不同时期葡萄各器官碳氮分配差异
2018-10-07刘光春王亚芳陈征文翟衡杜远鹏
刘光春,王亚芳,陈征文,翟衡,杜远鹏*
(山东农业大学园艺科学与工程学院,山东泰安 271018)
光合产物在树体内的运输、分配情况是影响果树优质、丰产、稳产的重要因素[1]。碳、氮代谢是植物体内最主要的两大代谢过程[2],因此研究植株体内碳氮营养的分配,对于提高植株的生产性意义重大。碳氮示踪技术是研究光合产物和氮素营养固定吸收分配的有效手段[3-7]。Farmer[8]首次将稳定性碳同位素13C用于植物的碳代谢研究。相关学者应用13C和15N示踪技术在苹果、桃等果树的碳氮营养研究方面已经取得了一定成果[9-13],而利用13C和15N示踪技术研究酿酒葡萄的碳氮营养吸收、分配的相关报道较少,由于葡萄的架式和栽培方式不同于其他果树,并且还具有枝条生长量大、副梢发生级次多、夏季修剪工作量大的特点。为此,本试验中以酿酒葡萄‘赤霞珠’为试材,在果实发育的3个重要时期对其新梢上部叶片进行标记,旨在研究新梢上部叶片固定的光合产物及吸收氮素营养在不同时期的主要分配方向,为研究葡萄碳氮营养吸收分配规律提供理论依据。
1 材料与方法
1.1 试材与处理
试验于2015年在山东农业大学实验站葡萄园进行,株行距0.8 m×2.0 m,试材均为7年生‘赤霞珠’,篱架,单干单臂树形。
试验2:标记物在植株中的运转速率探讨。于果实转色期选取长势基本一致、无病害植株6株,选取植株水平主蔓上第一发枝部位的新梢,分别对其上部叶片进行13C和15N的双标记,一组标记3 d后破坏性整株取样,另一组标记9 d后破坏性整株取样,单株小区,重复3次。
1.2 标记方法与取样
13C脉冲标记在一个由透明农用薄膜做成的标记室内进行[14],标记前先检查标记室的密封性[15]。用注射器向装有0.2 g(或0.6 g)Ba13CO3的离心管中注入一定量1 mol/L的盐酸溶液。此后每隔0.5 h向其中注入1次盐酸,以维持CO2浓度,标记时间持续4 h。用毛笔蘸0.4%的15N尿素溶液12.5 mL涂抹正反面叶片进行15N标记。72 h后破坏性全株取样。样品按清水→洗涤剂→清水→1%盐酸→3次去离子水顺序冲洗后,105 ℃下杀青30 min,随后在80 ℃下烘干至恒重,粉碎后过0.25 mm筛,混匀后装袋备用[16]。
1.3 测定与计算
13C丰度用DELTAplusXP型质谱仪(Thermo Fisher Scientific,inc., uSA)在中国林科院测定,15N丰度用ZhT-03质谱计(北京分析仪器厂)在中国农业科学院原子能研究所测定。
13C含量:13C/‰=[Ci×(Fi-Fnl)/100]×1000,式中Ci为各组分所含的碳量(g),nl表示未标记。
13C分配率:13Ci/%=(13C/13C净吸收)×100。
15N计算。Ndff指植株器官从肥料中吸收分配到的15N量对该器官全氮量的贡献率,反映了植株器官对肥料15N的吸收征调能力。
Ndff/%=[(样品中的15N丰度-自然丰度)/(肥料中15N的丰度-自然丰度)]×100;
另一方面,“一带一路”背景下的高等教育国际化发展又面临着一些新的问题。如需进一步深化课程教学的改革,将国际意识和能力的强化渗透到课程教学与科研当中,将“一带一路”国家和地区的有关情况作为应用情景,培养学生运用知识和能力去分析和处理国际事务的能力。又如建立国际化的师资队伍,引入国外师资力量,提升国内教师的国际化视野等等。如何“吐故纳新,中外结合”将会是我国高等教育改革的一大挑战。
15N分配率:15N/%=各器官从氮肥中吸收的氮量(g)/总吸收氮量(g)×100;
总N量/g=干物质量(g)×植株氮浓度N(%)。
所有数据均采用DPS 7.05软件进行单因素方差法分析,Tukey法进行差异显著性比较,应用Microsoft Excel 2003进行图表绘制。
2 结果与分析
2.1 标记时期对各器官碳氮营养吸收分配的影响
2.1.1 不同标记时期植株各器官的13C含量和分配率
从表1可以看出,葡萄上部叶片固定的光合产物其叶片自留量无论在果实发育的哪个时期均稍高于30%(30.64%~32.49%),剩余近70%向其他器官转运。其中,果实转色期、成熟期标记新梢的果实中光合产物分配率较膨大期的高,分别是膨大期的4.03倍、2.90倍;而且上部叶片固定的光合产物在3个果实发育关键时期均未向粗根分配,在膨大期和转色期有极少的光合产物向细根分配,而且转色期的细根分配率是膨大期的3.33倍;随着葡萄果实生育期的发展,标记新梢的碳素自留率不断增加,即成熟期的最高,高达86.77%,转色期次之,膨大期最低。
2.1.2 标记时期葡萄植株各器官的15N含量和分配率
从表2可以看出,葡萄上部叶片自留的氮营养比率比碳素营养稍高,而且3个标记时期自留的氮营养量不同,成熟期最高,膨大期次之,转色期的自留量最低。其中成熟期标记新梢各器官获得的氮营养总量高达90%;标记新梢上果实的分配率表现为:成熟期>膨大期>转色期;膨大期和转色期有较少的氮营养分配给细根,成熟期没有分配,但膨大期细根的氮营养分配率是转色期的1.37倍,粗根则在各个时期依然没有分配到氮营养。
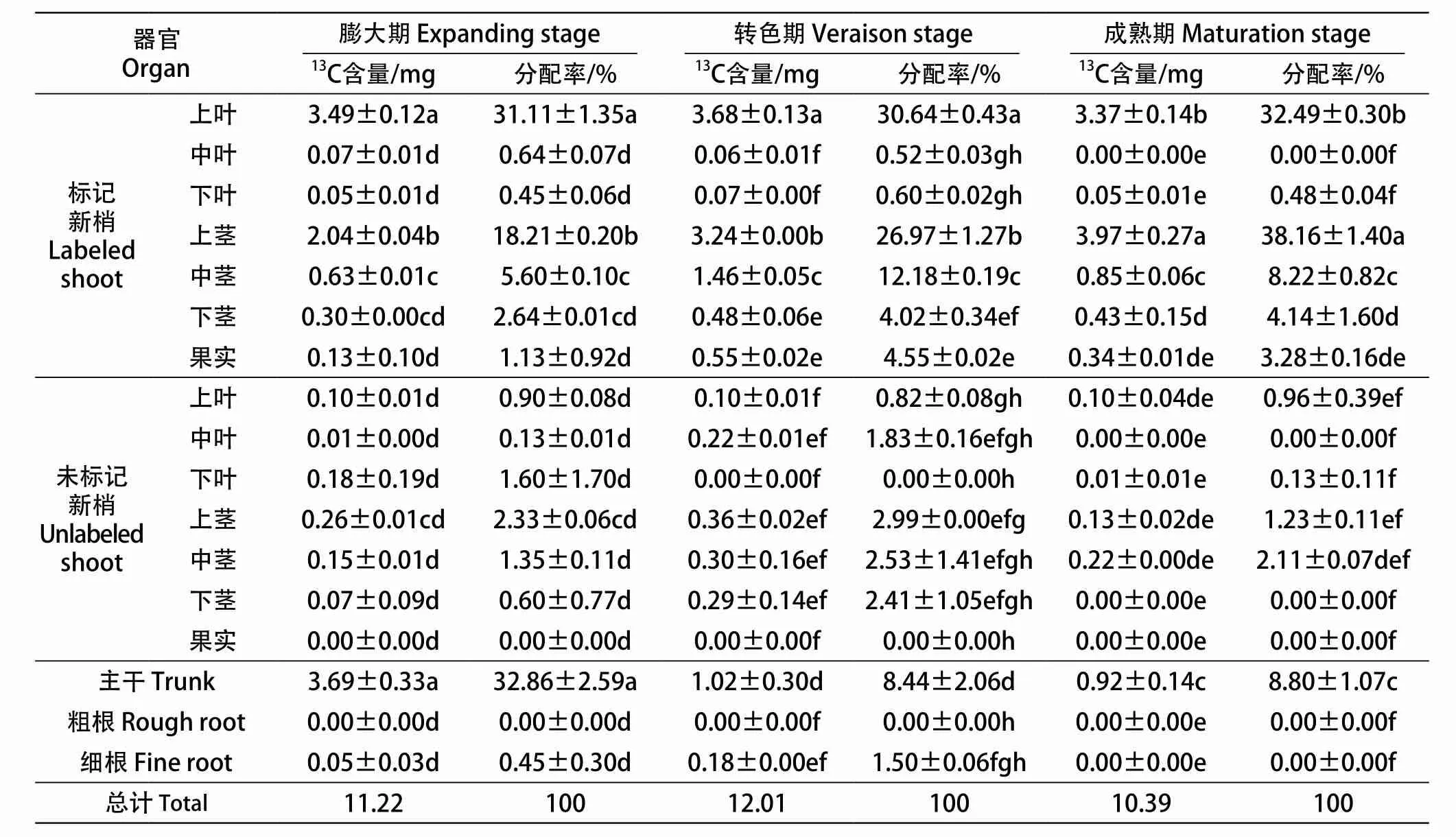
表1 不同标记时期葡萄植株各器官的13C含量和分配率Table 1 The 13C content and distribution rate of different organs at different labeling stage
2.1.3 不同标记时期葡萄植株各器官的Ndff值
从表2中可以看出,受标记新梢的上部叶片Ndff值无论在哪个发育时期都显著高于未标记器官,且随着果实成熟有增加趋势。相反,未标记的中部叶片对上部标记叶片氮营养的竞争能力随着生育期的延长不断降低,而未标记的下部叶片对上部标记叶片氮营养的竞争能力以果实膨大期最高,成熟期次之,转色期最低。转色期未标记新梢各器官Ndff值总量在果实3个发育时期中占比最高,果实膨大期次之,而果实成熟期最低,说明不同时期,未标记新梢对相邻新梢上部叶片的氮营养竞争能力不同,而且均较低。
2.2 13C、15N双标记在植株中的运转速率
2.2.113C在不同器官中的转运及分配率
从表3中可以看出,标记3 d后和标记9 d后上部叶片自留的碳营养基本一致,均有70%左右向其他器官转运。标记9 d后,标记新梢向果实和细根的分配率高于标记3 d的,是其1.65和13.64倍,而主干的分配率下降至标记3 d的51.37%。说明在果实转色期随着标记时间的延长,葡萄上部叶片固定的碳营养途径主干的碳营养继续向果实和细根这两个重要的库分配。
2.2.215N在不同器官中的转运分配率
标记3 d后和标记9 d后上部叶片自留的氮营养基本一致,约有一半的氮营养向其他器官转运(表4),与上部叶片自留的碳营养相比,上部叶片自留的氮营养高于碳营养。标记9 d后标记新梢的果实和细根的分配率均高于标记3 d后的,分别是其的2.49和2.23倍;这说明,在葡萄转色期,果实和细根是作为植株两个重要的库器官,上部叶片吸收的氮营养向库的分配量随着标记时间的延长而增加。
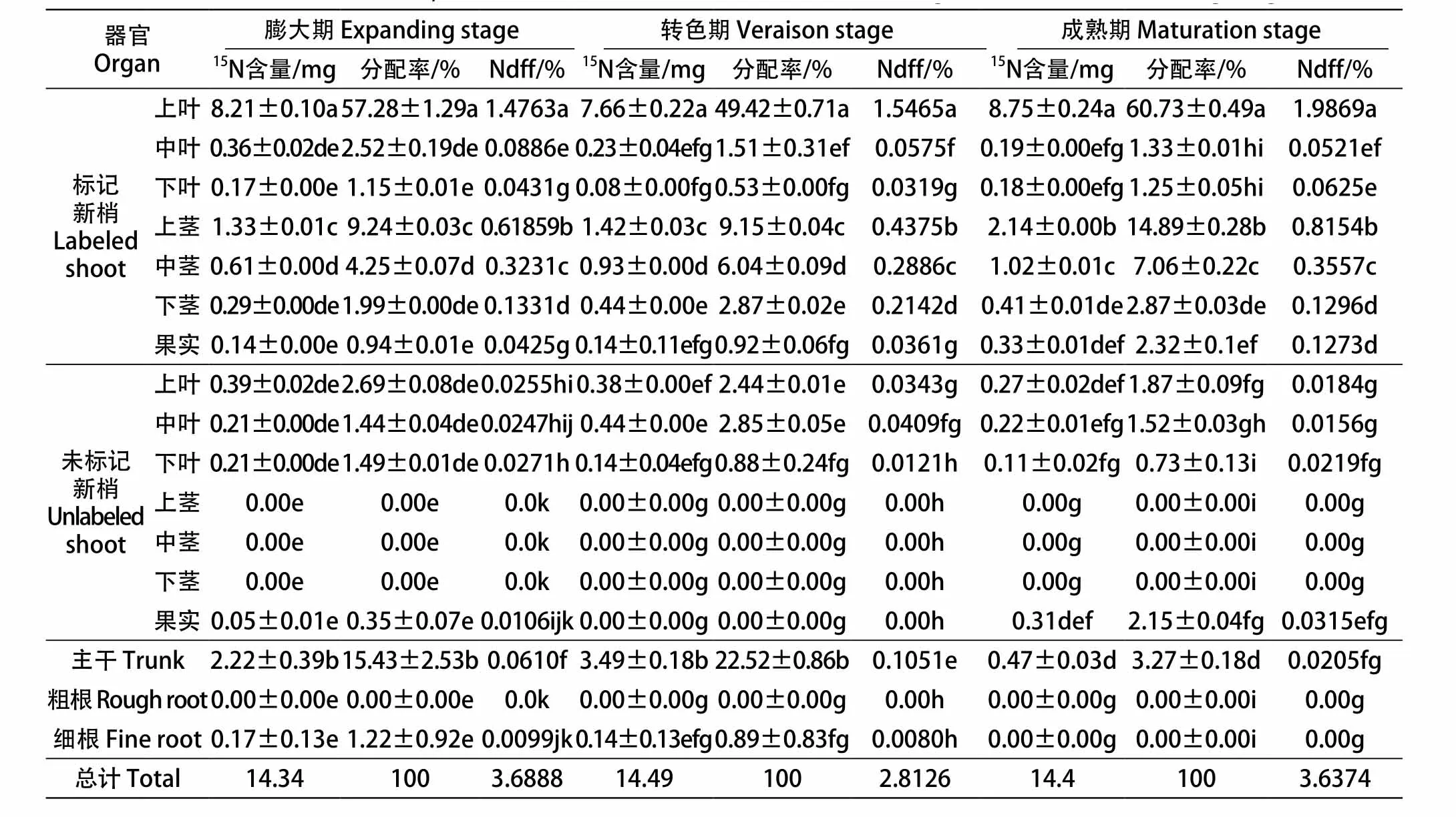
表2 不同标记时期葡萄植株各器官15N的含量和分配率以及Ndff%值比较Table 2 The 15N content, distribution rate and Ndff% of different organs at different labeling stage
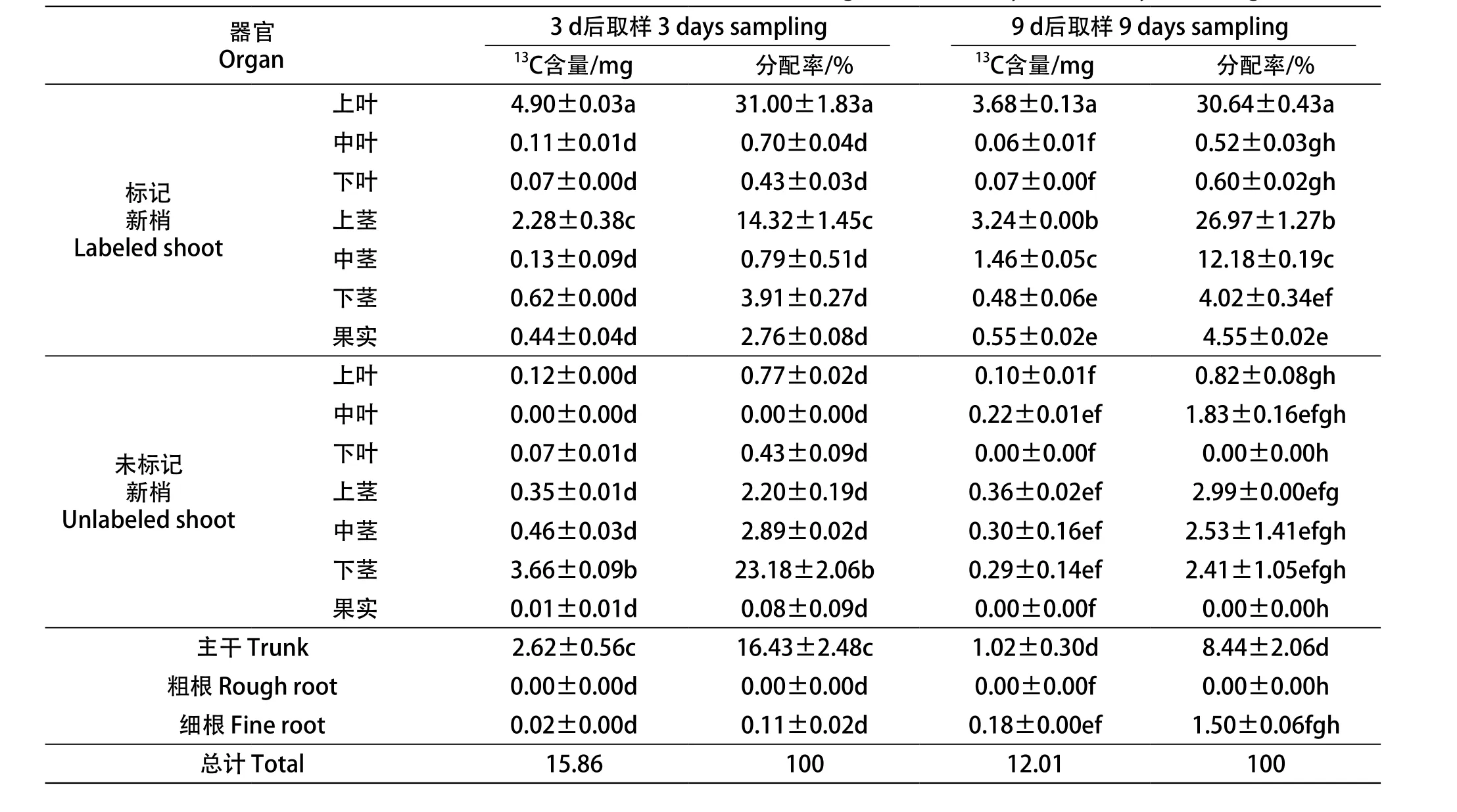
表3 标记3 d和9 d后各器官13C含量和分配率Table 3 The 13C content and distribution rate of different organs after 3 days and 9 days labeling

表4 标记3 d后和9 d后各器官15N含量和分配率以及Ndff值Table 4 The 15N content, distribution rate and Ndff% of different organs after 3 days and 9 days labeling
2.2.3 标记后3 d和9 d各器官Ndff值
从表4中可以看出,标记新梢的上部叶片及其未标记的其他器官(中部和下部叶片、果实、主干和细根)氮的征调能力随着时间的延长均有所上升,氮的分配率也随之上升。而且无论标记3 d还是9 d后,未标记上部叶片的新梢各器官Ndff值总和均低于标记上部叶片的新梢,分别是其的11.17%和3.34%。这说明新梢对于其他新梢上部叶片的氮营养的征调能力很低,而且随着时间的延长,征调能力还在不断降低。
3 讨论与结论
从试验结果可以看出,上部叶片固定的碳营养约有30%(30.64%~32.49%)自留,吸收的氮营养约有50%左右(49.42%~60.73%)自留,用于自身的生长发育,其自留比例稍低于焦培娟等[17]的研究结果,其在山葡萄的研究认为,上部枝和中部枝标记处理饲喂本枝的14C同化产物绝大部分滞留用于自身枝条建设,而下部枝饲喂枝主要是输送到下部的主蔓和根系中。这可能与品种类型有关,如杂交稻将更多的氮转移到稻谷中,而常规稻则贮藏到了稻草中[18]。本研究还发现,上部叶片自留的氮营养高于碳营养,除上部叶片自留的碳氮营养,剩下的均向其他器官分配,不同转运时间和不同时期,各器官的分配率均有所差异,这与刘洪家等[19]在黑穗醋栗上研究结果一致。
不同时间采样测定可以看出,碳氮营养分配率有随着转运时间的延长而提高的趋势,这说明3 d的转运时间对于7年生的‘赤霞珠’大树完成体内的光合产物和氮素营养的分配尚不充分,需要更长时间进一步完成转运,以达到标记产物在树体内的稳定。不同器官标记新梢的碳营养分配率总量上升,细根的碳营养分配率上升;未标记新梢的碳营养分配率总量下降,主干的碳营养分配率下降。标记新梢的氮营养的分配率总量上升,主干和细根的氮营养分配率均有所提高,未标记新梢的氮营养分配率下降,这些现象也可能与再分配有关,还需进一步研究证实。植物碳氮营养分配遵循由源到库的原则,源和库是相对的,在不同的生育期有着不同的源和库。从果实膨大期到成熟期果实中氮的分配率迅速上升[20]。本试验在果实成熟期,标记新梢的碳氮营养的分配率总量均最高,并且均没有向细根分配,未标记新梢和主干的碳氮营养分配均较低;膨大期和转色期在碳氮营养分配上有很大不同,膨大期果实迅速生长膨大,是具有较强竞争力的库,因此所获得的碳营养分配高于转色期,新梢无论标记与否其碳营养分配率总量均低于转色期,细根的碳营养分配率也低于转色期,而对于氮营养,膨大期标记新梢的氮营养分配率和细根氮营养的分配率均高于转色期,未标记新梢的氮营养分配率低于转色期,主干的分配率亦低于转色期。
研究发现,光合产物在各器官的分配率与该器官重量关系,有些器官因其在全株占的比重较大,造成分配率较高,但活度单位数却很小,说明该器官并非光合产物分配中心[21]。本试验中标记新梢的茎的分配率均比较高,但茎中的韧皮部是碳氮营养的运输组织,所以碳氮营养一方面用于新梢自身生长,另一方面也有一部分是在往其他器官的运输途中,尤其是3 d转运时间的主干的碳营养显著高于转运9 d的,这可能是由于3 d时间太短,还有部分碳氮营养在转运途中,增加转运时间,其中的碳氮营养继续向其他器官分配。深入了解植株的碳氮营养分配特征对指导树形构建、新梢管理和科学施肥均有科学意义。
