栎黄枯叶蛾低龄幼虫空间分布的地统计学分析
2018-09-19刘永华郑羽墨阎雄飞陆鹏飞
刘永华,郑羽墨,阎雄飞,贺 英,陆鹏飞
1 榆林学院生命科学学院,榆林 719000 2 省部共建森林培育与森林保护教育部重点实验室,北京 100083
空间格局是昆虫种群的一个重要特征,它是在昆虫的生物学特性和生境条件的共同作用下所呈现出的分布形式或散布状况[1]。研究昆虫种群的空间格局,对于详细了解昆虫的生物生态学特性、揭示种群消长规律、提高抽样技术精度、害虫监测和防治等均具有重要的理论和实践意义[2- 4]。利用地统计学方法研究昆虫种群空间格局,可以充分利用野外调查所获得的空间数据,最大限度避免系统误差,更深入的描述种群的空间分布特征,相比传统生物学统计方法随机选取的理论体系,地统计学方法同时考虑了样本值大小与样本的空间方位及其相互距离的关系,能够更为精确的揭示空间相关性和依赖性,已成为昆虫生态学领域的研究热点[5-6]。
沙棘Hippophaerhamnoides是我国西北干旱半干旱地区生态建设的先锋树种,并且在地方经济发展中具有重要的作用。尤其是陕北吴起县沙棘种植面积为全国之最,成林面积已达到12.53万hm2。栎黄枯叶蛾TrabalavishnougigantinaYang,属鳞翅目Lepidoptera枯叶蛾科Lasioeampidae,寄主种类众多,分布范围较广[7]。该虫近年在吴起沙棘林中为害特别严重,幼虫往往将叶片全部吃光,整片沙棘林仅剩枝干,严重影响沙棘生长,对沙棘的生态价值和经济效益产生巨大的破坏[8],因此,研究该虫在沙棘上危害的种群动态及空间分布格局,对于制定防治措施具有重要意义。
章一巧等[9- 10]应用地统计学方法对栎黄枯叶蛾蛹和卵的空间分布进行了研究,认为栎黄枯叶蛾卵在沙棘林的分布均呈聚集分布,而蛹在沙棘的密林中呈聚集分布,在疏林中呈随机分布;卵同雌蛹在分布上有一定的跟随性,蛹在沙棘林上的分布存在边缘效应,且方向性明显。幼虫是栎黄枯叶蛾为害沙棘的主要阶段,但目前对其空间分布格局研究尚未见报道。研究发现,栎黄枯叶蛾低龄幼虫取食量较低,往往群集于孵化位置附近取食,3龄后食量突然增加危害加重[11],因此对其低龄幼虫的空间分布格局进行探讨,将有助于掌握幼虫在林间的分布规律,提高对幼虫危害盛期进行预测预报的准确性,以便及时采取防控措施。
1 材料与方法
1.1 试验地概况
试验地设在陕西省吴起县(36°33′—37°24′ N,107°38′—108°32′ E),属典型的半干旱温带大陆性季风气候,年平均气温7.8℃,年均降水量478.4 mm,无霜期较短,平均146 d,海拔1233—1809 m。土壤类型以黄绵土为主,质地多为轻壤。林地均为人工栽植的沙棘纯林,间或有少量的山杏、榆树、刺槐、山桃。
调查时间为2015年5—8月,具体调查地点为吴起县吴仓堡乡。由于当地属于黄土高原丘陵沟壑区,地势起伏多变,再加上沙棘林经过十余年的自然演变,其疏密程度具有很大差异。因此本研究根据当地实际情况,将郁闭度0.30以下的林地划为疏林,郁闭度0.30以上的林地划为密林。在进行栎黄枯叶蛾低龄幼虫空间分布调查时,分别选取30 m×20 m 的样地4块,每块样地都代表不同坡向和疏密度,包括阴坡密林、阴坡疏林、阳坡密林、阳坡疏林。阴坡海拔由东南向西北呈上升趋势,光照条件东南方向最好;阳坡海拔由西北向东南呈上升趋势,光照条件西北方向最好。样地概况见表1。

表1 样地概况
1.2 调查方法
对每一样地内的沙棘进行逐株系统调查,记录沙棘的株行距和每株沙棘上的幼虫数量,将每株沙棘的相对位置记录在坐标纸上。
1.3 空间格局分析方法
1.3.1 半方差函数
半方差函数指的是区域化变量增量的方差,即变量Z(xi) 与Z(xi+h) 其增量平方的数学期望[12]。其公式为:
式中:γ(h) 是相对距离h的样本半变异函数值,N(h) 是间隔h距离的数据对 (xi,xi+h) 的对数,Z(xi) 和Z(xi+h) 分别是点xi和xi+h位置处样本的测量值,h为分隔2个样点的距离。
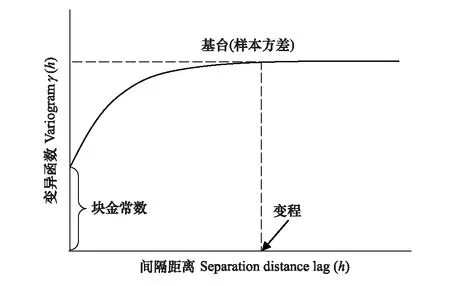
图1 典型的变异曲线图Fig.1 A typical variogram
通过半方差函数拟合生成变异曲线图(图1),图中块金常数、基台值和变程3个参数具有明显的生态学意义。这3个参数决定了半方差函数的形状和结构,并最终决定了种群的空间分布特征[10]。
1.3.2 变异函数理论模型的拟合
对于空间变异曲线的拟合主要有4种模型,即高斯模型、球状模型、指数模型和线性模型。一般来说,线性模型表示种群随机或均匀分布,即数据变化无规律,而其它模型均表示种群呈聚集分布。在应用GS+ 软件进行计算并拟合的过程中,必须根据模拟误差最小原则,首先比较各种模型的决定系数R2的大小,然后依次比较各模型残差RSS、变程A和块金常数C0的大小,最终来判断各种理论模型对半方差函数曲线进行拟合的优劣程度[13-14]。
1.3.3 空间分布图的生成
使用Kriging法对栎黄枯叶蛾低龄幼虫空间分布的区域化变量进行插值分析,通过Surfer 11软件生成空间分布等值线与矢量叠加图。
1.3.4 数据统计与分析
应用Excel 2010对基础数据进行统计分析,利用GS+ 9.0软件进行地统计学分析,采用DPS 13.5软件对数据进行方差分析(ANOVA), 差异显著性比较使用Duncan氏新复极差法。
2 结果与分析
2.1 栎黄枯叶蛾幼虫种群数量动态变化及低龄幼虫分布
调查表明,5月中旬栎叶蛾幼虫开始孵化,随着气温的升高孵化数量逐渐增多,6月上中旬进入孵化盛期,幼虫数量短时间内急剧增加。初孵幼虫聚集于孵化场所附近取食,但由于虫龄较低,取食量也不大,此时危害并不严重。6月下旬,幼虫虫龄进一步增加,开始进入暴食期,危害逐渐加重。7月下旬开始老熟,大部分幼虫7月底到8月初开始结茧化蛹,幼虫数量大幅度下降,危害程度大大降低。
图2为密林样地低龄幼虫密度分布直方图,阴坡样地和阳坡样地的有虫株率分别为90.74%和75.93%,密度>6头/株的样本比例分别为9.26%和3.70%;图3为疏林样地低龄幼虫密度分布直方图,阴坡样地和阳坡样地的有虫株率分别为92.59%和72.22%,密度>6头/株的样本比例分别为11.11%和5.56%。比较密林和疏林中低龄幼虫密度,发现阴坡样地均显著高于阳坡样地(P<0.05)。
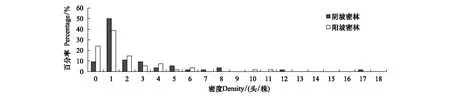
图2 密林样地低龄幼虫密度分布直方图Fig.2 Histogram of density for the early stage larvae in Dense forest
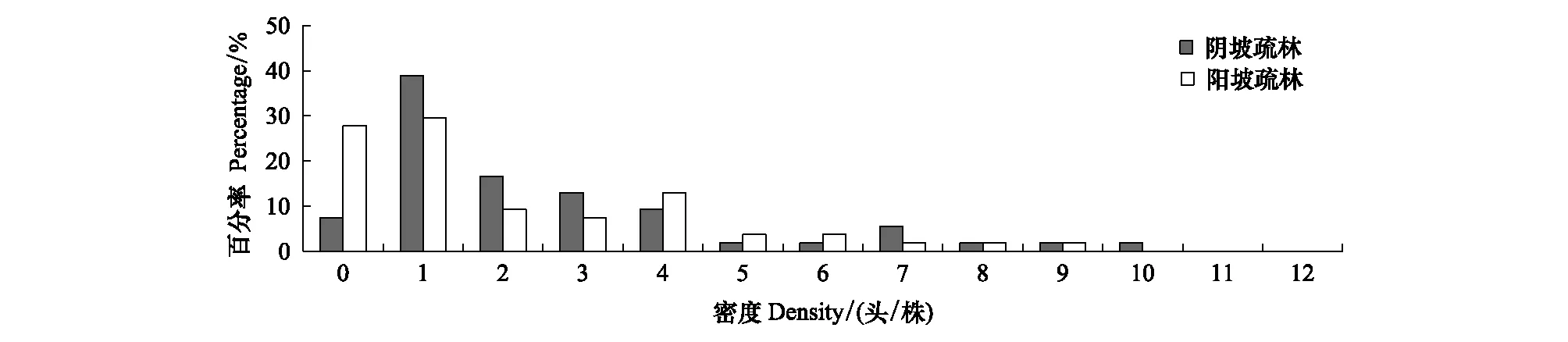
图3 疏林样地低龄幼虫密度分布直方图Fig.3 Histogram of density for the early stage larvae in Open forest
2.2 栎黄枯叶蛾低龄幼虫数量的半变异函数及空间分布分析
应用GS+9.0软件对沙棘样地中低龄幼虫的空间分布进行地统计学分析,最佳拟合模型选取标准为R2最大、RSS最小,拟合函数模型及分布格局见表2,空间变异函数见图4。
利用Surfer 11软件通过Kriging法对栎黄枯叶蛾低龄幼虫在不同坡向和林分结构的样地中沙棘位置与其幼虫数进行插值分析,生成低龄幼虫空间分布的等值线与矢量叠加图(图4)。
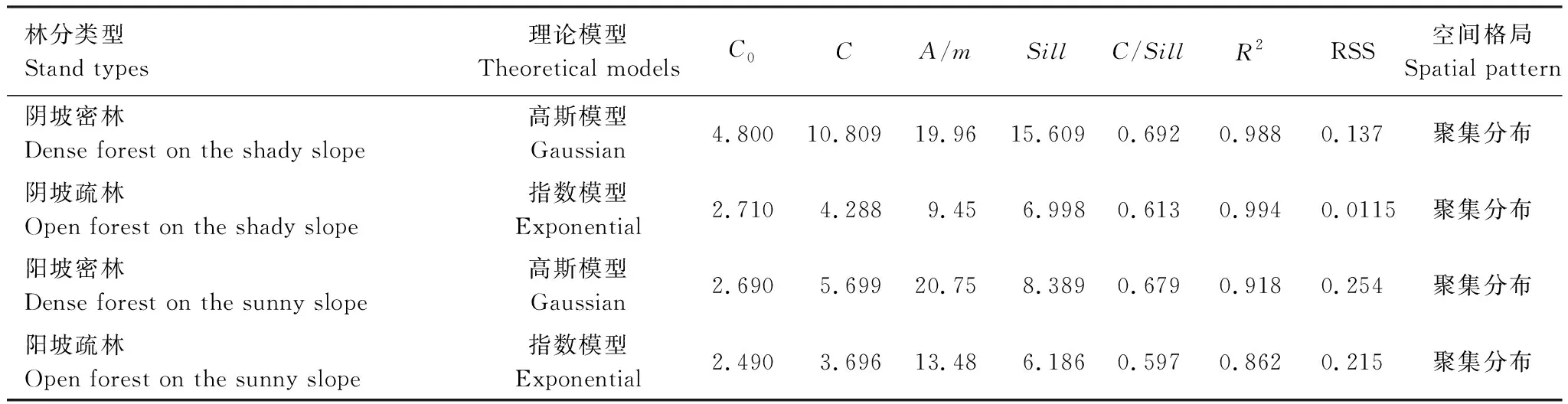
表2 栎黄枯叶蛾低龄幼虫的变异函数理论模型、拟合参数及空间分布格局
C0:块金常数,Nugget;C:偏基台值,Patial sill;A:变程,Range;Sill:基台值;C/Sill:空间变异,Spatial variation;R2:决定系数,Determination coefficient;RSS:残差平方和,Residual sum of squares

图4 栎黄枯叶蛾低龄幼虫的空间变异函数Fig.4 Spatial variograms for Trabala vishnou gigantina early stage larvae
2.2.1 密林中栎黄枯叶蛾低龄幼虫的半变异函数分析
由表2和图4可以看出,在沙棘密林中,阴坡和阳坡的栎黄枯叶蛾低龄幼虫均表现为聚集分布。阴坡和阳坡的空间依赖范围分别为19.96 m和20.75 m,其意义为在抽样空间中任意2个样点之间距离小于此范围时,其数量之间存在有一定程度的空间相关或依赖关系,当距离大于此范围时,随着距离的逐渐增大,空间相关性逐渐减小。阴坡密林中低龄幼虫的空间聚集程度略大于阳坡。阴坡和阳坡密林中的偏基台值占比分别为69.2%和67.9%,这意味着总空间变异中由自相关现象引起的有69.2%和67.9%。分析结果中块金常数较高可能是由于样地内个别沙棘的缺失导致这一区域的幼虫分布出现空白,也可能是由于统计时出现了误差。
2.2.2 疏林中栎黄枯叶蛾低龄幼虫的半变异函数分析
由表2和图4得知,在沙棘疏林中,阴坡和阳坡的栎黄枯叶蛾低龄幼虫仍然呈现较明显的聚集分布。阴坡和阳坡的空间依赖范围分别为9.45 m和13.48 m,低龄幼虫在阳坡植株间的相互依赖性比阴坡小。对比密林中低龄幼虫的空间依赖范围,疏林中阴阳两坡的空间依赖范围均小于密林。阴坡和阳坡密林中的偏基台值占比分别为61.3%和59.7%,说明总空间变异中有61.3%和59.7%产生于空间自相关现象。
2.2.3 栎黄枯叶蛾低龄幼虫的空间分布分析

图5 栎黄枯叶蛾低龄幼虫的等值线与矢量叠加图Fig.5 Overlay map of isoline and vector for Trabala vishnou gigantina early stage larvae
由栎黄枯叶蛾低龄幼虫的等值线与矢量叠加图可以看出(图5),无论在阴坡还是阳坡,密林还是梳林,栎黄枯叶蛾低龄幼虫均呈现出明显的聚集分布特点:样地内有多个聚集斑块,每个聚集斑块的幼虫数量从中心区域向四周逐渐减小,这表明了低龄幼虫的聚集强度、空间尺度和扩散方向。聚集斑块在样地边缘的分布略多于样地内部,在林缘主要呈聚集条带状分布,在林间的低龄幼虫数量较少,呈现出明显的边缘效应。对比低龄幼虫在阴阳两坡的空间分布特征,未呈现出明显的方向性。
3 结论与讨论
深入掌握害虫主要为害阶段的数量动态变化规律,对于揭示害虫的危害特性与发生发展规律,采取有效措施控制其发生危害具有非常重要的意义。本研究发现,栎黄枯叶蛾幼虫在6月上中旬数量急剧增加,但由于其虫龄较低,危害较轻,而且幼虫抵抗力也较弱,因此可在此时及时采取措施杀灭幼虫,从而降低虫口数量,减轻危害。相关研究表明,不良的气候条件,如大风、暴雨等剧烈的天气变化,都会对害虫幼虫的种群密度[3,15]产生影响。由于调查时间和试验条件所限,本研究没有对气候因子如何影响栎黄枯叶蛾幼虫种群数量动态进行探讨,相关的研究还有待未来进一步开展。
地统计学的应用必须建立在相关生态学基本理论之上,并结合昆虫的生物学特性进行研究,其分析结果应符合昆虫的生物学特性[16]。本试验通过地统计学分析,表明不同坡向与疏密度的沙棘样地中栎黄枯叶蛾低龄幼虫均呈现明显的聚集分布。野外调查发现,栎黄枯叶蛾卵多产于茧壳表面、枝条及叶片等处,卵块一般呈长条状,卵粒双行相间排列,同一卵块基本在同一天整齐孵化,低龄幼虫有群集性,集中在产卵地附近取食,3龄后幼虫开始四散危害[11],因此在沙棘林中会产生一个为害中心,逐渐向四周扩散,因此成虫的产卵习性是形成其幼虫的空间分布格局的一个主要原因。低龄幼虫在疏林中不同坡向的空间依赖范围均小于密林,说明疏林中低龄幼虫的空间聚集度高于密林,这一现象可能是由于疏林中沙棘的枝叶间相互距离相比密林较远,低龄幼虫在疏林中不易扩散。低龄幼虫在密林与疏林阴阳两坡的空间分布特征未呈现出明显的方向性,说明坡向不同导致光照强度的方向性对低龄幼虫空间分布的影响十分有限。
本试验所选样地中沙棘分布比较规则,分析得出的拟合参数中R2值较大,RSS值较小,这与同样在陕西吴起沙棘人工林中对栎黄枯叶蛾蛹和卵[8- 9],以及在辽宁建平沙棘人工林中对沙棘木蠹蛾卵、幼虫和蛹[17-18]的空间分布分析结果较为相似。而孙鹏举等[19]在分布不规则的自然沙柳林中对柳毒蛾的空间分布分析则得出较小的R2与较大的RSS值,这说明样地内植被分布情况会直接影响害虫的空间分布。本试验再次验证了在植被分布较为规则的人工林中,使用地统计学方法分析空间变异得出的变异函数模型拥有较高的拟合度。
本研究结果显示,沙棘林中栎黄枯叶蛾低龄幼虫存在着明显的边缘效应。边缘效应的产生主要是由于林缘和林内的生境条件如温度、湿度、气流、光照度等存在差异。原因可能是林地边缘通风透光性能较强,干燥性较好,比较适宜雌蛾在此产卵。另外,由于林缘空气流动性较好,雌蛾释放的性信息素比林内传播的更远,容易吸引到雄蛾进行交配,雌蛾交配后,由于其飞行能力较差,一般就集中在林缘附近产卵,导致林缘低龄幼虫密度增加。根据栎黄枯叶蛾的生物学特性,卵的空间分布极大地影响了幼虫的空间分布情况,因此,林地边缘和内部不同环境因子如温度、湿度、光照等对产卵的影响都可能影响低龄幼虫的空间分布,相关的研究还需进一步加强。
本研究表明,栎黄枯叶蛾低龄幼虫在不同坡向和疏密度的沙棘林中均呈明显的聚集分布,且多集中于产卵地附近,成虫的产卵习性是形成其幼虫的空间分布格局的一个主要原因。低龄幼虫的分布呈现出明显的边缘效应,但未呈现出明显的方向性。在实际防治中根据本文结果并结合幼虫种群数量动态变化调查,于幼虫发生盛期及时采取措施杀灭幼虫,可以达到良好的防治效果。
