NEDDylation参与乙型肝炎病毒前基因组RNA的包装
2018-11-14李程张境王勇翔
李程 张境 王勇翔
200032上海,复旦大学基础医学院病原生物学系医学分子病毒学教育部/卫生部重点实验室
乙型肝炎病毒(hepatitis B virus,HBV)感染是世界范围的健康问题。据估计全球约有2.5亿人罹患慢性乙型肝炎[1]。目前治疗慢性乙肝的抗病毒药物有干扰素α、PEG化干扰素α和核(苷)酸类似物等[2]。干扰素治疗具有明显的副作用;核苷酸类似物治疗因停药后出现病毒复制反弹而需长期用药,有病毒耐药的风险[3]。因此,迫切需要开发新的抗HBV制剂。
HBV通过结合钠离子-牛磺胆酸共转运蛋白(sodium taurocholate co-transporting polypeptide,NTCP)受体进入肝细胞,核衣壳被运输到细胞核。在细胞核内,核衣壳内的松弛环状 DNA(relaxed circular DNA,rcDNA)被修复成共价闭合环状DNA(covalently closed circular,cccDNA)。 以cccDNA为模板,细胞RNA聚合酶II转录出5种HBV RNA,包括3.5 Kb编码HBe抗原前体的precore RNA和编码核心蛋白和聚合酶的前基因组RNA(pregenomic RNA,pgRNA)、2.4 Kb编码 L蛋白的 mRNA、2.1 Kb编码M和S蛋白的mRNA,0.7 Kb的编码X蛋白的mRNA。HBV聚合酶结合pgRNA5’端茎环结构,同时招募核心蛋白二聚体包裹pgRNA成核衣壳[4]。在核衣壳内,HBV聚合酶以pgRNA为模板逆转录合成负链 DNA后,通过 RNaseH活性将pgRNA/负链DNA双链中的pgRNA降解,再以负链DNA为模板合成正链DNA。核衣壳在多囊泡体出芽并释放出细胞[5]。
NEDD8是与泛素类似的蛋白。NEDD8在其活化酶(NEDD8 activating enzyme, NAE/E1)、结合酶(E2),连接酶(E3)的催化下,将其羧基端第76位的甘氨酸与底物的赖氨酸共价结合,这一过程叫做NEDDylation。NEDDylation的底物主要是含 cullin的泛素连接酶(cullin-RING liagses,CRLs)[6],参与调节泛素-蛋白酶体系统(ubiquitin-proteasome system,UPS)从而参与调节细胞信号转导、细胞周期、细胞生长及凋亡。近年来研究表明NEDDylation还与病毒复制有关。疱疹病毒、腺病毒和流感病毒通过NEDDylation激活CRLs而促进病毒复制[7-8];一些RNA病毒(如仙台病毒、水疱性口炎病毒等)通过 CRLs相关的 UPS促进干扰素调节蛋白-3(interferon regulatory factor 3,IRF3)降解而干扰抗病毒先天性免疫反应[9-10]。近年来研究还发现存在不依赖CRLs的NEDDylation[11],提示NEDD8可能具有额外的生物学功能。目前关于细胞NEDDylation在HBV复制中的作用仍不清楚。本研究采用NAE特异性抑制剂 MLN4924抑制细胞CRLs和其他一些底物的NEDDylation,检测HBV复制和HBV pgRNA包装效率。
1 材料与方法
1.1 材料 pCH-9/3091质粒由德国弗赖堡大学Michael Nassal教授提供,包含D基因型的HBV基因组(GenBank登记号V01460)。为检测包装入核衣壳的pgRNA,本研究构建了HBV聚合酶失活变异体表达质粒pCH-9/3091-YMHD:以pCH-9/3091质粒为基础,将聚合酶催化中心YMDD基序基因突变为YMHD基因而获得HBV聚合酶失活变异体pCH-9/3091-YMHD,将该质粒转染Huh-7细胞后,pgRNA可被包装入核衣壳,但不能合成负链DNA,pgRNA也不被降解,因此核衣壳内的HBV核酸为pgRNA。将 HBV核心蛋白基因扩增后克隆入pCDNA6载体获得pCDNA-core质粒,将该质粒转染Huh-7细胞后,细胞虽然产生了大量核心颗粒,但用HBV特异的探针作杂交发现这些颗粒不包装任何HBV特异的核酸,用作对照。Huh-7细胞和四环素调控且稳定表达HBV的HepAD38细胞为本室保存。MLN4924购自 MedChemExpress公司,胎牛血清购自Gibco公司,DMEM培养液和核酸预杂交液购于ThermoFisher Scientific公司,随机引物DNA标记试剂盒购自Roche公司,α-32P-dGTP购于Perkin-Elmer公司。实验中所用抗体及来源如下:anti-core多克隆抗体(Dako公司),anti-core单克隆抗体由张继明教授(复旦大学附属华山医院)馈赠,actin单克隆抗体和gpr78多克隆抗体(Proteintech公司),辣根过氧化物酶(horseradish peroxidase,HRP)标记的二抗(Jackson ImmunoResearch公司)。
1.2 方法
1.2.1 细胞培养:Huh-7细胞用含10%胎牛血清、100 U/ml青霉素、100 U/ml链霉素的DMEM培养基,于37℃、5%CO2恒温恒湿培养箱中培养,每1~2 d进行细胞传代。HepAD38细胞在上述培养基中额外添加200μg/ml G418抗生素,在相同培养条件下生长,不含四环素的培养液培养使pgRNA转录而起始HBV复制,每2~3 d传代。
1.2.2 DNA转染:计数3.0×105个Huh-7细胞铺于6孔板中,18~24 h后进行转染。取250μl opti-MEM,加入质粒 DNA 2.5μg,混匀后加入7.5μl TransIT-LT1转染试剂,混匀后静置15 min。将混合物均匀滴加到已更换新鲜培养液的6孔板中。
1.2.3 药物处理:向HepAD38细胞培养上清中加入梯度浓度的MLN4924,并用不同浓度的HBV聚合酶抑制剂 LAM作对照,每 3 d更换1次含MLN4924或LAM的培养基,加药后第6天用NP40裂解缓冲液裂解细胞;向转染了pCH-9/3091-YMHD的Huh-7细胞中添加MLN4924(终浓度为0.1 μmol/L),并添加DMSO作为对照(MLN4924溶于DMSO),每3 d更换1次含MLN4924或DMSO的培养基,加药后第6天裂解细胞。
1.2.4 细胞内病毒核衣壳检测:利用非变性琼脂糖凝胶电泳(native agarose gel electrophoresis,NAGE)分离HBV核衣壳:NP40裂解液裂解细胞,细胞膜、细胞器膜和HBV包膜破裂,核衣壳释放出来。裂解液经1%的琼脂糖凝胶分离,衣壳颗粒在电场的作用下向正极迁移。随后用免疫印迹检测核衣壳:通过虹吸转膜装置将衣壳颗粒转移至尼龙膜上,5%脱脂奶粉封闭后与anti-core多克隆抗体孵育,TBST漂洗后与 HRP标记的二抗孵育,用化学发光液(chemiluminescence, ECL)检测。
1.2.5 十二烷基硫酸钠-聚丙烯酰氨凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)和免疫印迹:细胞样品用含SDS的样品缓冲液处理,100℃变性10 min。配制4.5%浓缩胶和12.5%分离胶进行电泳。电泳后将蛋白转移到PVDF膜上,按照1.1.3所述方法检测蛋白。
1.2.6 细胞内HBV DNA提取:取一定量细胞裂解上清加入DNase I 37℃消化处理30 min,再加入蛋白酶K 53℃消化释放病毒基因组,最后经酚/氯仿抽提和DNA沉淀获得HBV DNA。
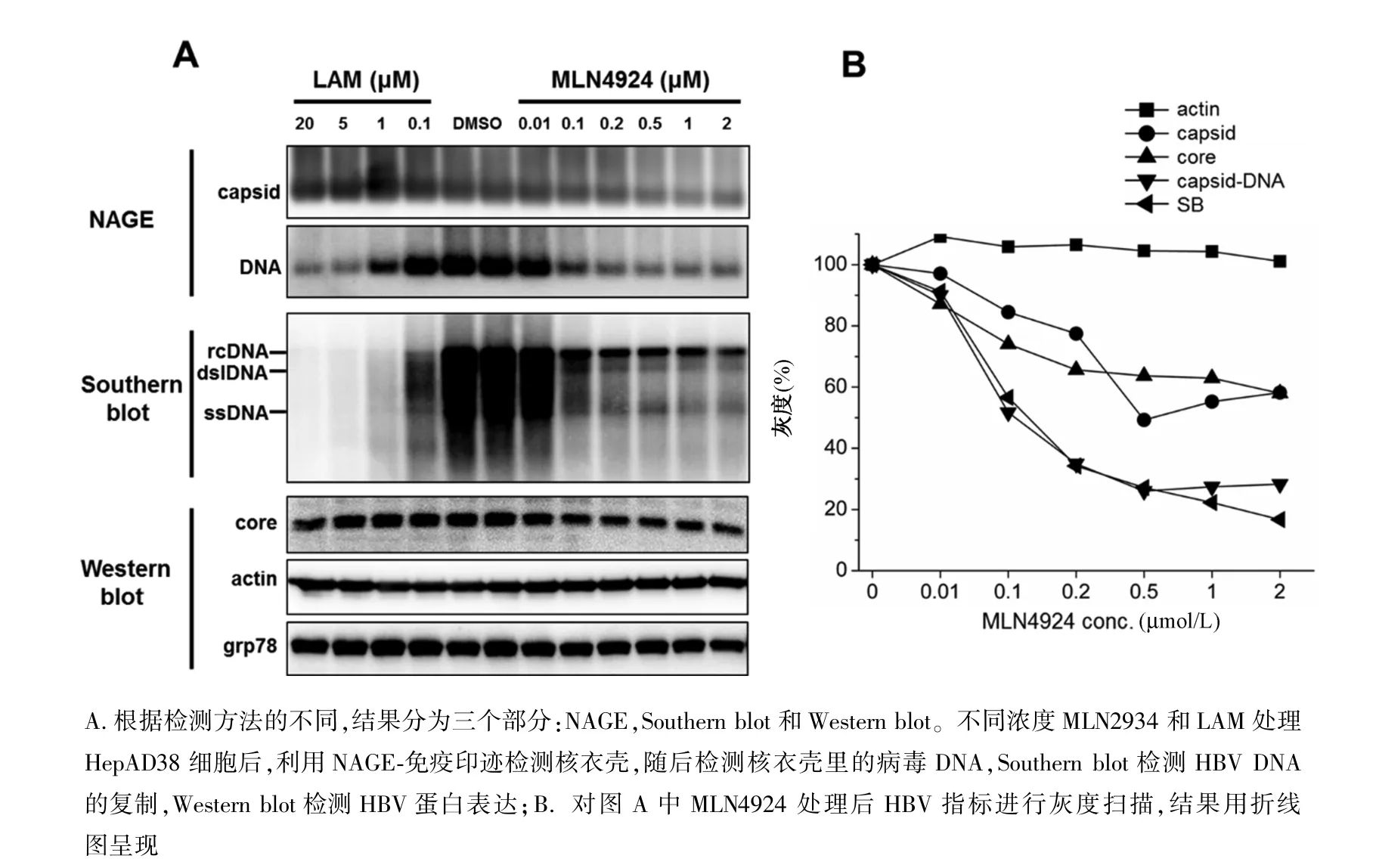
图1 MLN4924剂量依赖地抑制HepAD38细胞HBV DNA复制A.According to the different detection methods, the results were divided into three parts:NAGE, Southern blot and Western blot.HepAD38 Cells were incubated with various concentrations of MLN4924 and lamivudine for 6 days.Intracellular capsids were analyzed by NAGE, capsid associated viral DNAs by Southern blot, and viral and cellular proteins by Western blot;B.Graphic presentation of HBV replication marker levels at the treatment of increasing concentrations of MLN4924Fig.1 MLN4924 inhibited HBV DNA replication in a dose-dependent manner in HepAD38 cells
1.2.7 Southern blot检测病毒 DNA复制:HBV DNA样品经1%的NAGE分离;变性液和中和液分别处理凝胶45 min后,通过虹吸转膜装置将病毒DNA转移至尼龙膜上;尼龙膜80℃烤膜固定;预杂交液42℃孵育30 min;用随机引物DNA标记试剂盒标记探针;将变性后的掺入32P标记的dGTP的DNA探针加入预杂交液中42℃杂交过夜;洗膜后磷屏曝光,最后用多功能生物分子成像仪扫描磷屏记录的信号并成像。
1.2.8 原位检测核衣壳中DNA:1.2.4中经NAGE分离和免疫印迹检测的核衣壳在80℃烤箱中烘烤2 h,使衣壳颗粒固定在尼龙膜上,DNA变性液处理5 min,中和液处理5 min;再用1.2.6所述方法检测颗粒内的病毒DNA。
1.2.9 原位检测核衣壳中RNA:相关溶液均用DEPC水配制,检测方法同1.2.7。
1.2.10 数据处理:利用 FUJIFILM公司的 Multi Gauge V2.2软件对蛋白、DNA及RNA信号进行灰度扫描;利用数据处理软件Origin 8.0进行折线图的绘制;两组数据均数的比较采用Student-t检验,P<0.05具有统计学意义。
2 结果
2.1 MLN4 924剂量依赖地抑制HBV DNA复制Western blot结果显示0.1~20μmol/L LAM和0.01~2μmol/L MLN4924对actin和GRP78蛋白的表达无明显影响(图1A,Western blot),提示高达20 μmol/L拉米夫定和2μmol/L MLN4924不引起细胞毒性。Western blot检测细胞内核心蛋白,NAGE-免疫印迹检测细胞内核衣壳,结果显示LAM不改变核心蛋白的表达和核衣壳的形成,而MLN4924能轻度剂量依赖地减少核心蛋白和核衣壳(图1A,Western blot和NAGE)。Southern blot和原位病毒DNA检测结果显示LAM和MLN4924都能显著地且剂量依赖地减少核衣壳相关病毒DNA(图1,Southern blot和NAGE)。灰度扫描结果显示,随着MLN4924浓度增高,actin的量无明显变化,核心蛋白和核衣壳变化趋势一致,Southern blot和NAGE原位检测病毒DNA结果一致(图1B,Southern blot和NAGE),且随着MLN4924浓度增加,核衣壳中病毒DNA下降幅度显著高于核衣壳(图1A,capsid和DNA;图1B),这提示HBV pgRNA包装或下游DNA合成可能被抑制。
2.2 MLN4924抑制HBV pgRNA包装 pCDNA-core转染Huh-7细胞后,细胞产生了大量核心颗粒(图2A,capsid),但HBV特异的探针作杂交结果显示颗粒中不包装任何HBV特异的核酸(图2A,pgRNA)。pCH-9/3091-YMHD转染Huh-7细胞后,细胞也产生了大量的核心颗粒(图2A,capsid),而且这些颗粒包装了pgRNA(图2A,pgRNA)。当用核心颗粒的量作校正后,发现0.1μmol/L MLN4924使核衣壳包装的pgRNA减少至DMSO对照组的53.4%(图2B),而相同浓度的MLN4924也使衣壳相关病毒 DNA减少至对照组的56.6%(图1B)。因此MLN4924可能通过抑制pgRNA包装而减弱HBV复制。
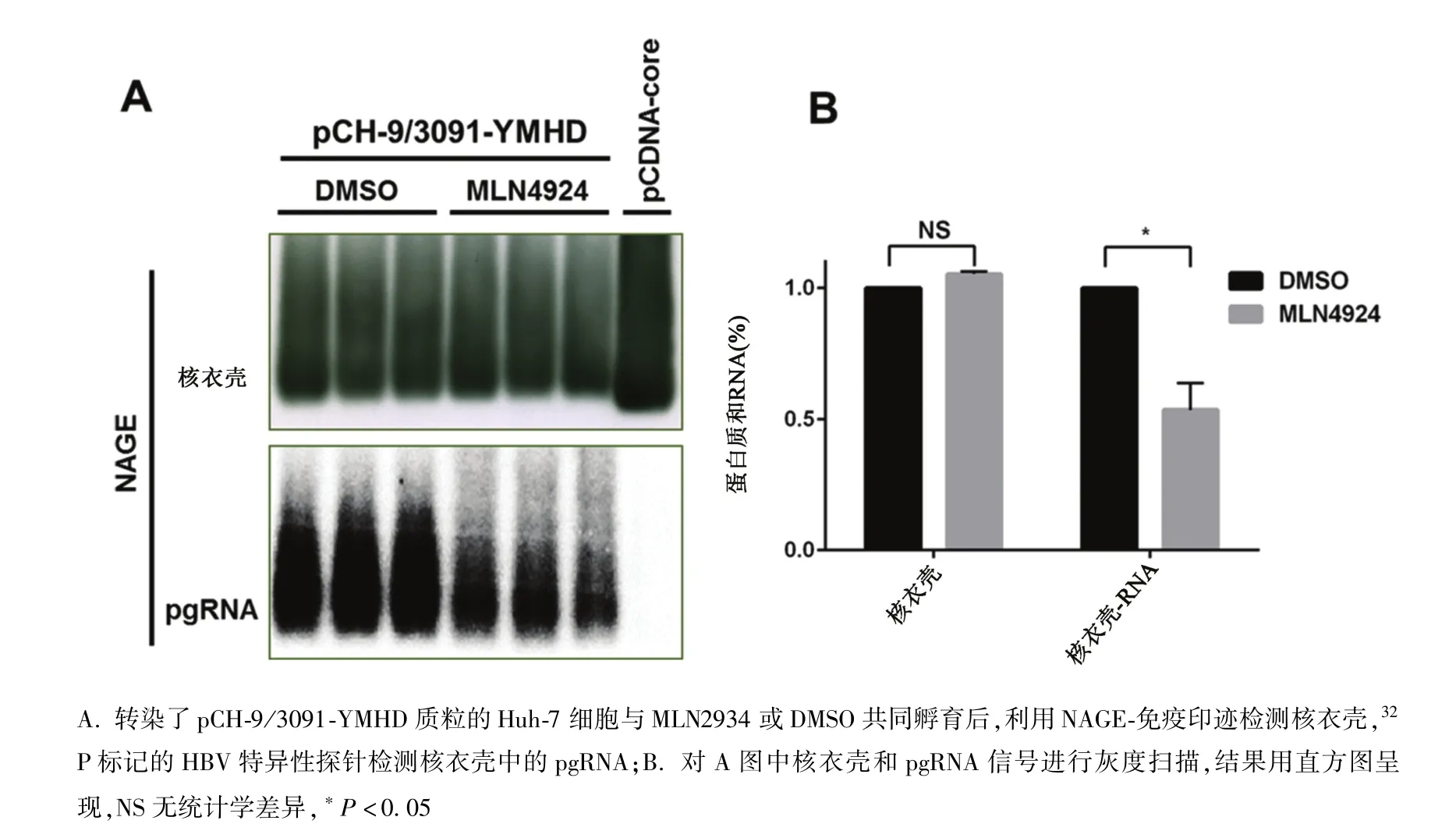
图2 MLN4924抑制HBV pgRNA的包装A.Huh-7 cells were respectively transfected with pCH-9/3091-YMHD or pCDNA-core plasmids,then incubated with MLN4924 or vehicle-DMSO.Cytoplasmic capsids and capsid-associated RNA were sequentially detected with NAGE-immunoblotting and hybridization with 32 P labeled HBV specific DNA probes;B.Graphic presentation shows that relative pgRNA encapsidation efficiency by normalizing the amount of packaged pgRNA with that of capsids.NS:Not significant, ∗P <0.05 Fig.2 MLN4924 inhibited pgRNA encapsidation
3 讨论
本研究采用NAE抑制剂-MLN4924干扰细胞内蛋白NEDDylation修饰反应。MLN4924轻度剂量依赖地抑制核心蛋白的表达。有研究发现NEDDylation修饰HBV X蛋白(HBx)后可阻止HBx通过泛素化途径降解[12],从而促进cccDNA转录激活[13]。NAE抑制剂MLN4924抑制HBx NEDDy lation,HBx降解增多,其促进cccDNA转录作用减弱,从而导致编码核心蛋白的pgRNA减少。
MLN4 924剂量依赖地减弱pgRNA包装和HBV DNA复制,而且两者下降程度相近。HBV DNA复制是 pgRNA包装入核衣壳的后续步骤,因此MLN4924通过抑制pgRNA包装而减弱HBV DNA复制。pgRNA包装是个复杂的过程。在宿主因子如HSP90等的帮助下,HBV聚合酶识别并结合pgRNA,再招募核心蛋白二聚体组装成核衣壳,如果NEDDylation修饰上述病毒或宿主蛋白,或修饰其他未知宿主因子而调节pgRNA包装,则像MLN4924这样的NEDDylation抑制剂将抑制pgRNA包装。因此,本研究还将检验 HBV聚合酶、核心蛋白和HSP90是否为NEDDylation的底物,这将有助于进一步了解HBV复制机理。
利益冲突 无
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
