m·s–1左右[32]。本研究发现小环藻在流速为0.075 m·s–1时最大比增长速率较高, 在相对较高的流速(0.135和0.25 m·s–1)时, 小环藻的生长开始出现抑制, 说明当东江干流剑潭水库水流大于0.075 m·s–1时, 随着流速的增加小环藻生长受到抑制; 斜生栅藻也是在流速大于0.075 m·s–1时最大比增长速率有所降低, 以上结果表明剑潭水库水体流速增加至0.075 m·s–1以上将对藻类生长产生抑制作用。
4.2 东江水体水华防控措施
作为快速有效改善水库型河流和水库库区水环境的手段之一, 通过改善水利条件从而减少水华暴发风险的研究越来越受到重视。Maier等[12]研究发现通过水利调节打破水体热分层、减少水体滞留时间, 可以消除澳大利亚Darling河的鱼腥藻水华; 辛小康等[35]通过建立平面二维水动力数学模型发现, 利用干支流水库对香溪河库湾流速的联合调控可适度控制三峡库区支流库湾水华的暴发; 殷大聪等[36]研究提出汉江中下游春季硅藻水华暴发的关键水文阈值及生态调度控制策略; 刘德富等[37]研究提出用于防控三峡支流水华的 “潮汐式”生态调度方案。东江作为香港、深圳及广州等特大城市的重要水源地, 水库调度是有效解决流域突发性水污染的重要措施, 同时由于新丰江、枫树坝与白盆珠三大水库对东江全流域水资源有控制性调蓄作用, 东江具有开展水利调度的有利条件, 已有研究[20,38]表明利用东江水利枢纽进行水量调度可有效改善东江干流水质。本研究结果发现铜绿微囊藻在东江水体中长势较弱, 发生水华的可能性较低, 而斜生栅藻和小环藻均在较低流速<0.075 m·s–1时生长速度快, 且随着流速增大, 对藻类生长有促进作用, 容易发生水华, 而在较高流速>0.075 m·s–1条件下, 随着流速的增加, 藻类的生长受到抑制。因此, 针对当前枯水期东江存在水华的风险, 及实验结果东江水华发生临界流速为0.075 m·s–1, 并结合东江惠州段现场实测流速, 确定东江剑潭水库防控水华的临界流速为0.07—0.08 m·s–1。在水华易发的气象条件下, 可通过流域内水库联合调度以保证东江的生态流速, 从而有效防止水华的发生。
5 结论
(1)实验中斜生栅藻临界流速值约为0.075 m·s–1; 铜绿微囊藻在不同流速条件下生长都较弱; 小环藻对流速响应关系为在流速>0.075 m·s–1时藻类生长受到抑制。因此, 东江流域惠州剑潭水库大坝处浮游藻类生长的临界流速为0.075 m·s–1。
(2)实验中不同藻类对流速的响应适应性具有明显差异, 并且流速条件的改变对藻类的生物量和生理生化特征产生较大影响。因此, 若要通过水利调度来预防和控制水华, 需视具体藻类来优化水利调度方案。
[1] SMITH V H. Eutrophication of freshwater and coastal marine ecosystems a global problem [J]. Environmental Science and Pollution Research, 2003, 10(2): 126–39.
[2] MORSE R E, SHEN J, BLANCO-GARCIA J L, et al. Environmental and physical controls on the formation and transport of blooms of the DinoflagellateMargalef in the lower Chesapeake Bay and its tributaries [J]. Estuaries & Coasts, 2011, 34(5): 1006–1025.
[3] HA K, JANG M H, JOO G J. Winterbloom development in the Nakdong River regulated by an estuary dam and tributaries [J]. Hydrobiologia, 2003, 506-509(1-3): 221–227.
[4] 张远, 夏瑞, 张孟衡, 等.水利工程背景下河流水华暴发成因分析及模拟研究[J].环境科学研究, 2017, 30(8): 1163–1173.
[5] LIU Xia, LI Yunliang, LIU Baogui, et al. Cyanobacteria in the complex river-connected Poyang Lake: horizontal distribution and transport [J]. Hydrobiologia, 2016, 768(1): 95–110.
[6] 程辉, 吴胜军, 王小晓, 等.三峡库区生态环境效应研究进展[J].中国生态农业学报, 2015, 23(2): 127–140.
[7] 梁培瑜, 王烜, 马芳冰.水动力条件对水体富营养化的影响[J].湖泊科学, 2013, 25(4): 455–462.
[8] ZHOU Guangjie, ZHAO Xuemin, BI Yonghong, et al. Phytoplankton variation and its relationship with the environment in Xiangxi Bay in spring after damming of the Three-Gorges, China [J]. Environmental Monitoring & Assessment, 2011, 176: 125–141.
[9] 边归国, 陈宁, 胡征宇, 等.福建某河流甲藻水华与污染指标的关系[J].湖泊科学, 2010, 22(3): 405–410.
[10] 潘晓洁, 朱爱民, 郑志伟, 等.汉江中下游春季浮游植物群落结构特征及其影响因素[J].生态学杂志, 2014, 33(1): 33–40
[11] 吴晓辉, 李其军.水动力条件对藻类影响的研究进展[J].生态环境学报, 2010, 19(7): 1732–1738.
[12] MAIER H R, BURCH, M D, BORMANS M. Flow management strategies to control blooms of the cyanobacterium,, in the river Murray at Morgan, South Australia [J]. Regulated Rivers Research & Management, 2001, 17(6): 637–650.
[13] ACUNA V, VILCHES C, GIORGI A. As productive and slow as a stream can be——the metabolism of a Pampean stream [J]. Journal of the North American Benthological Society, 2011, 30(1): 71–83.
[14] MITROVIC S M, OLIVER R L, REES C, et al. Critical flow velocities for the growth and dominance ofin some turbid freshwater rivers [J]. Freshwater Biology, 2003 48(1): 164–174.
[15] 陈瑞弘, 李飞鹏, 张海平, 等.面向流量管理的水动力对淡水藻类影响的概念机制[J].湖泊科学, 2015, 27(1): 24–30.
[16] MITROVIC S M, HARDWICK L, DORANI F. Use of flow management to mitigate cyanobacterial blooms in the lower Darling River, Australia [J]. Journal of Plankton Research, 2011, 33(2), 229–241.
[17] YAN Liang, FUNG Puika, MAN Fungtse, et al. Sources and seasonal variation of PAHs in the sediments of drinking water reservoirs in Hong Kong and the Dongjiang River (China) [J]. Environmental Monitoring & Assessment, 2008, 146: 41–50.
[18] 王博, 梁志诚, 任斐鹏, 等.东江干流富营养化评价及区域差异性研究[J].环境污染与防治, 2011, 33(6), 15–18.
[19] 廖剑宇, 彭秋志, 郑楚涛, 等.东江干支流水体氮素的时空变化特征[J].资源科学, 2013, 35(3): 505–513.
[20] 江涛, 钟鸣, 邹隆建, 等.石马河泄洪与东江水利枢纽调节不同情景下东江水质的模拟与分析[J].中山大学学报(自然科学版), 2016, 55(2): 117–123.
[21] 胡芳, 许振成, 姚玲爱, 等.剑潭水库浮游植物群落特征与水环境因子关系研究[J].环境科学学报, 2014, 34(4): 950–958.
[22] 田时弥, 杨扬, 乔永民, 等.珠江流域东江干流浮游植物叶绿素a时空分布及与环境因子的关系[J].湖泊科学, 2015, 27(1): 31–37.
[23] 章宗涉, 黄祥飞.淡水浮游生物研究方法[M].北京: 科学出版社, 1991.
[24] 刘强, 彭晓春, 周丽旋, 等.城市饮用水水源地生态补偿标准测算与资金分配研究——以广东省东江流域为例[J]. 生态经济, 2012, (1): 33–37.
[25] GAO Chanjun, ZHOU Ping, JIA Peng, et al. Spatial driving forces of dominant land use/land cover transformations in the Dongjiang River watershed, Southern China [J]. Environmental Monitoring & Assessment, 2016, 188(2): 84.
[26] 陈永灿, 俞茜, 朱德军, 等.河流中浮游藻类生长的可能影响因素研究进展与展望[J].水力发电学报, 2014, 33(4): 186–195.
[27] LONG Tianyu, WU Lei, MENG Guohu, et al. Numerical simulation for impacts of hydrodynamic conditions on algae growth in Chongqing Section of Jialing River, China [J]. Ecological Modelling, 2011, 222(1): 112–119.
[28] 焦世珺.三峡库区低流速河段流速对藻类生长的影响[D].重庆: 西南大学, 2007.
[29] 王华, 逄勇.藻类生长的水动力学因素影响与数值仿真[J].环境科学, 2008, 29(4): 884–889.
[30] 福迪. 藻类学[M].罗迪安, 译.上海: 上海科学技术出版社, 1980: 392–394.
[31] 杨敏, 毕永红, 艾鹰, 等.人工控制条件下水流速对香溪河库湾浮游植物影响的初步研究[J]. 长江流域资源与环境, 2012, 21(2): 220–224.
[32] 卢大远, 刘培刚, 范天俞, 等.汉江下游突发“水华”的调查研究[J].环境科学研究, 2000, 13(2): 28–31.
[33] 王建慧.流速对藻类生长影响试验及应用研究[D].北京: 清华大学, 2012.
[34] 边归国.九龙江拟多甲藻水华预警和应急处置的初步研究[J].中国环境管理, 2013, 5(3): 30–34.
[35] 辛小康, 尹炜, 叶闽.水动力调控三峡库区支流水华方案初步研究[J].水电能源科学, 2011, 29(7): 16–18.
[36] 殷大聪, 尹正杰, 杨春花, 等.控制汉江中下游春季硅藻水华的关键水文阈值及调度策略[J].中国水利, 2017, (9): 31–34.
[37] 刘德富, 杨正健, 纪道斌, 等.三峡水库支流水华机理及其调控技术研究进展[J].水利学报, 2016(3): 443–454.
[38] 黄国如, 李春.东江干流突发性重金属污染的水库调度效应研究[J].水电能源科学, 2013(6): 82–85.
The effect of flow velocity on the growth of different phytoplankton
ZHOU Jing1,2, GOU Ting2, ZHANG Luohong1, LAN Yu3, MA Qianli2, LIANG Rongchang2, ZHAO Xuemin2,*
1. School of Environmental and Chemical, Xi’an Polytechnic University, Xi’an 710048, China 2. South China Institute of Environmental Sciences, Ministry of Environmental Protection, Guangzhou 510655, China 3. College of Life Science and Technology, Jinan University, Guangzhou 510632, China
To study the effect of flow velocity on the growth of different phytoplankton, a simulation experiment was conducted in the annular flume of organic glass using,andsp., which were cultured in laboratory using surface water from the Huizhou reach of Dongjiang River in March 2015. The effects of different flow velocity on the algal cell density, the concentration of chlorophyll, the maximum specific growth rate and chlorophyll fluorescence parameter (Fv/Fm) were studied by adjusting the flow rate of water. The results showed that the physiological and biochemical characteristics varied at different flow rates. The growth ofwas slow with all flow velocity conditions, the growth ofandsp. was fast at low flow rate (<0.075m·s–1) and increased with the increase of flow velocity. However, under the condition of high velocity (<0.075m·s–1), the growth of algae was inhibited with the increase of flow velocity. The different continuous flow may directly induce the changes of the cell density and physiological indicators of phytoplankton, indicating the control of eutrophication and algae bloom by adjusting flow velocity in Dongjiang water.
Dongjiang River; flow velocity; phytoplankton; eutrophication; algal bloom
10.14108/j.cnki.1008-8873.2018.06.010
X522
A
1008-8873(2018)06-075-08
2018-06-11;
2018-08-02
广州市科技计划产学研协同创新重大专项(201704030110); 中央级公益性科研院所基本科研业务费专项(PM-zx703-201602-048); 国家水体污染控制与治理科技重大专项(2014ZX07206-005-04)联合资助
周静(1992—), 女, 硕士研究生, 主要从事水环境污染防治, E-mail:1602111799@qq.com
赵学敏, 博士, 研究员, 主要从事水生态风险评估与控制研究, Email: zhaoxuemin@scies.org
周静, 苟婷, 张洛红, 等. 流速对不同浮游藻类的生长影响研究[J]. 生态科学, 2018, 37(6): 75-82.
ZHOU Jing, GOU Ting, ZHANG Luohong, et al. The effect of flow velocity on the growth of different phytoplankton[J]. Ecological Science, 2018, 37(6): 75-82.
