入侵种互花米草空间扩张模式识别与景观变化模拟
2018-09-19刘红玉李玉凤谢富赋
王 娟,刘红玉,李玉凤,刘 伶,谢富赋
1 南京师范大学地理科学学院,虚拟地理环境教育部重点实验室,南京 210023 2 盐城师范学院城市与规划学院,盐城 224051 3 江苏省地理环境演化国家重点实验室培育建设点,南京 210023 4 江苏省地理信息资源开发与利用协同创新中心,南京 210023
互花米草(Spartinaalterniflora)原产于大西洋沿岸,是禾本科米草属多年生草本植物,秆密丛生,耐盐耐淹,具有较强的物种竞争力和保滩促淤能力[1- 2]。1979年互花米草引入中国,1982年在江苏沿海地区分批试种[3- 4]。1985年,江苏沿海试种面积为8.00 km2。经过多年的定居和扩散,到2007年,互花米草在中国跨越近22个纬度,自辽宁到广西的大部分滩涂上均有分布,其中以江苏的面积最大、范围最广,面积已扩展为187.11 km2,占到全国海岸带互花米草总面积的54.31%[5- 6]。盐城海岸是江苏海岸主要组成部分,属于淤泥质海岸类型,适宜的气候、水文条件以及合适的生态位为互花米草生存与空间扩张创造了十分有利的环境与条件。1987年,盐城滨海湿地互花米草面积仅为0.49 km2,2007年扩散到123.17 km2,2012年的面积为158.30 km2;斑块平均面积由1987年的0.13 km2增加到2007年的0.75 km2[7]。
盐城保护区核心区是互花米草分布最为集中和扩张最为典型区域,互花米草的入侵改变了原有区域生态系统的组成与景观结构,一直受到许多学者的广泛关注[8- 9]。基于景观格局角度的相关研究表明,自20世纪90年代,互花米草以侵占光滩向海扩张为主,其质心一直向东南偏移。1987年盐城保护区核心区互花米草面积为0.44 km2,至2006年,景观格局上已由斑块状分布发展为连续、条带状分布,形成面积33.81 km2、平均宽度为1780 m、最大宽度为2900 m的互花米草景观带;斑块平均面积由1987年的0.05 km2增加到2006年的1.21 km2。互花米草与本土碱蓬(Suaedasalsa)竞争后成为滨海湿地的先锋植被[4, 10- 12]。互花米草滩占据潮间带的中下部,导致碱蓬沼泽与光滩完全分隔。1996—2006年互花米草侵占碱蓬沼泽的平均速度为1.03 km2/a,侵占光滩的平均速度为1.27 km2/a;2006—2010年互花米草沼泽侵占光滩的面积大于其入侵碱蓬沼泽的面积;但2010—2011年互花米草侵占光滩面积仅有0.44 km2,而侵占碱蓬面积达1.80 km2[13]。同时,互花米草的入侵对区域生态过程和生态功能产生了重要影响。基于功能的角度,互花米草被列为湿地的主要威胁之一,对潮汐水盐和营养物质输送过程产生重要影响[14-15],从而改变了大型底栖动物群落的结构和多样性[16],对丹顶鹤越冬食物来源带来不利[17-18]。另外,相关学者从种群繁殖方式研究了扩张能力,互花米草无性繁殖方式的定居成功率、生长、繁殖等指标均优于有性繁殖方式,互花米草的竞争力大于本土碱蓬的竞争力[19- 20]。互花米草向陆入侵本土碱蓬植被群落的趋势突出,在生态交错带与碱蓬植被产生竞争[21- 22]。
由此可见,当前研究中包括了互花米草入侵后景观结构的改变、生态功能的退化、种群繁殖方式等方面,但利用景观指数识别互花米草向陆入侵碱蓬的基本模式鲜有报道。本研究利用高分遥感影像,结合野外详细调查研究,从景观生态学角度,深入探讨互花米草向陆扩张特征与规律,首次提出入侵的3种基本模式,并预测未来5年互花米草侵占碱蓬沼泽的状态,以期为进一步认识互花米草入侵机制打下基础,也为区域湿地生态系统的功能维持提供科学依据。
1 研究区概况
江苏盐城国家级自然保护区,地处江苏中部沿海,地跨响水、滨海、射阳、大丰、东台五县(区、市),主要保护丹顶鹤(Grusjaponensis)、黑嘴鸥(Larussaunders)等珍稀野生动物及其赖以生存的滩涂湿地生态系统[23-24]。本研究区域为江苏盐城国家级珍禽自然保护区的核心区,其范围北至射阳县的新洋港出海河南岸,南至大丰区的斗龙港出海河北岸,西至海堤,东至光滩边缘。该区受潮汐作用显著,是典型的淤泥质潮滩湿地,其上发育着复杂的潮沟系统。区域内湿地受人类干扰较弱,植被类型丰富,保留着较完整的潮滩植被演替序列,由陆地向海洋主要分布有芦苇(Phragmitescommunis)沼泽、碱蓬沼泽、互花米草沼泽和光滩。研究区的位置范围和区内景观类型如图1所示。
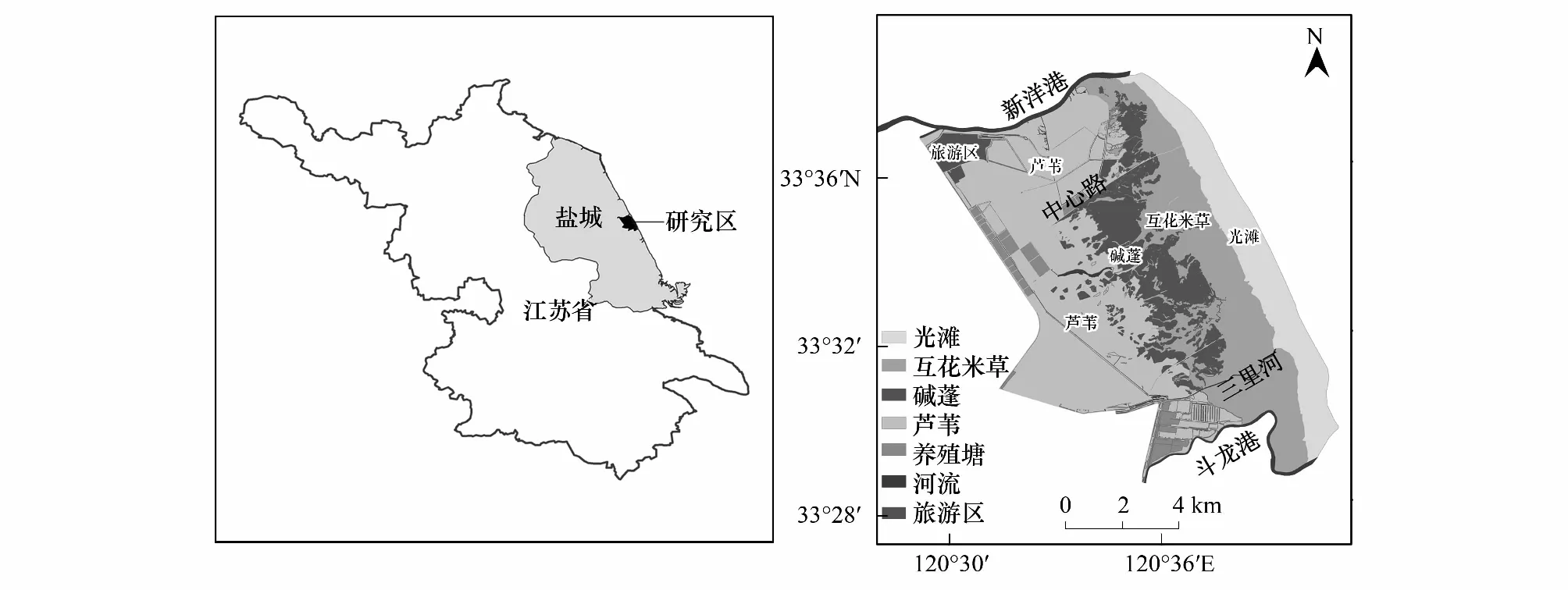
图1 研究区位置和范围Fig.1 Location and scope of the study area
2 数据来源与研究方法
2.1 数据来源与处理
遥感技术广泛应用于生态系统中入侵物种的研究[25-29]。为了揭示区域互花米草空间扩张基本模式,需要从时间和空间两方面,把握互花米草扩张的特征与规律。实地调查发现互花米草向陆入侵扩张斑块具有小而分散特征,必须选取高分辨率遥感影像进行细节研究。为此,研究选取了两期ETM+影像和一期高分影像。
选取的ETM+遥感影像分别是2006年5月21日和2011年5月9日处于植被生长期的两期影像,该影像的7个多光谱波段和1个全色波段进行融合后的分辨率为15 m,用于识别互花米草的双向扩张特征与趋势。本研究选取的是高分二号(GF- 2)在2015年4月10日的遥感影像,用于识别互花米草侵占碱蓬的模式。该影像包含了多光谱的4个波段(分辨率3.2 m)与1个全色波段(分辨率0.8 m),两种分辨率数据可以实现高精度融合,光谱特征明显,图像纹理清晰,层次分明,可以满足精细尺度研究需要。ETM+遥感影像,经ENVI 5.1去除影像条带、大气校正、几何校正、决策树分类等步骤,结合人工判读,通过Confusion matrix进行精度检验,总体精度可达95.6%以上。高分二号遥感影像数据获取时会有几何和辐射变形,需要进行预处理,再在ENVI 5.1中选择面向对象的分类方法解译并解决“椒盐现象”,然后对解译精度进行检验,总体精度在98%以上,能够达到生态系统和植物群落尺度,足够反映互花米草群落在碱蓬滩的小斑块分布与扩张信息,满足研究需要。最终在ARCGIS 10.2中生成2006年、2011年和2015年3期景观类型图,采用统一的坐标系(WGS1984)、统一的地图投影(UTM)和统一的景观分类系统。
2.2 研究方法
首先利用质心方法,揭示区域互花米草扩张方向和速度,再结合景观指数分析互花米草景观变化特征和空间扩张基本模式。
2.2.1 质心的变化
通过空间质心偏移变化,能够揭示互花米草在空间上的扩张方向和速度[30- 31]。公式为:
式中,Xc和Yc是按面积加权的景观类型质心坐标,Xi和Yi是某一景观类型的第i个斑块的质心坐标,Ci为某一景观类型的第i个斑块的面积,n是某一景观类型的斑块总数目。
2.2.2 景观指数分析
景观指数可以高度浓缩景观格局信息,反映其格局变化简单定量指标[32]。针对区域景观变化特征,选择以下景观格局指数揭示互花米草景观格局特征及其变化,包括斑块密度(PD)、聚合度指数(AI)、面积加权平均形状指数(AWMSI)、斑块分维数(PFD)和破碎度指数(SPLIT)来反映景观格局变化,如表1所示。景观指数的计算在Fragstats 3.4中实现。

表1 景观指数描述
2.2.3 扩张模式识别
利用景观扩张指数(Landscape Expansion Index,LEI),定量描述互花米草扩张规模,并识别扩张模式[33]。其表达式为:
从上式可以看出,LEI是对斑块水平的扩张过程进行定量表达。式中,LEI为斑块水平的景观扩张指数,Ap为斑块的扩张面积(Ap>0),A0为斑块的原面积。若A0≠0,LEI的取值范围为(-1,1),其扩张模式为以原生斑块为基础的边缘扩张,并且LEI的值越大,说明景观扩张规模越大。当扩张面积小于原有斑块面积时,LEI<0;扩张面积等于原有斑块面积时,LEI=0;扩张面积大于原有斑块面积时,LEI>0。若A0=0时,假设原有斑块为一个点,此时LEI的值为1,其扩张模式为非边缘式扩张。非边缘式扩张包括外部隔离扩张和潮沟引领式扩张两种模式,两种模式的斑块复杂性相差较大,可以根据斑块分维数PFD来确定是外部隔离扩张还是潮沟引领式扩张[34]。
3 结果及分析
3.1 互花米草空间扩张特征与景观格局变化
3.1.1 互花米草空间扩张特征

图2 不同年份互花米草斑块质心变化图 Fig.2 The patch centroid variation of Spartina alterniflora in different years
就斑块质心整体移动方向而言,两个时段均以向西北方向偏移为主;从移动速度来看,2006—2011年以向北偏移速度较快,2011—2015年以向西偏移速度较快,如图2所示。
结合图1和图2可以看出,3期数据中互花米草的质心均位于中心路以南。从偏移速度来看,在2006—2011年,斑块质心向西偏移约93.0 m,平均每年向碱蓬方向偏移18.6 m;向北偏移约477.6 m。在2011—2015年,斑块质心向西偏移约111.6 m,平均每年向碱蓬方向偏移27.9 m;向北偏移约33.3 m。上述数据表明互花米草沼泽向海方向演替的速度在降低,而向陆地演替的速度有所增加,植被质心向陆方向移动。互花米草向陆方向侵占碱蓬和向海方向侵占光滩的面积如图3所示。

图3 2006—2015年互花米草沼泽向陆和向海扩张状况Fig.3 Landward and seaward expansion of Spartina alterniflora marsh in 2006—2015
2006、2011和2015年互花米草沼泽的面积分别为33.81、41.50 km2和45.95 km2,年均扩张1.35 km2。从图3中可以看出,2006—2011年和2011—2015年期间,互花米草均向陆侵占碱蓬沼泽,同时向海侵占光滩湿地,表现为向海与向陆的双向扩张。2011年互花米草沼泽的平均宽度比2006年增加了427.3 m,平均向陆地方向推进了约193.71 m,平均向海洋方向扩张了约233.59 m,以向海扩张为主;2015年互花米草沼泽的平均宽度比2011年增加了247.27 m,平均向陆地方向推进了约229.58 m,平均向海洋方向扩张了约17.69 m,表明这一时段互花米草扩张转向向陆为主。
从向陆方向来看,2006—2011年互花米草的最大扩张宽度达到1327.42 m,扩张的总面积为5.10 km2;2011—2015年最大扩张宽度达到1879.7 m,扩张的总面积为7.25 km2。与前一时段相比,向陆扩张宽度增大,向陆扩张面积增大。相对于2011年,2015年互花米草面积增加了4.43 km2,而向陆扩张面积为7.25 km2,表明在一些海岸段发生了侵蚀现象。受互花米草扩张影响,碱蓬沼泽无论在向海一侧还是在向陆一侧都呈现出减少的趋势。
3.1.2 互花米草景观格局变化
近10年研究区内互花米草沼泽各景观指数变化如表2所示。

表2 2006—2015年互花米草景观格局指数变化
表2反映出近10年中互花米草沼泽各景观指数变化。其中,2015年数据基础为高分辨率影像图,能清晰地定位到每个小斑块,考虑到ETM+影像的分辨率为15 m,所以对2015年的互花米草景观进行像元过滤,使其与前两期数据具有可比性,可以更加真实地反映出互花米草沼泽的景观格局。景观斑块密度PD呈现增大趋势,2015年PD值为2.81,说明互花米草斑块数量不断增多,在一定程度上反映了区域内的景观破碎化趋势。从2006年到2015年,AWMSI的值逐渐增大到5.35,说明互花米草条带形状变得越来越复杂。从聚合度指数看,AI值在2006—2015年总体上稳步上升,说明近10年来互花米草的聚集程度越来越高。
3.2 互花米草扩张基本模式及其变化
3.2.1 互花米草扩张基本模式识别
利用野外调查和景观扩张指数(LEI),对互花米草扩张模式进行识别。研究发现,从扩张形式上看,互花米草的扩张分为边缘扩张、外部隔离扩张和潮沟引领式扩张3种模式,如图4所示。
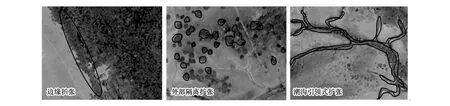
图4 2015年GF- 2影像互花米草扩张模式示意图Fig.4 Expansion patterns of Spartina alterniflora on the GF- 2 image in 2015
第一类扩张模式:边缘扩张。对于核心区,互花米草沼泽包括三大原生斑块,第一块位于中心路以北,第二块位于中心路和三里河之间,第三块位于三里河和斗龙港之间。这种扩张方式的特征是相对于前一期数据来看,从原生斑块的边缘区域向外扩张,边缘扩张斑块和原生斑块相邻接。
第二类扩张模式:外部隔离扩张,非边缘扩张方式之一。其基本特征是相对于前一期数据,新生互花米草斑块和其他所有的互花米草斑块不相邻,近似于圆形,呈现离散分布状态。
第三类扩张模式:潮沟引领式扩张,非边缘扩张的另一种方式。潮沟引领式扩张的基本特征是互花米草沿着潮沟发育的路线向陆碱蓬区突起,呈现树枝状扩张,且潮沟发育越完善,互花米草突起越显著。
3.2.2 3种扩张模式互花米草空间变化特征
互花米草边缘扩张模式主要表现为以原生斑块为基础,向海入侵光滩,向陆入侵碱蓬沼泽,抑制碱蓬生长,加速了碱蓬景观的质心西移。外部隔离扩张特征表现为互花米草的种子落到碱蓬滩内,成功定居后逐渐长成近似圆状,每个斑块之间均不相邻接并且以“摊大饼”方式不断向外扩张,连接成带状斑块后继续扩张。可见,外部隔离扩张模式互花米草扩张表现为景观类型的斑块密度大,破碎化程度高。潮沟引领式扩张模式表现为互花米草在边缘扩张的基础上沿着潮沟发育的方向在潮沟两侧不断向外扩张。潮沟引领式扩张模式的斑块分维数PFD均大于外部隔离扩张模式的斑块分维数,表明潮沟引领式扩张模式比外部隔离扩张模式的互花米草斑块形状更复杂。LEI区间分布及部分景观指数如表3所示。

表3 2011—2015年互花米草LEI区间分布及景观指数
由表3中可知,利用LEI区间分布可以识别出边缘扩张和非边缘扩张。LEI≠1时为边缘扩张,LEI=1时为非边缘扩张,再用PFD进一步区分非边缘扩张模式的两种模式,若LEI=1且PFD<1.10,则为外部隔离扩张,若LEI=1且PFD>1.15,则为潮沟引领式扩张。在边缘扩张模式中,包括LEI∈(-1,-0.9)的80个斑块和LEI∈(-0.9,-0.6)的4个斑块。LEI的值越小,说明扩张面积越小于原生斑块面积,是一种小规模的边缘扩张但不容忽视。
从2011年到2015年,相对于3大原生斑块,互花米草扩张的斑块总数为7368个。从数量看,互花米草的主要扩张模式为大量的外部隔离扩张,斑块数为7254,占总扩张斑块数的98.45%;仅有84个斑块为边缘式扩张,所占比例为1.14%;潮沟引领式扩张的斑块数量为30个,所占比例不足1%,但这种扩张方式对潮沟系统发育起到重要的作用。从面积看,边缘扩张斑块的面积为4.71 km2,外部隔离扩张斑块的面积为0.84 km2,潮沟引领式扩张斑块的面积为2.80 km2,三类扩张的总面积为8.35 km2,占2015年互花米草总面积的18.18%。从景观指数看,外部隔离扩张的AWMSI值为1.00,说明此类扩张斑块的形状均接近于圆形,而边缘扩张和潮沟引领式扩张斑块的形状较为复杂;外部隔离扩张的破碎度指数(SPLIT)为1479.89,显著大于另外两类扩张模式,说明外部隔离扩张的小斑块数量多,碱蓬沼泽的破碎化态势严重。
3.3 未来年份互花米草向陆扩张预测
通过在外部扩张斑块处设置22个样点,在沿潮沟方向扩张互花米草边缘处设置8个系列样带(共32个样点),在原生斑块向陆一侧设置16个样点,每年在每个样点附近取5个距离,确定不同扩张方式的年扩张速度。2014—2016年以来,向陆一侧边缘扩张的平均速度约为1.06 m/a;外部隔离扩张斑块往外生长的平均速度约为0.78 m/a;潮沟引领式扩张沿潮沟向两侧生长的平均速度约为0.94 m/a。以2015年影像作为基础数据,用上面3个扩张速度以生成缓冲区的方式模拟出2016年、2018年和2020年的互花米草沼泽向陆扩张态势,效果图如图5所示;互花米草的面积、斑块数量和面积加权平均形状指数等结果数据如表4所示。
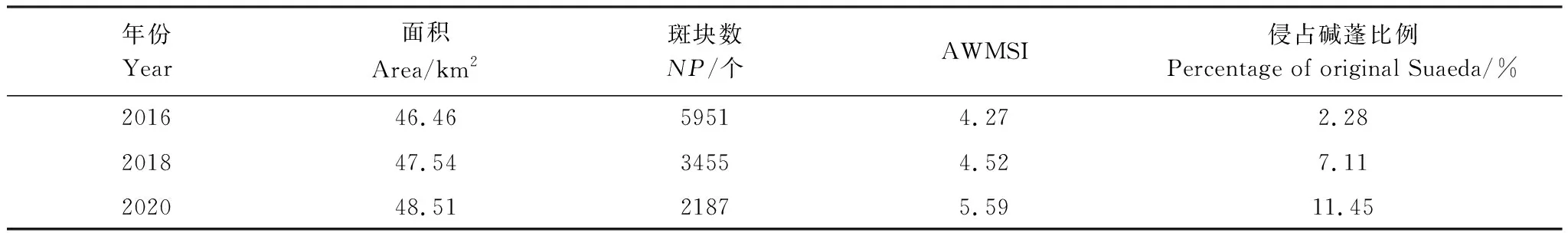
表4 预测互花米草沼泽景观特征
表4和图5显示,在未来5年,互花米草的扩张斑块数由2015年的7368个逐渐减少到2187个,斑块密度将随之变小,说明很多外部隔离扩张模式的斑块由离散个体逐渐连接成片呈现连续分布;随着年份推移,AWMSI的值逐渐增大,说明互花米草景观形状复杂度提高;到2020年,互花米草侵占碱蓬的比例达到11.45%,研究区内的景观结构必将发生较大的变化。

图5 互花米草沼泽向陆扩张模拟Fig.5 The landward expansion simulation of Spartina alterniflora marsh
4 讨论
互花米草入侵对潮间带盐沼景观演变的影响是研究关注点之一[35-36]。通过对互花米草沼泽的扩张特征分析发现,近期内其质心持续向西北方向偏移,这与张华兵、王聪等的研究结果一致[15,37]。2006—2011年间,互花米草向海侵占光滩的速度略大于向陆侵占碱蓬的速度,而2011—2015年间,平均向陆推进的距离远大于向海的距离,互花米草向海扩张趋势显著减弱,向陆扩张趋势更加明显。左平等[7]的研究也表明到2012年,核心区部分岸段已受到海岸侵蚀影响,互花米草前缘出现陡坎现象,这可能与核心区北部的二类口岸射阳港和核心区南部的一类口岸大丰港的建设相关。互花米草只有通过滩面的不断淤高才能继续向海扩展[38],而港口的建成改变了潮流,造成核心区某些岸段的侵蚀。野外调查同样发现,在中心路附近的光滩上,原来成片的互花米草景观现在只剩下了岛屿状分布的互花米草斑块。
互花米草秸秆粗密、地下根茎发达,能够促进悬浮泥沙沉淀,导致沉积物积累,在潮滩湿地生境中具有超强的繁殖力[3,39]。受到水盐条件、微地形和潮汐等多种因素的影响[14],2006—2015年,互花米草的面积加权形状指数增大了近一倍(2015年AWMSI=8.63),聚集程度不断增高,众多新生斑块与碱蓬沼泽交错分布。与之相对应的,本土碱蓬沼泽面积从2006年的31.25 km2收缩到2015年的22.36 km2,斑块数剧增,呈现出面积减少、破碎化程度加剧的趋势。这极不利于依赖于碱蓬生境的丹顶鹤、黑嘴鸥等珍稀鸟类的种群维持,也影响了潮滩湿地及其物种多样性保护[21- 22]。互花米草的扩张与潮沟的发育相辅相成,互花米草沼泽的景观形状会更加复杂。潮沟的侧向侵蚀较难,涨潮时,潮水只能沿着潮沟不断西进碱蓬滩[40],调查发现在碱蓬滩西侧的碱蓬芦苇交错带中已有零星互花米草斑块分布。互花米草强大的滞流作用使潮水滞留在互花米草-碱蓬交错带滩面,加快了互花米草的生长和繁殖,互花米草向陆扩张侵占碱蓬的趋势会更加明显[41]。
利用高分辨率影像更适合及时发现和识别互花米草入侵本土碱蓬的模式[42]。本文对互花米草向陆的规律进行分析并结合野外调查,首次利用LEI识别出了3种侵占碱蓬的模式。连续的野外监测发现边缘扩张的速度和面积要大于外部隔离扩张和潮沟引领式扩张模式。边缘扩张主要依靠地下根茎进行分蘖繁殖,而互花米草高度发达的通气组织可为其根部提供足够的氧气有利于邻近互花米草植株的生长[43]。外部隔离扩张模式中,互花米草的有性繁殖对开拓新生境有着非常重要的意义,即先借助种子在碱蓬滩占据新生境,再通过分蘖繁殖以“摊大饼”方式扩张成入侵斑块。外部隔离扩张的斑块数量占扩张斑块总数的98.45%,碱蓬沼泽的破碎化态势严重,其中60.26%的斑块面积小于100 m2,这在一定程度上也说明了近几年互花米草以向陆扩张为主的现状。潮沟引领式扩张是互花米草迅速入侵碱蓬沼泽不可忽视的扩张模式,潮沟的发育对互花米草入侵碱蓬有引领式作用[40]。在潮汐的作用下,种子与部分根状茎一并随潮水漂流,主要依靠种子在潮沟边缘萌发、定居,再依赖于分蘖繁殖不断向潮沟两侧扩张侵占碱蓬生境。同时,互花米草的不断扩张也会影响潮滩滩面和潮沟的发育[40]。互花米草的入侵使得潮滩滩面不断淤高,从而改变了潮汐动力,潮沟的发育呈现窄而深的趋势并且两侧边缘往往有高低不等的不对称的沿岸堤。其3种模式的扩张均以分蘖繁殖为主,这类似于在已有斑块外部生成缓冲区,但潮汐作用和风力作用会使互花米草种子在碱蓬沼泽内定居,生成新的斑块,所以本研究仅依据扩张速度模拟了未来5年内互花米草向陆侵占碱蓬的状态。可以预见,互花米草沼泽的向陆扩张速度会快于模拟效果,5年后碱蓬沼泽减少的面积会超过2.56 km2,潮沟系统也会更加发育。今后的研究可以在两期及以上高分影像的基础上,考虑潮汐作用和水盐过程,揭示互花米草入侵碱蓬滩的机理并结合前期影像进行模拟预测扩张趋势。
5 结论
盐城淤泥质潮滩湿地互花米草的入侵改变了原有的湿地景观,其质心逐渐向西北偏移,高成功入侵率使其向陆扩张速度和面积均大于向海扩张速度和面积,近期入侵以向陆方向移动为主。2006—2015年互花米草面积呈显著增加趋势,年均扩张1.35 km2,并与碱蓬群落呈大片交错分布,景观形状日趋复杂。互花米草群落扩张模式分为边缘扩张、外部隔离扩张和潮沟引领式扩张3种模式,潮沟发育是互花米草快速向陆入侵碱蓬的重要动力。
