文心兰25条miRNA前体序列特性及其表达分析
2018-11-06王培育王丛巧张舒婷王雪晶赖钟雄林玉玲
王培育,王丛巧,张舒婷,王雪晶,叶 炜,赖钟雄,林玉玲
(福建农林大学 园艺植物生物工程研究所,福州 350002)
microRNA(miRNA)是一种长度约为21~24 nt大小的内源性非编码单链RNA,其产生依赖于miRNA初级体primary miRNA(pri-miRNA)的存在[1-2]。miRNA主要通过与靶mRNA结合,在转录水平或转录后水平介导靶mRNA降解或翻译进行调控,对植物基因表达、生长发育和抵抗胁迫及病害有着十分重要的影响。在已报道的一系列受病原菌侵染负调控的miRNA中,miR393是第一个被发现的,它主要通过负调控TIR1/AFBs的表达,从而响应病原菌胁迫[3]。此外,miR393和miR167能够响应根癌农杆菌的胁迫[4-5];miR159前体获得能够增强抗病性[6];miR398能受病原菌胁迫诱导最终能抵抗病原菌入侵[7]。
文心兰(Oncidiumhybridum)为兰科(Orchidaceae)文心兰属植物,被插花界誉为切花“五美人”之一,具有较高的经济价值。虽然文心兰与其他热带兰花相比,更具有广范的适应性,但在高温高湿季节恰好是文心兰的切花高峰期,此时非常容易受各种病原菌侵染,从而阻碍了文心兰产业的发展[8]。而由欧文氏菌(Erwiniaspp)侵染而引起的软腐病(Soft rot)给文心兰栽培产业带来了毁灭性破坏,因此文心兰的抗病研究是文心兰栽培过程中的重要方向[9-13]。目前实验室已做了关于OnFd基因在文心兰响应抗软腐病过程中所产生作用的相关研究[14]。然而,有关抗文心兰软腐病miRNA的研究鲜有报道。在本研究中,拟利用实验室前期已建立的文心兰miRNA数据库,以文心兰‘南茜’为对象,对文心兰miRNA的25条前体(pre-miRNAs)序列和15条成熟体序列、前体二级结构及其在不同组织部位和软腐病侵染下的表达谱进行系统分析,以期为文心兰抗病的调控提供理论参考。
1 材料和方法
1.1 材 料
本试验所用文心兰植株以及文心兰转录组和miRNA数据库由福建农林大学园艺植物生物工程研究所提供。以‘南茜’文心兰植株为材料,选取长势相近的健康文心兰植株10株,取其根、茎、叶组织材料,用于实时荧光定量PCR分析;同时,选取长势相近的健康文心兰植株10株,以制备好的软腐病菌液注射侵染假鳞茎,放置于设定好的人工气候箱培养,处理0、4、8和12 h 后取假鳞茎部位。病原菌为欧文式杆菌,其获得是从实验室已分离出来保存于-80 ℃冰箱的软腐病菌菌液进行纯化,病原菌的分离参考吴晓佩[13]方法。试验均3次生物学重复。样品经液氮处理后保存于-80 ℃冰箱,用于总RNA的提取及后续试验。
1.2 方 法
1.2.1文心兰pre-miRNAs序列的获取从本实验室的文心兰转录组和miRNA数据库筛选pre-miRNAs和成熟体序列。从文心兰miRNA前体中,扣除重复序列;同时获取成熟体序列。
1.2.2文心兰pre-miRNAs及成熟体序列分析利用DNAMAN ver.6.0软件对上述获得的文心兰前体序列进行多重比对分析,并对文心兰前体及成熟体序列进行比对分析;利用3.5版在线软件Mfold (http://unafold.rna.albany. edu/?q=mfold)对其进行二级结构预测。
1.2.3文心兰不同组织部位和软腐病菌侵染过程中pre-miRNAs表达的实时荧光定量分析为了进一步了解文心兰pre-miRNAs在植株中的潜在功能,首先采用Trizol UP RNA 提取试剂盒(全式金生物技术有限公司),提取样品不同组织部位和软腐病侵染下假鳞茎的总RNA。用超微量分光光度计(Thermo Electron Corp.,USA)检测RNA 样品浓度,并用1%琼脂糖凝胶电泳检测其完整性。用Prime ScriptTMRT Reagent逆转录试剂盒(TaKaRa),将已经提取的各个组织部位总RNA反转录为cDNA。其次,利用DNAMAN ver.6.0对25条文心兰前体序列进行引物设计(表1)。引物均由尚亚生物公司合成。参考TaKaRa SYBR Premix Ex TaqTM试剂盒说明书的方法,使用罗氏LightCycler480实时荧光定量PCR仪,检测25条pre-miRNAs在文心兰不同组织部位及软腐病侵染下假鳞茎中的表达情况。选用miR168-unigene0047942为内参基因,根据2-ΔΔCt公式计算25条pre-miRNAs在文心兰不同组织部位和软腐病侵染下假鳞茎中的相对表达量,3次生物学重复。

表1 文心兰25条pre-miRNAs qPCR引物
1.2.4统计分析数据统计与图表制作用Excel软件。用在线网站Omicshare(http:// www.omicshare.com)作文心兰pre-miRNAs的表达量热图。
2 结果与分析
2.1 文心兰25条pre-miRNAs的筛选及序列分析
为了解文心兰pre-miRNAs及成熟体序列特性,本研究从文心兰转录组和miRNA数据库中筛选文心兰miRNA 31条前体,扣除重复序列,共获得分布于13个家族的25条pre-miRNAs序列,并根据其在unigene中的位置命名(表1)。在这13个pre-miRNAs家族中,miR398、miR535和miR2950家族各含2个基因座,而miR162、miR167和miR396家族均含有3个基因座,miR168家族含4个基因座。这25条pre-miRNAs序列长度在259 (miR2950-unigene0006151)~ 883 nt(miR169-unigene0022341)间,其GC含量在29.0%~ 40.0%之间。另外,从文心兰转录组数据库和miRNA文库中筛选获取分布于13个家族的15条成熟体序列(表2)。
利用DNAMAN ver.6.0软件对上述25条文心兰miRNA前体及15条成熟体进行多序列比对,结果如图1所示。除miR162家族和miR167家族有2~3个碱基差异,miR396家族中的miR396-u0019032只有8个碱基发生重叠,其余各家族序列存在一个明显的完全重叠区,推测文心兰miRNA成熟体可能来源于此区域。可见文心兰miRNA家族成员进化过程可能存在高度保守区,且不同家族在由前体形成成熟体的过程中可能存在特异性。
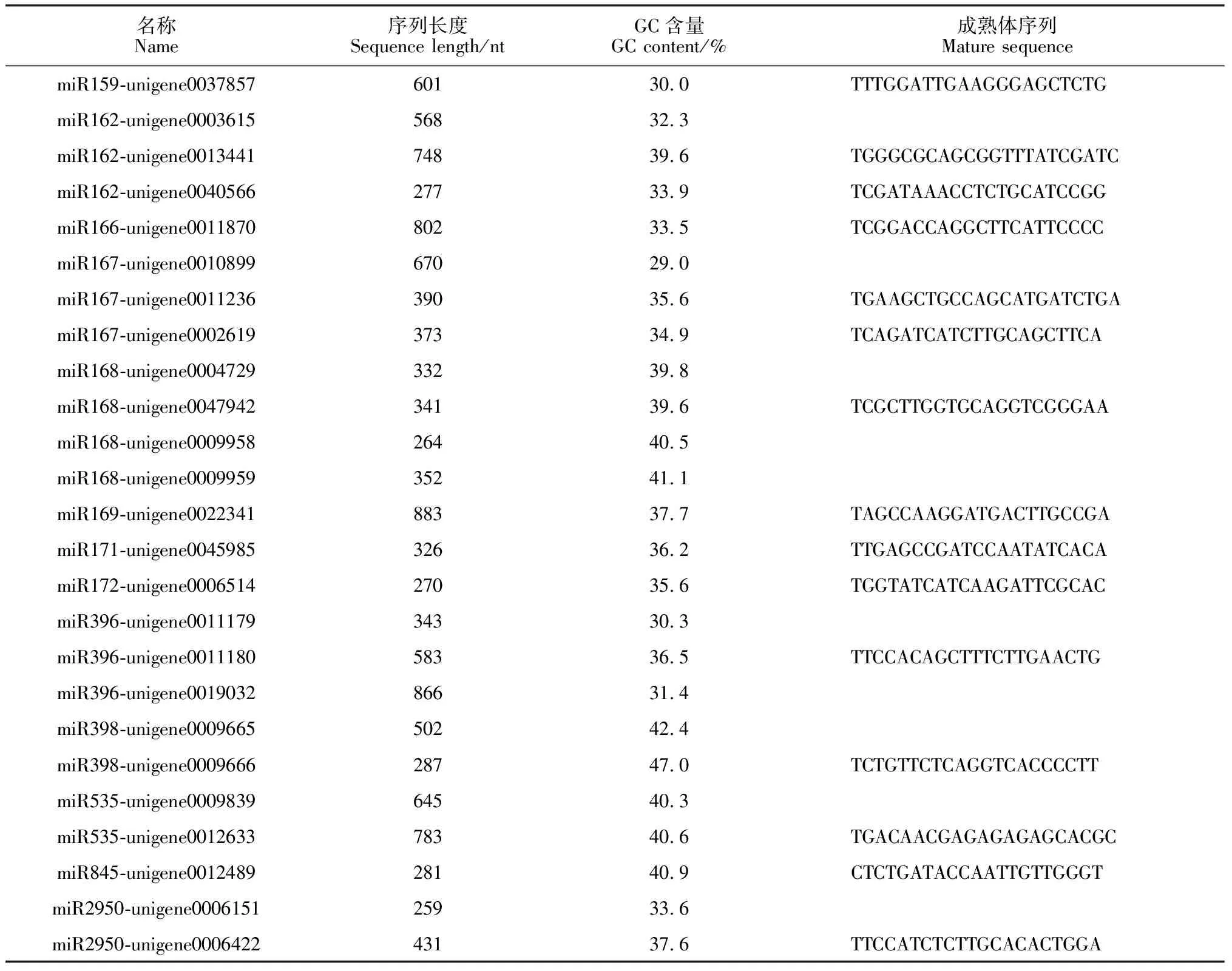
表2 文心兰25条pre-miRNAs 及15条成熟体

A. miR159-unigene0037857与miR159a;B. miR162家族与miR162a、162b;C. miR166-unigene0011870与miR166a;D. miR167家族与miR167a、167b;E. miR168家族与miR168a;F. miR169-unigene0022341与miR169a;G. miR171-unigene0045985与miR171a;H. miR172-unigene0006514与miR172a;I. miR396家族与miR396a;J. miR398家族与miR398a;K. miR535家族与miR535a;L. miR845-unigene0012489与miR845a;M. miR2950家族与miR2950-5p图1 文心兰pre-miRNAs家族序列比对分析A. miR159-unigene0037857 and miR159a; B. miR162 family and miR162a, 162b; C. miR166-unigene0011870 and miR166a; D. miR167 family and miR167a, 167b; E.miR168 family and miR168a; F. miR169-unigene0022341 and miR169a; G. miR171-unigene0045985 and miR171a; H. miR172-unigene0006514 and miR172a; I. miR396 family and miR396a; J. miR398 family and miR398a; K. miR535 family and miR535a; L. miR845-unigene0012489 and miR845a; M. miR2950 family and miR2950-5pFig.1 Analysis of pre-miRNAs sequences in O. hybridum
2.2 文心兰25条pre-miRNAs 包含的miRNA成熟体序列分析
此外,本研究对文心兰25条pre-miRNAs中提取出的15条miRNA成熟体,通过与已经登录到miRBase的植物miRNA成熟体进行比对分析。结果(图2)显示,不同物种miRNA成熟体序列长度绝大多数为21 nt,也有部分为22 nt,说明不同物种的各个家族miRNA成熟体主要是以21 nt的长度存在并发挥作用。15条文心兰miRNA成熟体与拟南芥、小麦、大豆、玉米等植物的miRNA成熟体出现多条成熟体序列完全一致的现象;但也有部分成熟体存在较大的差异(图2),如miR162、miR167、miR171、miR172、miR845这5个家族:miR162、miR167家族均存在2条成熟体,2条成熟体之间的差异较为明显,但2条成熟体均能与其他物种的同一家族成熟体重叠;miR171、miR172,miR845这3个家族的成熟体与其他物种的miRNA成熟体存在着明显的碱基差异(图2)。说明了植物miRNA家族的保守性与特异性并存。

ath. 拟南芥;sbi. 高粱;osa. 水稻;bdi. 二穗短柄草;zma. 玉米;mtr. 苜蓿;ptc. 毛果杨;gma. 大豆;ctr. 柑橘;ccl. 克莱门柚;gra. 雷蒙德氏棉;csi. 甜橙;ghr. 棉花;aly. 琴叶拟南芥;ppt. 小立碗藓;vvi. 葡萄;atr. 无油樟;mes. 木薯; A. miR159a;B. miR162a、162b;C. miR166a;D. miR167a、167b;E. miR168a;F. miR169a;G. miR171a;H. miR172a;I. miR396a;J. miR398a;K. miR535a;L. miR845a;M. miR2950-5p图2 文心兰miRNA家族成熟体序列比对分析ath. Arabidopsis thaliana; sbi. Sorghum bicolor; osa. Oryza sativa; bdi. Brachypodium distachyon; zma. Zea mays; mtr. Medicago truncatula; ptc. Populus trichocarpa; gma. Glycine max; ctr. Citrus trifoliata; ccl. Citrus clementina; gra. Gossypium raimondii; csi. Citrus sinensis; ghr. Gossypium hirsutum; aly. Arabidopsis lyrata; ppt. Physcomitrella patens; vvi. Vitis vinifera; atr. Amborella trichopoda; mes. Manihot esculentaFig.2 Analysis of mature sequences of miRNA family in O. hybridum
2.3 文心兰25条pre-miRNAs二级茎环结构分析
本研究采用mfold在线软件对文心兰25条pre-miRNAs序列进行二级茎环结构预测与分析。所研究的文心兰pre-miRNAs序列长度偏长,因此,在本研究中,截取成熟体上下游各100 bp左右片段长度进行二级茎环结构预测。结果(图3)显示,文心兰pre-miRNAs折叠之后均能形成典型、复杂的发夹结构,但各个成员间的结构存在差异。miR167-unigene0010899、miR167-unigene0011236中有2个小环,miR168-unigene0009958、miR172-unigene0006514中有3个小环,miR162-unigene0013441、miR162-unigene0040566、miR168-unigene0009959、miR396-unigene0011179、miR396-unigene 0019032、miR845-unigene0012489中有4个小环,miR162-unigene0003615、miR168-unigene 0004729、miR396-unigene0011180、miR398-unigene 0009665、miR398-unigene0009666、miR2950-unigene0006422中有6个小环,miR166-unigene 0011870、miR168-unigene0047942、miR169-unigene0022341、miR171-unigene0045985、miR535-unigene0009839、miR2950-unigene 0006151中有7个小环,miR159-unigene0037857中有8个小环,miR167-unigene0002619中有9个小环,miR535-unigene0012633中有10个小环。其序列的最小折叠自由能在-32.04~-120.98 kal/mol之间,在一定程度上,文心兰pre-miRNAs序列的碱基数越多,折叠后最小自由能的负值越大,序列G/A比值、茎环数目等因素也可能影响着最小折叠自由能。
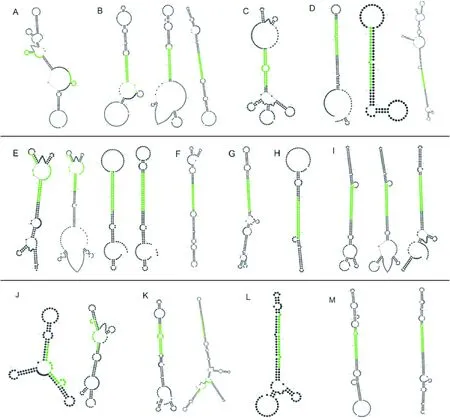
2.4 25条pre-miRNAs在文心兰不同组织部位中的定量表达分析
为进一步探究文心兰miRNA家族不同成员在文心兰中的时空表达特性,本研究采用qRT-PCR技术检测pre-miRNAs在‘南茜’文心兰不同组织部位中的表达情况。结果(图4)表明,除了miR162-unigene0040566、miR167-unigene0010899、miR168-unigene0004729、miR168- unigene0047942、miR168-unigene0009959、miR398-unigene0009665、miR535-unigene 0009839、miR535-unigene0012633这8个前体在文心兰不同组织部位中不表达外,其余pre-miRNAs在文心兰生长发育过程中的转录水平存在着明显的差异。主要可以分为以下3类:miR159-unigene0037857、miR167-unigene0011236、miR167-unigene0002619、miR169-unigene0022341、miR172-unigene006514、miR396-unigene19032、miR398-unigene0009996、miR2950-unigene0006151、miR2950-unigene0006422等9个前体均在叶中大量表达,而在根中低表达;miR162-unigene0003615、miR162-unigene0013441、miR166-unigene 0011870、miR168-unigene0009958在根和叶中大量表达,在茎中低表达;而miR171-unigene 0045985、miR396-unigene-0011179、miR396-unigene0011180在根和茎中大量表达,在叶中低表达。可以看出miR162-unigene0003615、miR162-unigene0013441、miR166-unigene0011870、miR168-unigene0009958和miR171-unigene0045985、miR396-unigene0011179、miR396-unigene0011180在叶和茎中表达趋势相反,提示它们在功能上可能存在互补性。miRNA各个家族在文心兰生长发育过程中的表达模式各不相同,它们可能参与文心兰不同器官生长发育过程的维持。
A. miR159-unigene0037857, dG=-33.6 kal/mol; B. miR162-unigene0003615, dG=-34.79 kal/mol; miR162-unigene 0013441, dG=-35.20 kal/mol; miR162-unigene0040566, dG=-67.00 kal/mol; C. miR166-unigene0011870, dG= -58.44 kal/mol; D. miR167-unigene0010899, dG=-62.10 kal/mol; miR167-unigene0011236, dG=-37.20 kal/mo; miR167-unigene 0002619, dG=-78.33 kal/mol; E. miR168-unigene0004729, dG=-49.16 kal/mol; miR168-unigene0047942, dG=-47.32 kal/ mol; miR168-unigene0009958, dG=-49.58 kal/mol; miR168-unigene0009959, dG=-50.50 kal/mol; F. miR169-unigene0022341, dG=-72.39 kal/mol; G. miR171-unigene0045985, dG=-44.76 kal/mol; H. miR172-unigene0006514, dG=-52.70 kal/mol; I. miRmiR396-unigene0011179, dG=-64.08 kal/mol; miR396-unigene0011180, dG=-70.56 kal/mol; miRmiR396-unigene 0019032, dG=-59.04 kal/mol; J. miR398-unigene0009665, dG=-32.04 kal/mol; miR398-unigene0009666, dG=-36.56 kal/mol; K. miR535-unigene0009839, dG=-56.10 kal/mol; miR535-unigene0012633, dG=-120.98 kal/mol; L. miR845-unigene 0012489, dG=-37.48 kal/mol; M. miR2950-unigene0006151, dG=-47.40 kal/mol; miR2950-unigene0006422, dG=-56.64 kal/mol图3 文心兰pre-miRNAs二级茎环结构分析Fig.3 Hairpin structure analysis of pre-miRNAs sequences in O. hybridum
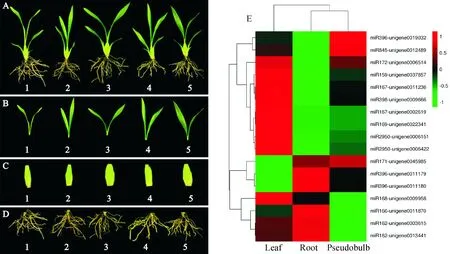
A. 植株;B. 叶;C. 假鳞茎;D. 根;E. pre-miRNAs家族表达聚类分析图4 pre-miRNAs家族在文心兰不同组织部位中的表达分析A. Plant; B. Leaves; C. Pseudobulb; D. Roots; E. Cluster analysis of pre-miRNAs family expressionFig.4 Expression analysis of pre-miRNAs family in different tissues of O. hybridum

A. 植株;B. 假鳞茎;C. 文心兰pre-miRNAs家族表达聚类分析图5 pre-miRNAs家族在文心兰软腐病菌侵染过程中的表达分析 A. Plant; B. Pseudobulb; C. Cluster analysis of pre-miRNA family expressionFig.5 Expression analysis of pre-miRNAs family in the soft rot pathogen infection of O. hybridum
2.5 25条pre-miRNAs在软腐病菌侵染文心兰假鳞茎中的定量表达分析
进一步选取长势相近的健康文心兰植株,以20 μL软腐病菌液注射侵染,放置于温度为35 ℃、80%湿度、20%光照的人工气候箱培养12 h,注射软腐病病菌0、4、8和12 h 后取假鳞茎部位,对文心兰pre-miRNAs进行实时荧光定量分析。结果(图5)表明,去除miR162-unigene0013441、miR167-unigene0010899、miR168-unigene0004729、miR168-unigene0009959、miR172-unigene0006514、miR398-unigene0009665、miR535-unigene0009839、miR535-unigene0012633、miR2950-unigene0006151、miR2950-unigene0006422这10个不表达的前体外,其余pre-miRNAs在文心兰软腐病菌液侵染中的转录水平存在着明显差异,并可以分3类:miR159-unigene0037857、miR167-unigene0011236、miR396-unigene0011179、miR396-unigene 0011180、miR396-unigene0019032、miR845-unigene0012489的表达量在软腐病菌侵染后呈现下降趋势并在处理12 h后表达量达到最低;miR162-unigene0040566、miR168-unigene0009958、miR171-unigene0045985、miR166-unigene0011870在软腐病菌液侵染4 h后其表达量升高,之后表达量下调,说明这4个miRNA可能在病菌侵染早期响应软腐病侵染过程;miR162-unigene0003615、miR167-unigene0002619、miR169-unigene 0022341、miR845-unigene0012489、miR168-unigene 0047942在菌液侵染过程中其表达量呈现先下降后上升的趋势,并在侵染8 h后表达量达到最低,提示上述4条miRNA可能通过下调表达而参与了软腐病侵染过程。上述结果表明,不同家族的miRNA均在一定程度上参与了文心兰软腐病菌侵染过程,但其响应方式与响应时期则各不同,不同miRNA家族可能通过上调或下调表达参与菌液侵染过程,且不同家族的miRNA 均分散在各个时期出现明显的上调或者下调表达,提示它们可能在软腐病菌对文心兰的侵染过程中具有功能上的互补性。
3 讨 论
3.1 文心兰pre-miRNAs和成熟体的进化特性
miRNA通过碱基互补原则切割靶基因mRNA[15-18]、介导DNA甲基化[19-21]、抑制翻译[22-24]等方式在植物生长发育中起着调控功能[26]。植物miRNA的进化特性分析能在一定程度上阐明miRNA的功能。近年来,随着测序技术的不断发展,对于miRNA进化特性的研究已取得一定进展。研究发现,miRNA家族一般在植物中呈现出保守性,如对miR159[27]、miR166[28]、miR171[29]家族的进化特性分析结果显示植物这3个家族均具有较高的保守性。目前已发现的miRNA家族中,miRNA前体中非成熟序列存在较大差异,推测可能与物种在长期自然选择过程中出现基因缺少或插入突变相关。例如,本研究中25条pre-miRNAs中提取出15条成熟序列,除了存在一个明显的完全重叠区之外,其非成熟序列存在着实质性差异,这有可能是文心兰在长期自然选择过程中突变积累并固定的结果。然而大多数miRNA家族的成熟体序列均具有较高的保守性,通过miRBase下载所有物种的miRNA成熟体序列并进行比对,发现miR159a、miR166a、miR168a、miR171a、miR172a、miR398a等家族的miRNA具有较高的物种间保守性。本研究从miRBase数据库提取拟南芥、水稻、小麦、玉米、苜蓿、柑橘、甜橙等物种不同家族的miRNA成熟体序列与文心兰不同家族成熟体进行比对分析,发现文心兰miR159a、miR166a、miR168a、miR169a、miR396a、miR398a、miR535a、miR2950-5p等miRNA成熟体与其他物种间具有极高的序列相似性,推测这几个miRNA家族可能具有较高的保守性;而miR162a、miR162b、miR167a、miR167b、miR171a、miR172a等miRNA成熟体物种间的保守性较低,推测部分miRNA家族可能存在物种间的特异性。
3.2 pre-miRNAs可能广泛参与了文心兰不同组织器官的生长发育过程
关于miRNA参与植物生长发育过程的研究已经很多,不同miRNA家族在植物的生命周期中均发挥了不同的作用。如草莓miR159a和miR159b通过靶向MYB基因,参与了草莓花器官的生长[30];拟南芥miR171c可通过抑制其靶基因SCL6的表达,最终抑制其腋芽的发育[31];玉米miR172通过调控其靶基因gl15的表达,从而调控其叶片的发育[32];龙眼中的pre-miR398b可能在龙眼体胚发生过程中参与了球形胚和子叶胚的发育过程[33]。在本研究中,以文心兰不同组织部位根、茎、叶为材料,通过qRT-PCR方法对文心兰25条pre-miRNAs 进行定量分析发现miR159-unigene0037857、miR167-unigene0011236、miR167-unigene0002619、miR169-unigene0022341、miR172-unigene006514、miR396-unigene19032、miR398-unigene0009996、miR2950-unigene0006151、miR2950-unigene0006422等pre-miRNAs在叶片中大量累积可能有利于文心兰叶的形态建成;miR162-unigene0003615、miR162-unigene0013441、miR166-unigene 0011870、miR168-unigene0009958等pre-miRNAs在根和叶中大量表达促进文心兰叶和根的发育;而miR171-unigene 0045985、miR396-unigene0011179、miR396-unigene0011180等pre-miRNAs同时在根和茎中大量表达可能利于其根和茎的发育。此外,miR162-unigene0003615、miR162-unigene0013441、miR166-unigene0011870、miR168-unigene0009958和miR171-unigene0045985、miR396-unigene00111 79、miR396- unigene0011180在叶和茎中表达趋势相反,提示它们在功能上可能存在互补性。而miR162-unigene0040566、miR167-unigene0010899、miR168-unigene 0004729、miR168-unigene0047942、miR168-unigene0009959、miR398-unigene 0009665、miR535-unigene0009839、miR535-unigene 0012633等 8个pre-miRNAs在文心兰不同组织部位中不表达,暗示其可能并未参与植物组织分化与形态建设等;miRNA各个家族在文心兰生长发育过程中的表达模式各不相同,提示它们可能广泛参与了文心兰不同器官的生长发育过程。
3.3 文心兰软腐病侵染过程的不同阶段可能由不同miRNA参与调控
miRNA在调控植物基因表达、生长发育和抵抗胁迫及病害等方面扮演着重要的角色,已有的研究表明miR171[34]、miR159[6]、miR393[5]等miRNA均在参与植物生物胁迫中发挥了重要作用。研究发现根癌农杆菌的致瘤菌株可在植物的注射位置诱导miR393的表达,而非致瘤菌株则不会诱导[35]。miR393和miR167能够抑制生长素信号通路,在根癌农杆菌诱导后的根瘤中,其表达水平明显降低[5]。小麦白粉病叶片中克隆得到153个miRNA,其中24个与白粉病相关[36]。在拟南芥中过量表达芜菁花叶病毒TuMV毒性蛋白基因,发现miR171表达水平降低,其靶基因表达水平上调,植物出现病毒病症状;另外,通过改造拟南芥miR159的前体获得能够靶向Hc-Pro的人造miRNA,过表达后发现,其抗病性增强[6];病原菌胁迫诱导miR398表达下调进而导致其靶标CSD过氧化物歧化酶基因上调表达,CSD基因在植物体内大量富集从而激活其自身防御系统,产生系列的防卫反应并启动抗氧化胁迫机制,最终抵抗病原菌入侵[7]。本研究以‘南茜’文心兰不同组织部位为材料,通过qRT-PCR技术对pre-miRNAs在感病文心兰假鳞茎的表达情况进行分析,结果显示不同pre-miRNAs在文心兰感病过程中的表达趋势均有差异。miR162-unigene0013441、miR167-unigene0010899、miR168-unigene0004729、miR168-unigene0009959、miR172-unigene 0006514、miR398-unigene0009665、miR535-unigene0009839、miR535-unigene 0012633、miR2950-unigene0006151、miR2950-unigene0006422等10个pre-miRNAs在文心兰软腐病侵染过程中不表达,揭示其或许未能参与文心兰植株软腐病的应激过程。miR159-unigene0037857、miR167-unigene0011236、miR396-unigene 0011179、miR396-unigene0011180、miR396-unigene0019032、miR845-unigene00 12489的表达量在软腐病菌侵染后呈现下降趋势并在处理12 h后表达量达到最低,提示这6个pre-miRNAs可能通过下调miRNA的表达,进而导致其相应靶标基因的上调表达,最终参与了文心兰的抗病过程。miR162-unigene0040566、miR168-unigene0009958、miR171- unigene 0045985、miR166-unigene0011870在软腐病菌菌液侵染4 h后其表达量升高,之后表达量下调,说明这4个pre-miRNAs可能在软腐病菌侵染早期响应软腐病侵染过程;miR162- unigene0003615、miR167-unigene0002619、miR169-unigene0022341、miR845-unigene0012 489、miR168-unigene 0047942在软腐病菌菌液侵染过程中其表达量呈现先下降后上升的趋势,在0 h时表达量最高,并在侵染8 h后表达量达到最低,提示上述4个pre-miRNAs可能通过下调表达而参与了软腐病侵染过程,并在处理8 h后其响应效果最明显;而该研究将为探讨文心兰miRNA家族在文心兰抗病过程中的作用提供一定参考依据。
