小麦阿拉伯木聚糖阿魏酸酰基转移酶基因的 克隆与功能标记开发
2018-10-16战帅帅白璐谢磊夏先春任毅吕文娟曲延英耿洪伟
战帅帅,白璐,谢磊,夏先春,任毅,吕文娟,曲延英,耿洪伟
小麦阿拉伯木聚糖阿魏酸酰基转移酶基因的 克隆与功能标记开发
战帅帅1,白璐2,谢磊1,夏先春3,任毅1,吕文娟1,曲延英1,耿洪伟1
(1新疆农业大学农学院/新疆农业大学生物技术重点实验室,乌鲁木齐 830052;2新疆农业大学科学与技术学院,乌鲁木齐 830091;3中国农业科学院作物科学研究所/国家小麦改良中心,北京 100081)
【目的】克隆小麦阿拉伯木聚糖阿魏酸酰基转移酶(arabinoxylan feruloyl transferase,AFT)基因,开发与FAX含量紧密连锁的功能标记,提高对FAX含量预测的准确性,为小麦加工品质的改良提供依据。【方法】应用FR846233为探针,通过同源克隆的方法获得小麦FAX含量基因gDNA全序列,使用DNAMAN软件比较高、低FAX含量品种间的序列差异性;基于序列差异使用Primer5.0软件设计特异性引物,开发与FAX含量紧密连锁的功能标记并利用一套中国春缺体-四体系和3AL、3AS双端体系进行染色体物理定位;同时结合PCR验证的方法,利用253份来自于中国主要冬麦区的小麦品种(系)对功能标记的实用性进行验证,采用IBM SPSS statistics 19.0软件进行FAX含量与基因型间的相关性分析等数据统计分析。【结果】2对特异性引物B1、B2最终扩增出长度分别为800 bp及710 bp的片段,B1和B2扩增的PCR片段有80 bp重叠,将片段拼接得到位于3A染色体上AFT基因。序列由1 429个碱基对组成,得到等位变异和,2个等位变异都拥有一个1 266 bp的开放阅读框(open reading frame,ORF),都具有2个外显子和1个内含子,内含子符合典型GT-AG结构,等位变异序列之间相似度为98.08%,具有24个单核苷酸多态性(single nucleotide polymorphisms,SNPs)位点和3个插入缺失(insertion-deletion,InDel)InDel位点,可编码421个氨基酸残基,预测分子量为45.2 kD。基于107 bp SNP处开发了2个互补显性标记AFTA2和AFTB2。AFTA2在材料中能扩增出692 bp片段,与高FAX含量相关,在具有等位变异的材料中未能扩增出片段。AFTB2则只能在等位变异类型的材料中扩增出438 bp片段,并与低FAX含量相关,在等位变异类型的材料中未能扩增出片段。同时利用一套中国春缺体-四体系和双端体材料将AFTA2和AFTB2定位在小麦3AL染色体。用功能标记AFTA2和AFTB2检测253份中国冬小麦材料,结果表明,不同基因型的FAX含量差异达到显著水平(<0.05)。从不同麦区来看表现各有不同,其中在北部冬麦区不同基因型的FAX含量在北部冬麦区材料间差异不显著;而在黄淮麦区,含有的品种FAX含量显著高于含有的品种(<0.05)。等位变异分布频率表明,是与高FAX含量相关优异等位变异,且北部冬麦区的频率(71.3%)显著高于黄淮冬麦区(60.2%)。【结论】基于序列成功开发1对显性互补标记AFTA2/AFTB2并定位于3AL染色体,标记与FAX含量相关可用于FAX含量的遗传改良。
普通小麦;阿魏酰阿拉伯木聚糖;基因克隆;功能标记开发
0 引言
【研究意义】随着人们生活水平的提高,小麦品质已是影响产业竞争力的重要因素,小麦品质改良已成为小麦育种和生产的重要目标[1-4]。阿拉伯木聚糖(arabinoxylan,AX)是小麦中一种非淀粉多聚糖,通过阿魏酰阿拉伯木聚糖(feruloyl arabinoxylan,FAX)结构中的阿魏酰基氧化交联形成凝胶网络,进而对小麦面团流变学特性及其加工品质产生影响[5]。AX不仅能增加面团的吸水能力和黏度,提高面筋强度[6],而且还成为倍受关注的膳食纤维重要组分[7],具有降低血清胆固醇、调节血糖水平、抗氧化等重要生理功能[8],因而与小麦的营养品质与健康品质密切相关。FAX由阿拉伯木聚糖阿魏酸酰基转移酶(arabinoxylan feruloyl transferase,AFT)催化合成,其含量高低是决定小麦AX结构和性质的关键因素[9]。功能标记是依据目标基因多态性序列引起表型性状差异开发出来的一种新类型标记,可以多种遗传背景下确定目标等位基因的存在[10-11]。因此,探究AFT基因的特性并开发相应的功能标记,有助于提高对表型预测的准确性,提高分子标记辅助育种的效率,对于改良小麦品质和分子辅助育种具有重要意义。【前人研究进展】FAX含量是由多基因控制的复杂数量性状,受环境影响大,但其主要受遗传因素影响[12],直接对多基因控制的复杂数量性状进行选择难度较大,利用分子标记聚合相关基因是进行遗传改良的重要途径。Martinant等[13]以面粉提取物黏度等参数指标发现遗传相关性,并在Synthetic与Opata杂交小麦的1BL染色体上发现了相对黏度和Ara/Xyl比值有关的数量特性基因位点。Rao等[14-15]研究表明在不同植物或同种植物不同品种间FAX含量的差异很大。ORDAZ-ORTIZ等[16]也发现小麦品种间同时存在AX数量和结构的差异,而且水溶性阿拉伯木聚糖(water-extractable arabinoxylan,WEAX)与水不溶性阿拉伯木聚糖(water-unextractable ara-binoxylan,WUAX)的结构变化趋势(Ara/Xyl)基本一致。木糖(Xyl)和阿拉伯糖(Ara)聚合成戊聚糖(Pentosans),通常称之为阿拉伯木聚糖[17]。张岐军等[17]以17份软质小麦品种进行了两年多点试验,并提出在软质小麦育种中应选取戊聚糖含量低的品种。在总戊聚糖含量未显著差异的前提下,软麦的水溶性戊聚糖含量显著低于硬麦[18]。植物中特有的BAHD酰基转移酶家族,是次生代谢产物酰基化的一类修饰蛋白,对多种酰基化生物合成有重要关系[19]。AFT基因是BAHD基因家族成员之一,且阿拉伯木聚糖阿魏酸酰基转移酶是影响FAX含量的关键酶[20]。Mitchell等[20]发现AFT基因由酰基转移酶基因家族成员所编码。如今,水稻中AFT基因已被定位于第1染色体组[21]。根据禾本科植物基因共线性原理,结合水稻中AFT基因的定位结果,Tanaka等[22]和MATSUMOTO等[23]推测小麦AFT基因位于第3同源染色体组。Li等[24]利用PH82-2/内乡的240个重组自交系群体(recombinant inbred lines,RIL)测定小麦籽粒中总AX含量(TOT-AX),共使用195个SSR和STS标记构建遗传图谱并检测数量性状基因座,在1A、1B、2B、3B、5A、5B、6B、7A和7B染色体上鉴定了9个加性QTL,解释了3.8%—15.2%的表型变异。吕文娟等[25]利用周麦16/藁城8901的176个RIL群体的表型数据同时使用90 K的iSelect基因芯片针对FAX含量作了全基因组连锁分析,在RIL群体中发现了小麦FAX含量的5个主效QTL位点,分布在2AS、2BS、2DL、3AS和6AS染色体上,单个QTL可解释的6.3%—18.2%的表型变异,其中在3A染色体上发现的位点可解释最大的表型变异。【本研究切入点】目前,国内外对FAX性能关注度较高[26-27],而对FAX含量的遗传特性报道较少。吕文娟等[25]通过全基因组关联分析,利用距最近的SNP标记对200份冬小麦品种(系)进行检测,验证了小麦第3同源染色体组上存在调控FAX含量的主效基因。但这些SNP标记与基因之间尚有一定距离(1.3 cM),为了更准确地鉴定小麦品种FAX含量的基因型,有必要对主效位点基因进行克隆,并根据该基因位点不同等位基因的序列开发共分离的功能标记,进一步提高分子标记选择的准确性和育种效率。【拟解决的关键问题】本研究通过同源克隆获得小麦FAX含量基因gDNA全序列,且根据高、低FAX含量品种间的序列差异设计特异性引物来挖掘与FAX含量紧密连锁的功能标记,同时利用253份来自于中国主要冬麦区的小麦品种(系)对功能标记的实用性进行验证,明确等位基因在冬麦区的分布。
1 材料与方法
1.1 供试材料
中国春缺体-四体系(Chinese Spring Nullisomic- tetrasomic)和双端体系3AL、3AS材料用于功能标记的染色体物理定位,由中国农业科学院作物科学研究所小麦品质课题组赠予。藁城8901/周麦16(与吕文娟等[25]为同一RIL群体176个株系的亲本)用于基因克隆,于2013—2014年度和2014—2015年度种植于河南安阳。北部冬麦区和黄淮麦区的253份国内外冬麦品种(系)用于功能标记开发验证。上述材料基本反映了中国主要冬麦区小麦育种和生产现状。87份北部小麦品种(系)分别于2012—2013年度和2013—2014年度种植于北京和石家庄,166份黄淮麦区小麦品种(系)分别于2012—2013年度和2013—2014年度种植于河南安阳和安徽濉溪。以上全部材料随机区组设计,3次重复3行区,行长2 m,行距25 cM,田间管理按照当地常规栽培管理方式,小麦田间长势良好,正常收获,无倒伏和穗发芽现象。
1.2 FAX含量测定
FAX含量使用722型分光光度计测定。试验以空白溶液为对照,用草酸溶液作为提取液,测定345和375 nm双波长下的吸光度值(A),最终依据朗伯比尔定律计算FAX的含量。具体检测方法参考吕文娟等[25]方法并略作修改。
1.3 数据的统计分析
253份冬小麦品种(系)表型数据[25]及FAX含量与基因型间的相关性分析等数据统计分析采用IBM SPSS statistics 19.0软件进行[28]。
1.4 TaBahd-A1的克隆策略
利用Primer 5.0软件,应用普通小麦AFT基因的cDNA序列FR846233为探针对GenBank数据库进行BLAST检索。将得到的相似性高的cDNA序列比对,在序列的SNP和InDel基础上,设计覆盖编码区的染色体特异性引物。同时,利用中国春缺体-四体系对所设计引物进行特异性定位,确保所克隆基因来源于3A染色体。取三粒小麦籽粒分别磨成粉末,采用SDS法提取小麦基因组DNA,详细步骤参照李冰等[29]酚氯仿法,但取用量略作调整。以小麦籽粒提取的DNA为模版,通过B1、B2引物PCR扩增AFT基因的gDNA全序列。
PCR反应条件为94℃ 5 min;94℃ 50 s,63℃/ 62℃ 50 s;72℃ 1 min;35/30个循环;72℃ 10 min,4℃保温。扩增产物经1.5%的琼脂糖凝胶、1×TAE电泳缓冲液体系电泳后,进行溴化乙锭染色。采用琼脂糖凝胶回收试剂盒进行目标条带的回收纯化,再选用平末端连接试剂盒进行连接转化,连接载体为Trins-T1的大肠杆菌感受态细胞,以上试剂均由北京全式金生物技术有限公司(http://www.transgen.com. cn)提供,最后将筛选的阳性单克隆菌液送测序,每个PCR反应最少3次。以上引物合成及测序由上海生工生物技术有限责任公司(http://www.sangon.com/)完成。
1.5 TaBahd-A1的功能标记开发
通过DNAMAN 5.0软件比较不同冬麦区高、低FAX含量品种间的序列差异性,设计多对引物用来筛选不同品种间的多态性,以期开发功能标记。依据SNP及InDel设计的引物,应使扩增产物大小适宜便于观察,从而能够较好地在琼脂糖胶上进行电泳结果分析。PCR扩增条件参照1.4,退火温度及循环数略作调整。利用一套中国春缺体-四体系和双端体系材料对标记进行染色体定位。
2 结果
2.1 TaBahd-A1的克隆
以普通小麦籽粒中的cDNA序列(GenBank登录号FR846233)为探针,在GenBank/ EMBL/DDBJ中BLAST小麦表达序列标签(Expressed sequence taq,EST)数据库,检索出3个序列:普通小麦mRNA序列(GenBank登录号AK336201)、大麦mRNA序列(GenBank登录号AK362895)和水稻mRNA(GenBank登录号AK287601)。cDNA序列FR846233是小麦全部编码序列,与上述3个序列分别具有99%、95%和83%的序列相似性。本研究基于上述4个序列的SNP和InDel,设计了2个染色体特异性引物B1和B2(表1),并通过中国春缺体-四体系将B1和B2扩增产物定位于3A染色体上(图1)。B1和B2能分别扩增出长度为800和710 bp的DNA片段,两者有80 bp的重叠区,重叠区序列相似性达100%,最后将扩增序列进行拼接得到位置在3A染色体上的的gDNA全序列。
2.2 TaBahd-A1的结构及序列分析
的gDNA序列由1 429个碱基对组成,具有一个1 266 bp完整的开放阅读框(open reading frame,ORF),同FR846233具有100%的序列相似性,包含2个外显子和1个内含子,内含子的剪接位点结构符合典型的GT-AG型(图2)。能编码421个氨基酸残基,预测分子量为45.2 kD。利用DNAMAN5.0软件对RIL群体176个株系的2个亲本藁城8901和周麦16的序列进行比对分析,获得2个等位变异序列,分别命名为和。其中与等位变异序列之间相似度为98.08%,共有24处SNP位点,而且内含子区有3处插入缺失(图2)。

表1 克隆小麦TaBahd-A1及根据TaBahd-A1不同等位变异开发功能标记的引物序列
a和b代表不同碱基长度,a代表聚合连锁反应产物大小,b代表等位基因全长大小
aandbrepresent different base lengths,arepresents the size of the polymerization chain reaction products, andbrepresents the full length of theallele
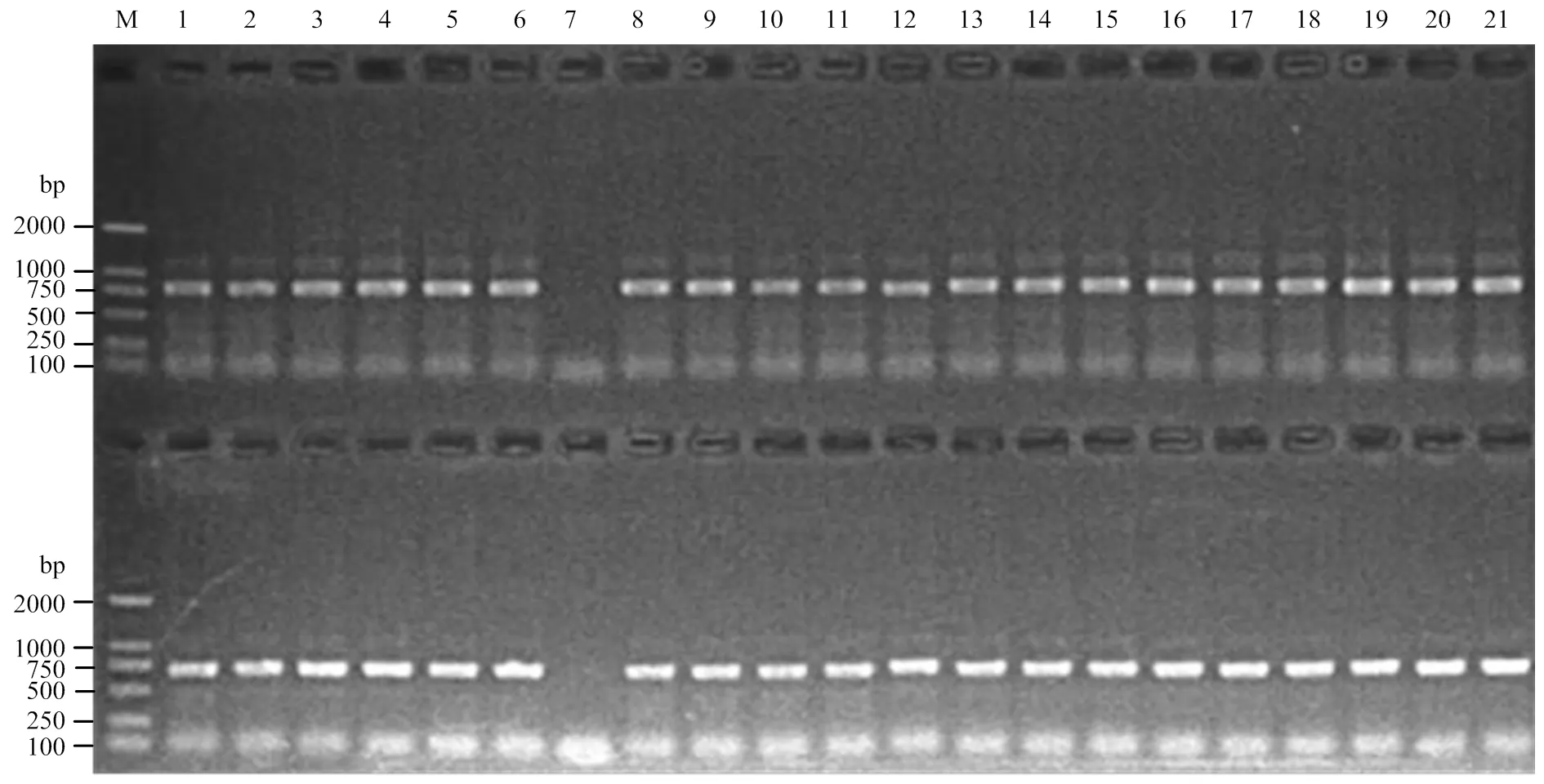
M:DL2000;1:N1A-T1B;2:N1B-T1D;3:N1D-T1A;4:N2A-T2D;5:N2B-T2A;6:N2D-T2B;7:N3A-T3B;8:N3B-T3D;9:N3D-T3A;10:N4A-T4B;11:LDN4D(4B);12:DT4BS;13:N5A-T5B;14:N5B-T5D;15:N5D-T5A;16:N6A-T6B;17:N6B-T6D;18:N6D-T6A;19:N7A-T7B;20:N7B-T7D;21:N7D-T7A
2.3 TaBahd-A1的功能标记开发
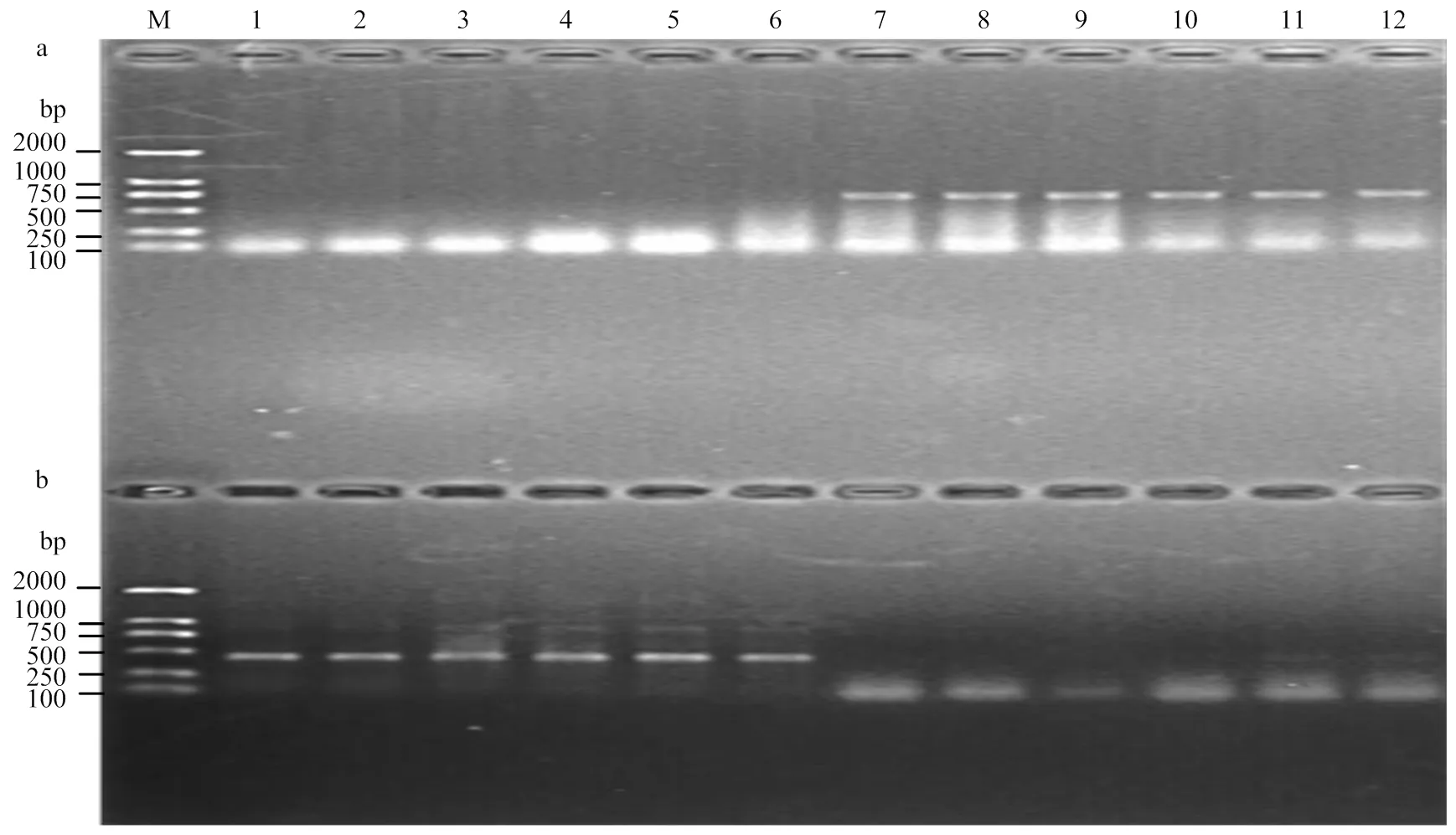
基于的等位变异与在107 bp位置的1个SNP位点,开发1对显性互补的功能标记AFTA2与AFTB2(表1)。AFTA2在具有的材料中能扩增出692 bp片段,而在具有的材料中不能扩增出片段;AFTB2在具有的材料中能扩增出438 bp片段,而在具有的材料中不能扩增出片段(图3)。利用中国春缺体-四体系和3AL、3AS双端体系对标记AFTA2/AFTB2进行染色体物理定位。表明用AFTB2扩增缺体-四体系时,仅N3A-T3B和DT-3AS没有扩增出目的DNA片段,而DT-3AL等其余泳道均扩增出438 bp的DNA片段(图4)。因此,推断所开发的标记及其对应的位于小麦3AL染色体上。
2.4 TaBahd-A1的功能标记的验证
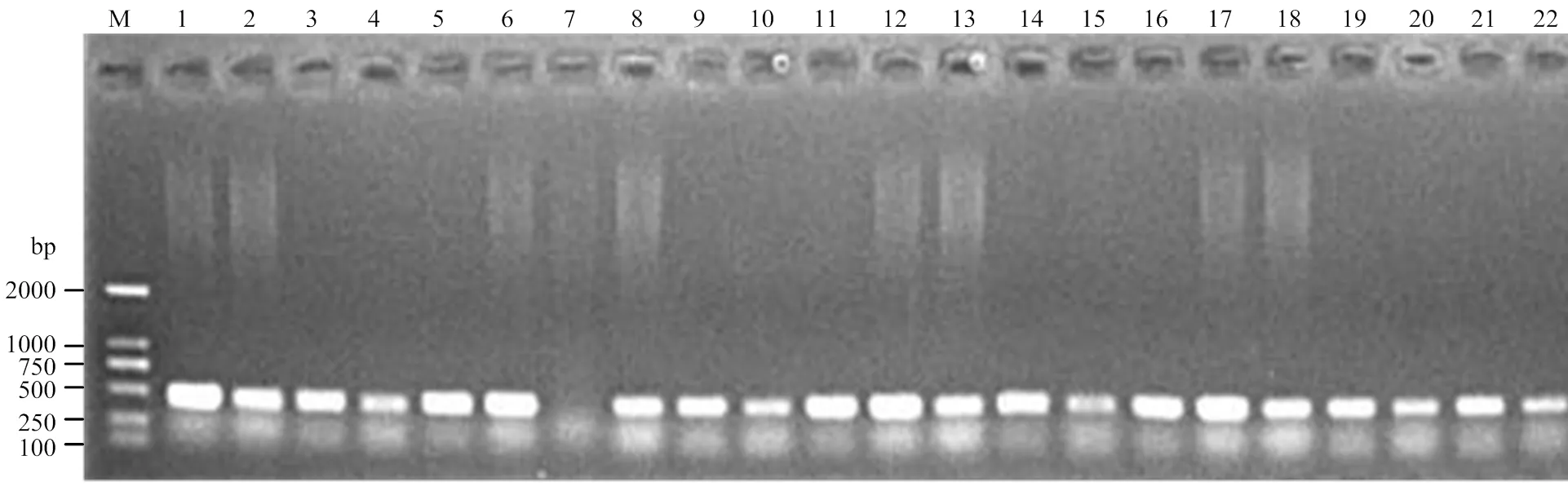
利用253份小麦品种(系)对所开发功能标记进行验证,表明黄淮麦区、北部冬麦区和全部材料中具有等位变异类型品种(系)的平均FAX含量分别为4.419、4.625和4.498×10-5mol·L-1;而具有等位变异类型品种(系)的平均FAX含量分别为4.570、4.384和4.435×10-5mol·L-1。从全部材料总体上和黄淮麦区来看,具有等位变异类型的材料FAX含量均显著高于具有等位变异类型的材料(<0.05);但在北部麦区,具有和等位变异类型品种(系)的FAX含量间差异不显著。因此,不同等位变异与FAX含量显著相关,AFTA2/AFTB2能有效应用于分子标记辅助选择育种实践,以提高对FAX含量表型预测的准确性(表2)。
2.5 TaBahd-A1等位基因的分布
通过对等位变异在北部麦区(BV)和黄淮麦区(HV)的频率分布进行统计分析。是高FAX含量优异等位变异,在253份冬小麦品种(系)中,162份材料中扩增出692 bp片段,表明含有与高FAX含量相关的占比64%;在91份材料中扩增出438 bp片段,表明含有与低FAX含量相关的占比36.0%(表3)。在北部冬麦区的87份品种(系)中,62份(71.3%)材料具有等位变异基因,25份(28.7%)材料具有等位变异基因。而黄淮冬麦区166份品种(系)中,100份(60.2%)材料具有等位变异基因(表3)。高FAX含量可使AX产生更广泛地交联,可提高小麦的面筋强度和改善小麦的加工品质。总体来看,中国小麦材料FAX含量具有较大的选择潜力,尤其是北部冬麦区品种高FAX含量频率较高,通过选择等位变异从而对提升面制品品质,提高人们的消费质量具有重要意义。

阴影表示SNP和InDel;下划线表示内含子;方框表示功能标记AFTA2和AFTB2的上游引物的反向互补序列、下游引物序列以及起始密码子
M:D2000;1:PALPICH(4.18);2:Tx03A0148(4.30);3:NSA09-3645(4.78);4 F498U(4.34);5:中麦175(4.41);6:Genio(4.10);7:Manital(4.78);8:晋麦67(4.79);9:洋小麦(4.83);10:Kitanokaori(4.55);11:Aca801(4.55);12:小堰54(4.60)。括号中数字代表FAX含量,单位为10-5 mol·L-1
M:DL2000;1:N1A-T1B;2:N1B-T1D;3:N1D-T1A;4:N2A-T2D;5:N2B-T2A;6:N2D-T2B;7:N3A-T3B;8:N3B-T3D;9:N3D-T3A;10:N4A-T4B;11:LDN4D (4B);12:DT4BS;13:N5A-T5B;14:N5B-T5D;15:N5D-T5A;16:N6A-T6B;17:N6B-T6D;18:N6D-T6A;19:N7A-T7B;20:N7B-T7D;21:N7D-T7A;22:中国春CS;23:DT-3AS;24:DT-3AL
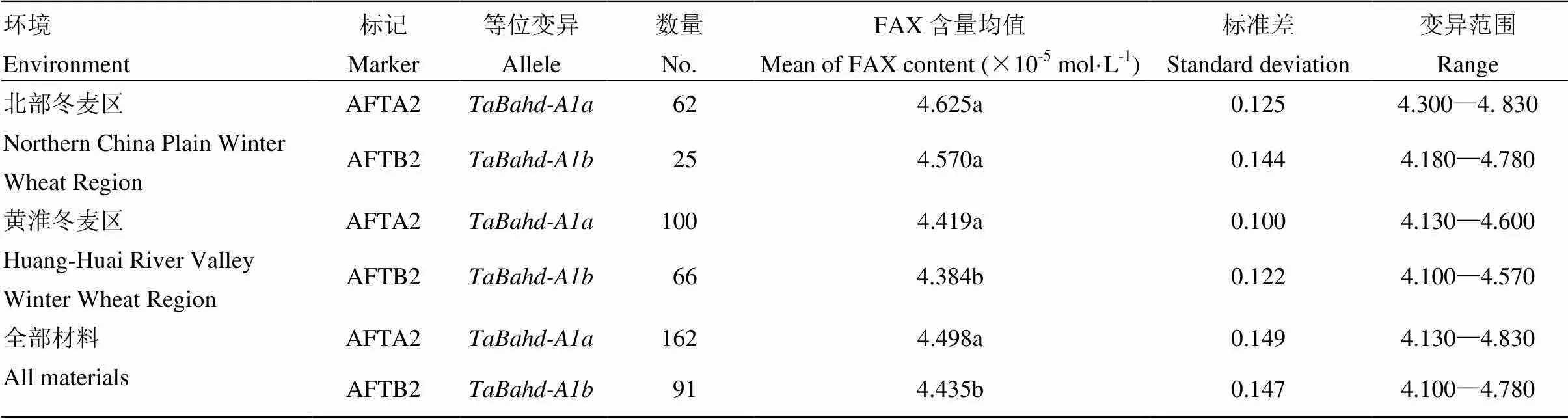
表2 利用标记AFTA2和AFTB2检测253份冬小麦材料的统计分析
不同字母表示两组间的均值差异显著(<0.05)
Mean behind the different letters represent the average difference between two groups is significant (<0.05)

表3 等位基因TaBahd-A1a和TaBahd-A1b在不同麦区的分布频率
3 讨论
3.1 小麦TaBahd-A1的克隆和染色体定位
普通小麦为异源六倍体,不仅基因组庞大复杂,而且重复序列多,不利于直接进行基因的图位克隆操作[10]。小麦大多基因在A、B、D基因组上均有分布[3],有些基因甚至在同一染色体上还有多拷贝[30],这些都增加了基因克隆和功能分析的难度,所以对基因染色体的定位和序列所在连锁群的验证是基因克隆获得成功的必要条件。本研究采用同源克隆的方法,以cDNA全序列(FR846233)为探针对GenBank数据库进行BLAST检索,将得到的相似性高的cDNA序列进行序列分析和归类。基于以上序列的SNP和InDel,设计了用于克隆的2对引物B1和B2,并用中国春缺体-四体系对2对引物染色体的特异性进行了验证,最终将其定位于3A染色体上。B1和B2分别扩增出的两DNA片段有80个bp的重叠区,且重叠区序列相似性达100%,将定位于同一3A染色体上两序列拼接起来,从而获得的全序列。同样,李冰等[29]将在二倍体、四倍体、六倍体进行比对区分最终将标记定位在5A染色体上。李亚青等[31]通过使用中国春缺体-四体系,最终利用Southern杂交将基因定位在第一同源群。
遗传图谱依据分子标记所建立,其对目标性状进行QTL分析是基因遗传定位的高效方法[3]。然而分子标记在群体偏小特别研究数量性状时会产生偏差[32],故用中国春缺体-四体系对基因进行物理定位是必要的。本研究通过用AFTA2与AFTB2扩增中国春缺体-四体系和3AS、3AL双端体系,最终成功将所开发标记定位于3AL染色体上。利用同样方法,Hessler等[32]将硬粒小麦的LOX基因定位于4BS染色体上。Sun等[33]开发了位于2A染色体上PPO基因的功能标记PPO18。而吕文娟等[25]通过全基因组连锁分析在5个染色体上均发现了小麦FAX含量主效位点,其中3AS上的位点可解释最大的表型变异(18.2%),该结果与本研究中获得的被物理定位于3AL的不一致,表明3A染色体至少存在1个与FAX含量相关基因。通过全基因组连锁分析发现位点是基于RIL群体,构建群体的亲本间遗传背景多样性的局限性影响很大,可能会对其他重要位点有所遗漏[25]。同时Carrera等[34]指出,一些与QTL连锁的分子标记并非功能标记,不能准确预测基因型。下一步我们的工作将是对位点进行精细定位,并利用合适的遗传群体对FAX含量进行QTL研究,进一步发掘与FAX含量相关的主效QTL,并开发能应用于育种实践的分子标记,有助于通过分子标记辅助选择加速FAX含量的遗传改良。
3.2 TaBahd-A1与其他作物BAHD基因的比较
水稻中位于第1染色体组的BAHD基因全长gDNA序列(GenBank登录号AK287601)已被克隆[35]。BAHD基因序列的聚类分析表明,水稻BAHD基因(AK287601)与小麦分别具有1 339和1 266 bp的开放阅读框。两者在gDNA水平的相似性为79.0%,在cDNA的相似性为83.6%,在推导出的氨基酸水平上的相似性为83.6%。国外研究发现BAHD家族大多数功能特征酶都有2个保守的功能域HXXXD和DFGWG,中HXXXD域(His159至Asp163)位于中心酶催化合成附近,该功能特征酶已被证明是乙酰转移酶催化机制的重要组分[36]。DFGWG相关机制尚未明确,可能其位置远离中心区仅起到结构作用[37]。BAHD基因位于小麦3A染色体,而在水稻中位于第1染色体。虽然它们具有不同的序列长度和序列变异,但都包含2个外显子和1个内含子。可是小麦BAHD基因(和)和水稻BAHD基因(AK287601)第1外显子和第2外显子的长度差别较大[35-37]。在2种小麦BAHD基因中,外显子大小有很高的保守性,而在不同的谷物中,外显子大小的差异比较大。以上结果表明,在同一种谷类作物特别是普通小麦中,与其他谷物相比,BAHD基因的外显子长度更保守[38]。
3.3 TaBahd-A1的功能标记的实用性评价
功能标记可以准确区分与表型相关的不同等位变异类型,从而在小麦育种中能显著提高分子标记选择的准确性和选择的高效性。小麦品质方面功能基因的挖掘和转化利用在近几年取得重要进展,并获得了多个与小麦品质相关的功能基因[1-2]。高华利等[11]表明62个相关小麦籽粒加工品质的等位基因已被克隆,共有63个功能标记被验证。由于FAX含量遗传特性研究报道较少,而与小麦AFT基因共分离的功能标记也尚未见报道。本研究中,基于开发功能标记,能有效用于FAX含量高低检测。中国小麦材料以与高FAX含量相关的优异等位变异类型为主(64%)。其中,北部冬麦区品种(系)高FAX含量相关的等位变异类型分布频率高于黄淮麦区(71.3% VS 60.2%),这可能由于北方地区的传统食品诸如馒头、面条等对小麦面筋、面团流变学特性等都有较高要求[5]。特别是自20世纪80年代后,中国开始对小麦品质研究逐步加强,为了提升面制品品质,经过长期有针对性的对高强筋、面团性好的小麦品种(系)进行选择,使得具有高FAX含量相关的等位变异类型的材料得以保留和积累。AX虽然在面粉中含量较少(4.8%—7.0%),却是小麦中膳食纤维的重要组分[5],已成为除淀粉、多糖外,近几年备受关注的小麦组分。虽然不同等位变异相关的FAX含量较为相近,但不同品种间FAX含量的差异即可通过其生理生化作用直接对加工品质产生影响,与此同时亦能间接的通过影响面筋含量等其他品质性状,从而造成材料间的加工品质差异显著。类似的现象在小麦其他小麦品质遗传改良的研究中[39]。Tremmel等[39]利用中国小麦品种豫麦34号和3个中欧品种对小麦籽粒硬度、面筋指数等参数进行了研究,结果表明,当WE-AX含量增加0.5%时,面粉的含量、面团流变学特性得到了改善,总AX含量提高1%时,千粒重量、蛋白质含量、面筋含量、面粉的水分和吸水性也会相应增加。因此,不同品种间FAX含量差异不大,但对小麦最终的加工品质而言是较为敏感成分,用相关标记进行FAX含量的遗传改良具有一定的研究意义。FAX含量由多基因控制,Li等[24]和吕文娟等[25]均在不同染色体发现不同QTL位点,同时单核苷酸位点微效累积贡献率和大量基因的影响也会对表型产生增强或抵消作用[40]。本研究中功能标记AFTA2和AFTB2与高、低FAX含量的2组材料密切相关。然而,个别基因型与FAX含量不相符,含有相同等位变异的品种间FAX含量也存在差异,并且型品种的FAX含量并非总是高于型品种,反之亦然。同样,Geng等[3]通过LOX基因等位变异一处SNP位点开发了1对显性互补标记LOX16和LOX18,表明的LOX活性并非总是高于型等位变异品种,印证小麦LOX活性受多基因控制。上述差异是由位于小麦其他染色体或同一染色体不同位点上的其他控制小麦FAX含量的QTL造成的[41-42],同时也印证了小麦FAX含量受多基因控制的结论。今后将进一步针对重要主效位点进行精细定位或者基于其他等位基因开发功能标记,通过多个组合标记检测来提高选择的效率和准确性。
4 结论
基于序列成功开发一对功能标记AFTA2/AFTB2并定位于3AL染色体,表明3A染色体可能至少存在1个与FAX含量相关基因,该标记能准确鉴定小麦品种中与FAX含量高低相关的和基因型,且北部冬麦区品种(系)高FAX含量相关的等位变异类型分布频率高于黄淮麦区,个别基因型与FAX含量不相符,含有相同等位变异的品种间FAX含量也存在差异,印证了小麦FAX含量受多基因控制。
[1] 何中虎, 晏月明, 庄巧生, 张艳, 夏先春, 张勇, 王德森, 夏兰芹, 胡英考, 蔡民华, 陈新民, 阎俊, 周阳. 中国小麦品种品质评价体系建立与分子改良技术研究. 中国农业科学, 2006, 39(6): 1091-1101.
HE Z H, YAN Y M, ZHUANG Q S, ZHANG Y, XIA X C, ZHANG Y, WANG D S, XIA L Q, HU Y K, CAI M H, CHEN X M, YAN J, ZHOU Y. Establishment of quality evaluation system and utilization of molecular methods for the improvement of Chinese wheat quality., 2006, 39(6): 1091-1101. (in Chinese)
[2] 刘志勇, 王道文, 张爱民, 梁翰文, 吕慧颖, 邓向东, 葛义强, 魏珣, 杨维才. 小麦育种行业创新现状与发展趋势. 植物遗传资源学报, 2018, 19(3): 430-434.
LIU Z Y, WANG D W, ZHANG A M, LIANG H W, LÜ H Y, DENG X D, GE Y Q, WEI X, YANG W C. Current status and perspective of wheat genomics, genetics, and breeding., 2018, 19(3): 430-434. (in Chinese)
[3] GENG H W, XIA X C, ZHANG L P, QU Y Y, HE Z H. Development of functional markers for a lipoxygenase geneon chromosome 4BS in common wheat., 2012, 52: 568-576.
[4] 何中虎, 夏先春, 陈新民, 庄巧生. 中国小麦育种进展与展望. 作物学报, 2011, 37(2): 202-215.
HE Z H, XIA X C, CHEN X M, ZHUANG Q S. Progress of wheat breeding in China and the future perspective., 2011, 37(2): 202-215. (in Chinese)
[5] 周素梅, 王强, 张晓娜. 小麦中功能性多糖阿拉伯木聚糖研究进展. 核农学报, 2009, 23(2): 297-301.
ZHOU S M, WANG Q, ZHANG X N. Research advances in functional polysaccharide-wheat arabinoxylan., 2009, 23(2): 297-301. (in Chinese)
[6] DELCOUR J A, VANHAMEL S, HOSENEY R C. Physicchemical and functional properties of rye non-starch polysaccharides: II. The impact of a fraction containing water-soluble pentosans and proteins on gluten-starch loaf volumes., 1991, 68: 72-76.
[7] IZYDORCZYK M S, BILIADERIS C G. Cereal arabinoxylans advances in structure and physicochemical properties., 1995, 28: 33-48.
[8] 袁小平, 王静, 姚惠源. 小麦麸皮阿魏酰低聚糖对红细胞氧化性溶血抑制作用的研究. 中国粮油学报, 2005, 20(1): 13-16.
YUAN X P, WANG J, YAO H Y. A study on the inhibition of erythrocyte oxidative hemolysis in wheat bran., 2005, 20(1): 13-16. (in Chinese)
[9] BONTPART T, CHEYNIER V, AGEORGES A, TERRIER N. BAHD or SCPL acyltransferase? What a dilemma for acylation in the world of plant phenolic compounds., 2015, 208: 695-707.
[10] 刘亚男, 夏先春, 何中虎. 普通小麦基因克隆与特异性标记开发. 作物学报, 2013, 39(4): 589-598.
LIU Y N, XIA X C, HE Z H.Characterization of dense and erect panicle 1 gene () located on common wheat group 5 chromosomes and development of allele-specific markers., 2013, 39(4): 589-598. (in Chinese)
[11] 高华利, 王黎明, 柴军琳, 董普辉, 王春平, 李兴锋. 小麦籽粒品质性状基因功能标记的开发及应用. 中国粮油学报, 2016, 31(8): 152-157.
GAO H L, WANG L M, CHAI J L, DONG P H, WANG C P, LI X F. Development and application of gene functional markers of grain quality traits in wheat., 2016, 31(8): 152-157. (in Chinese)
[12] BURR S J, FRY S C. Feruloylated arabinoxylans are oxidatively cross-linked by extracellular maize peroxidase but not by horseradish peroxidase., 2009, 2: 883-892.
[13] MARTINANT J P, CADALEN T, BILLOT A, CHARTIER S, LEROY P, BERNARD M, SAULNIER L, BRANLARD S G. Genetic analysis of water extractable arabinoxylans in bread wheat endosperm., 1998, 97: 1069-1075.
[14] RAO R S P, MURALIKRISHNA G. Non-starch polysaccharide- phenolic acid complexes from native and germinated cereals and millet., 2004, 84: 527-531.
[15] RAO R S P, MURALIKRIAHNA G. Structural characteristics of water-soluble feruloyl arabinoxylans from rice () and ragi (finger millet,): variations upon malting., 2007, 104: 1160-1170.
[16] ORDAZ-ORTIZ J J, DEVAUX M F, SAULNIER L. Classification of wheat varieties based on structural features of arabinoxylans as revealed by endoxylanase treatment of flour and grain., 2005, 53: 8349-8356.
[17] 张岐军, 钱森和, 张艳, 何中虎, 姚大年. 中国软质小麦品种戊聚糖含量的遗传变异及其与饼干加工品质的关系. 中国农业科学, 2005, 38(9): 1734-1738.
ZHANG Q J, QIAN S H, ZHANG Y, HE Z H, YAO D N. Variation of pentosans in Chinese soft wheat cultivars and correlations with cookie quality., 2005, 38(9): 1734-1738. (in Chinese)
[18] 钱森和, 张艳, 王德森, 何中虎, 张歧军, 姚大年. 小麦品种戊聚糖和溶剂保持力遗传变异及其与品质性状关系的研究. 作物学报, 2005, 31(7): 902-907.
QIAN S H, ZHANG Y, WANG D S, HE Z H, ZHANG Q J, YAO D N. Variation of pentosans and solvent retention capacities in wheat genotypes and their relationship with processing quality., 2005, 31(7): 902-907. (in Chinese)
[19] 刘雨雨, 莫婷, 王晓晖, 史社坡, 刘晓, 屠鹏飞. 植物来源BAHD酰基转移酶家族研究进展. 中国中药杂志, 2016, 41(12): 2175-2182.
LIU Y Y, MO T, WANG X H, SHI S P, LIU X, TU P F. Research progress of plant BAHD acyltransferase family., 2016, 41(12): 2175-2182. (in Chinese)
[20] MITCHELL R A C, DUPREE P, SHEWRY P R. A novel bioinformatics approach identifies candidate genes for the synthesis and feruloylation of arabinoxylan., 2007, 144: 43-53.
[21] OHYANAGI H, TANAKA T, SAKAI H, SHIGEMOTO Y, YAMAGUCHI K, HABARA T, FUJII Y, ANTONIO B A, NAGAMURA Y, IMANISHI T, IKEO K, ITOH T, GOJOBORI T, SASAKI T. The rice annotation project database (): hub forssp.genome information., 2006, 34: 741-744.
[22] TANAKA T, ANTONIO B A, KIKUCHI S, MATSUMOTO T, NAGAMURA Y, NUMA H, SAKAI H, WU J, ITOH T, SASAKI T, AONO R, FUJII Y, HABARA T, HARADA E, KANNO M, KAWAHARA Y, KAWASHIMA H, KUBOOKA H, MATSUYA A, NAKAOKA H, SAICHI N, SANBONMATSU R, SATO Y, SHINSO Y, SUZUKI M, TAKEDA J, TANINO M, TODOKORO F, YAMAGUCHI K, YAMAMOTO N, YAMASAKI C, IMANISHI T, OKIDO T, TADA M, IKEO K, TATENO Y, GOJOBORI T, LIN Y C, WEI F J, HSING Y, ZHAO Q, HAN B, KRAMER M R, MCCOMBIE R W, LONSDALE D, O’DONOVAN C C, WHITFIELD E J, APWEILER R, KOYANAGI K O, KHURANA J P, RAGHUVANSHI S, SINGH N K, TYAGI A K, HABERER G, FUJISAWA M, HOSOKAWA S, ITO Y, IKAWA H, SHIBATA M, YAMAMOTO M, BRUSKIEWICH R M, HOEN D R, BUREAU T E, NAMIKI N, OHYANAGI H, SAKAI Y, NOBUSHIMA S, SAKATA K, BARRERO R A, SATO Y, SOUVOROV A, SMITH-WHITE B, TATUSOVA T, AN S Y, AN G, OOTA S, FUKS G, MESSING J, CHRISTIE K R, LIEBERHERR D, KIM H, ZUCCOLO A, WING R A, NOBUTA K, GREEN P J, LU C, MEYERS B C, CHAPARRO C, PIEGU B, PANAUD O, ECHEVERRIA M. The rice annotation project database (): 2008 update., 2008, 36: 1028-1033.
[23] MATSUMOTO T, TANAKA T, SAKAI H, AMANO N, KANAMRI H, KURITA K, KIKUTA A, KAMIYA K, YAMAMOTO M, IKAWA H, FUJII N, HORI K, ITOH T, SATO K. Comprehensive sequence analysis of 24,783 barley full-length cDNA derived from 12 clone libraries., 2011, 156: 20-28.
[24] LI Y, ZHAO D H, JUN Y, ZHANG Y L, XIA, X C, TIAN Y B, HE Z H, ZHANG Y. QTL mapping of grain arabinoxylan contents in common wheat using a recombinant inbred line population., 2016, 208: 205-214.
[25] 吕文娟, 刘金栋, 白璐, 曲延英, 任毅, 闻伟鄂, 耿洪伟. 小麦籽粒阿魏酰阿拉伯木聚糖含量QTL定位. 分子植物育种, 2018, 16(5): 1530-1538.
Lü W J, LIU J D, BAI L, QU Y Y, REN Y, WEN W E, GENG H W. QTL mapping and genome-wide association study of Ferulic acid Arabiaxylan content in wheat grain., 2018, 16(5): 1530-1538. (in Chinese)
[26] GRIENENBERGER E, BESSEAU S, GEOFFROY P, DEBAYLE D, HEINTE D, LAPIERRE C, POLLET B, HEITZ T, LEGRAND M. A BAHD acyltransferase is expressed in the tapetum ofanthers and is involved in the synthesis of hydroxycinnamoyl spermidines., 2009, 58: 246-259.
[27] BEAUGRAND J, CONIER D, DEBEIRE P, CHABBERT B. Arabinoxylan and hydroxycinnamatecontent of wheat bran in relation to endoxylanase susceptibility., 2004, 40: 223-230.
[28] 杨莉, 黄玉莲, 常萍, 阎俊, 张业伦, 夏先春, 田宇兵, 何中虎, 张勇. 小麦阿拉伯木聚糖含量的QTL分析及其与品质性状的关系. 作物学报, 2014, 40(9): 1695-1701.
YANG L, HUANG Y L, CHANG P, YAN J, ZHANG Y L, XIA X C, TIAN Y B, HE Z H, ZHANG Y. QTL mapping for arabinoxylans content and its relationship with processing quality in common wheat., 2014, 40(9): 1695-1701. (in Chinese)
[29] 李冰, 张照贵, 王佳佳, 李斯深. 小麦基因克隆及其功能标记开发. 山东农业科学, 2014, 46(10): 6-11.
LI B, ZHANG Z G, WANG J J, LI S S. Development of wheatgene cloning and its functional markers., 2014, 46(10): 6-11. (in Chinese)
[30] HUANG S, MORRISON W R, Aspects of protein in Chinese and British common () wheats related to quality of white and yellow Chinese noodles., 1988, 8:177-187.
[31] 李亚青, 毛新国, 赵宝存, 葛荣朝, 沈银, 黄占景. 小麦糖原合成酶激酶基因()的染色体定位. 华农学报, 2006, 21(5): 39-41.
LI Y Q, MAO X G, ZHAO B C, GE R Z, SHEN Y, HUANG Z J. Chromosome localization of the gene of wheat glycogen synthase kinase ()., 2006, 21(5): 39-41. (in Chinese)
[32] HESSLER T G, THOMSON M J, BENSCHER D, NACHIT M M, SORRELLS M E. Association of a lipoxygenase locus,, with variation in lipoxygenase activity in durum seeds., 2002, 42: 1695-1700.
[33] SUN D J, HE Z H, XIA X C, ZHANG L P, MORRIS C F, APPELS R, MA W J, WANG H. A novel STS marker for polyphenol oxidase activity in bread wheat., 2005, 16: 209-218.
[34] CARRERA V, ECHENIQUE W, ZHANG M, HELGUERA F, MANTHEY A, SCHRAGER0 A, PICCA G, CERVIGNI J, DUBCOVSKY. A deletion at thelocus is associated with low lipoxygenase activity and improved pasta color in durum wheat., 2007, 45: 67-77.
[35] SOMERS D J, ISAAC P, EDWARDS K. A high-density microsatellite consensus map for bread wheat (L.)., 2004, 109: 1105-1114.
[36] D’AURIA J C. Acyltransferases in plants: a good time to be BAHD., 2006, 9: 331-340.
[37] YU X H, GOU J Y, LIU C J. BAHD superfamily of acyl-CoA dependent acyltransferases inand: bioinformatics and gene expression., 2009, 70: 421-442.
[38] PENG M, GAO Y Q, CHEN W, WANG W S, SHEN S Q, SHI J, WANG C, ZHANG Y, ZOU L, WANG S C, WANG J, LIU X Q, GONG L, LUO J. Evolutionarily distinct BAHD N-acyltransferases are responsible for natural variation of aromatic amine conjugates in rice., 2016, 28: 1533-1550.
[39] TREMMEL B K, LANG L, TOROK K, TOMOSKOZI S, VIDA G, SHEWRY P R, BEDO Z, RAKSZEGI M. Development and characterization of wheat lines with increased levels of arabinoxylan., 2017, 213: 291-306.
[40] WEI B, JING R L, WANG C S, CHEN J B, MAO X G, CHANG X P, JIA J Z.genes in wheat (L.): development of functional markers, gene mapping based on SNPs., 2009, 23: 13-22.
[41] SMITH P J, WANG H T, WILLIAM S, MARIA J, BREEANNA R. Designer biomass for next-generation biorefineries: leveraging recent insights into xylan structure and biosynthesis., 2017, 10: 286-300.
[42] EUDES A, MOUILLE M, ROBINSON D S, BENITES V T, WANG G, ROUX L, TSAI Y L, BAIDOO E E K, CHIU T Y, HEAZLEWOOD J L, SCHELLER H V, MUKHOPADHYAY A, KEASLING J D, DEUTSCH S, LOQUÉ D. Exploiting members of the BAHD acyltransferase family to synthesize multiple hydroxycinnamate and benzoate conjugates in yeast., 2016, 15: 198-214.
(责任编辑 李莉)
Arabinoxylan feruloyl transferase gene cloning and development of functional markers in common wheat
ZHAN ShuaiShuai1, BAI Lu2, XIE Lei1, XIA XianChun3, REN Yi1, Lü WenJuan1, QU YanYing1, GENG HongWei1
(1College of Agronomy, Xinjiang Agricultural University/Laboratory of Agricultural Biological Technology, Urumqi 830052;2College of science and technology, xinjiang agricultural university, urumqi 830091;3Institute of Crop Science, Chinese Academy of Agricultural Sciences/National Wheat Improvement Center, Beijing 100081)
【Objective】 The arabinoxylan feruloyl transferase (AFT) gene was cloned from wheat, and the functional markers linked to the content of FAX were developed to improve the accuracy of predicting the content of FAX , in order to provide the basis for the improvement of wheat processing quality. 【Method】 The complete gDNA sequence of wheat FAX gene was obtained by homologous cloning method,using FR846233 as a probe. The sequence differences between high and low FAX content varieties were compared by the DNAMAN software; based on sequence difference, specific primers were designed with Primer5.0 software to develop functional markers closely linked to FAX content and a set of Chinese spring Nullisomic-tetrasomic lines and ditelosomic line 3AS, and 3AL were used for chromosome physical mapping; and the practicability of functional markers were verified by using 253 wheat varieties (lines) from the main winter wheat regions in China, combined with the method of PCR verification. IBM SPSS statistics 19.0 software was used to analyze the correlation between FAX content and genotypes. 【Result】Two pairs of specific primers B1 and B2 finally amplified fragments of 800 bp and 710 bp, respectively. And 80 bp overlaps of PCR fragments amplified by B1 and B2 were spliced to obtain the AFT gene that is located on chromosome 3A. Thesequence is consists of 1 429 base pairs and allelic variantsandare obtained. The two allelic variants possess a 1 266 bp open reading frame, two exons and one intron. The introns conformed to the typical GT-AG structure. The similarity between the alleles was 98.08%, with 24 SNPs and 3 InDels, which could encode 421 amino acid residues and the predicted molecular weight was 45.2 kDa. Two complementary dominant markers AFTA2 and AFTB2 were developed based on the 107 bp SNP. AFTA2 was able to amplify a 692 bp fragment in thematerial, which was associated with a high FAX content, but not in the material withallelic variation. AFTB2 could only amplify a 438 bp fragment intype material and correlate with low FAX content, but not intype material, and AFTA2 and AFTB2 were located in 3AL chromosome of wheat by a set of Chinese spring Nullisomic-tetrasomic lines. Using the functional markers AFTA2 and AFTB2 to detect 253 Chinese winter wheat materials, the results showed that the difference in FAX content of different genotypes reached a significant level (<0.05), and there was no significant difference in FAX content in Northern China Plain Winter Wheat Region; but in the Huang-Huai River Valley Winter Wheat Region, The FAX content of the-containing variety was significantly higher than that of the- containing variety (<0.05). Therefore, the complementary dominant markers AFTA2 and AFTB2 are related to FAX content and can be effectively used for genetic improvement of FAX content. The frequency ofallele variation indicates thatis an excellent allelic variant associated with high FAX content, and the frequency ofin the Northern China Plain Winter Wheat Region (71.3%) is significantly higher than that in the Huang-Huai River Valley Winter Wheat Region (60.2%). 【Conclusion】 The results suggested that these two STS markers are closely related to FAX content related gene and could be used for the improvement of wheat processing quality for wheat-based products.
common wheat; feruloyl arabinoxylan; gene cloning; functional marker development
2018-04-21;
2018-05-25
国家自然科学基金(31771786)、自治区青年科技创新人才培养工程(QN2016JQ0025)
战帅帅,Tel:17690762406;E-mail:1269047935@qq.com。 通信作者耿洪伟,Tel:13579873801;E-mail:hw-geng@163.com
10.3864/j.issn.0578-1752.2018.19.002