不同寄主植物对梨小食心虫幼虫龄数的影响
2018-09-12孔维娜郭永福胡荣山柴晓晗马瑞燕
王 怡 ,孔维娜 ,郭永福 ,胡荣山 ,柴晓晗 ,李 捷 ,马瑞燕
(1.山西农业大学农学院,山西 太谷 030801;2.山西省农业科学院植物保护研究所,农业有害生物综合治理山西省重点实验室,山西 太原 030031;3.蒲县农业资源开发中心,山西 临汾 041200;4.山西省农业科学院果树研究所,山西 太谷 030800)
梨小食心虫(Grapholita molesta Busck)属鳞翅目(Lepidoptera)卷蛾科(Tortricidae),是一种世界性的蛀果类害虫[1-2]。作为寡食性昆虫,它主要为害蔷薇科植物,包括苹果(Malus pumila),桃(Amygdalus persica),油桃(Prunus persica var.nectarina),梨(Pyrus spp),杏(Armeniaca vulgaris),温柏(Pyrus cydonia),扁桃仁(Amygdalus communis),李子(Prunus salicina)和樱桃(Cerasus pseudocerasus)等[3-5],具有明显的转移寄主为害特性[6-8]。在年生活史早期,该虫最早为害桃树嫩梢,果实出现以后逐渐转移到果实上进行为害。在年生活史后期,该虫会转移到邻近的梨园或苹果园进行为害[9]。可见,寄主植物是影响梨小食心虫生长发育的重要生态因子之一。
大量试验结果表明,不同的寄主植物显著影响梨小食心虫生长、发育、存活率以及繁殖率,从而形成不同的寄主种群[10-13]。其中,幼虫在不同寄主植物上的生长发育速度存在差异。例如,在桃上的生长比苹果快[14]。幼虫脱果时期及盛期在不同种类的寄主植物上存在差异,山楂(Crataegus pinnatifida)饲喂的幼虫脱果各时期均比苹果饲喂的梨小食心虫提前4~5 d,但盛期均为6 d左右[15]。幼虫的发育速率主要由2个因素决定,即温度和食物[16]。温度较高或食物营养物质较为丰富时,会造成较快的发育速率[16]。RUSSEL等[17]研究认为,当室温为11~28℃时,用人工饲料饲养的梨小食心虫幼虫形成5个龄数;用苹果和桃饲养的幼虫在25℃时形成5个龄数;在田间环境下苹果上的幼虫同样形成5个龄数;另外,在新西兰和法国,桃梢上的幼虫只发育成4个龄数;在田间桃果上的梨小食心虫幼虫同时存在4个和5个龄数。PETERSON等[18]研究认为,田间苹果上的幼虫在6月存在4个龄数,在5,7,9,10月存在5个龄数,桃上的幼虫在6月和9月上旬存在4个龄数,在7月和9月下旬存在5个龄数。可见,寄主植物的差异与幼虫龄数的划分是密切相关的。
确定幼虫龄数的方法多种多样。通过测量幼虫头部口上片最大宽度、尾部钳状突最大宽度、钳状突最大长度、钳状突左侧侧缘最大长度等形态特征,对柑桔爆皮虫(Agrilus citri)幼虫的龄数进行了划分[19]。对于鳞翅目而言,由于头壳骨化强而形态较为稳定,头部宽度为较好的测定指标。DYAR等[20-21]在1890,1894年均通过测量计算鳞翅目幼虫的头壳宽度发现了与龄数分布有关的Dyar定律,认为相邻龄数的幼虫其骨化部位以一定的几何级数方式增长[22-23]。梨小食心虫幼虫龄数有的是根据上颚宽、幼虫的虫体颜色进行划定[24-26],但大多数是通过头壳宽度划分的[17,27]。但是,前期的研究结果在室内试验中未考虑寄主植物种类和成熟度,在田间试验中未排除温度对试验结果的干扰。寄主植物上幼虫的龄数测定是害虫预测预报以及制订有效防治策略的重要依据。目前寄主植物的种类和物候期对幼虫龄数的影响还不明确。
本研究在实验室恒温条件下,选取不同种类的寄主植物,用不同成熟度的寄主果实饲喂梨小食心虫幼虫,通过测定幼虫的头壳宽度,以明确寄主种类和成熟度这2个寄主因子对梨小食心虫龄数分布的影响,进而为寄主与梨小食心虫的生态适应性提供理论依据。
1 材料和方法
1.1 试验材料
1.1.1 供试寄主植物 试验所需不同种类的寄主植物均采自山西省农业科学院果树研究所苹果产业国家技术系统晋中太谷试验站、梨产业国家技术系统晋中太谷试验站以及桃种质资源圃。该区位于山西省晋中市太谷县,37°23′N,112°32′E,海拔 830m。桃树为6年生的大久保毛桃。苹果树为8年生的丹霞苹果。梨树为7年生的库尔勒香梨。果树每年进行常规修剪,定期喷施杀菌剂预防病害。它们的果实成熟期集中在7—10月。
1.1.2 供试虫源 梨小食心虫虫源来自于山西农业大学农学院生物安全与生物防治实验室继代培养30代的梨小食心虫种群,全部利用人工饲料进行饲养,以保证虫源状态的一致性。试验所用虫态为卵。选取同一批羽化的梨小食心虫成虫,按1∶1的比例放置于交配罐中,待其交配产卵后,每日更换卵纸并选取同一天所产卵用于试验。
1.2 试验方法
不同成熟状态的果实采集时间依据同一年对梨小食心虫发生动态监测的平行试验,分别在不同世代的梨小食心虫幼虫为害高峰期进行寄主材料的采集,保证果实的成熟状态与发生时间呈对应关系(表 1,2)。

表1 寄主植物物候期[28-30]

表2 梨小食心虫幼虫2015年发生世代时间(山西太谷县)
每个果实上放置5粒黑头卵,单独放置于带盖的塑料小碗(直径为10 cm)内饲养。在一个世代内,一个种类共需150个果实和750粒卵。每隔3 d,随机抽取每个种类中的120个果实进行解剖。试验在体视显微镜(MOTIC SMZ-140 SERIEC)下用游标卡尺(量程0~150 mm)对幼虫的头壳宽度(与虫体纵轴垂直的头壳最宽处Y)进行测量(图1)。

1.3 数据分析
使用SPSS Statistics 19.0统计分析软件进行频次分布分析,制作频次分布图,初步确定幼虫的龄数。然后计算每个龄数对应的头壳宽度均值。应用Brooks指数和Crosby指数验证分龄的合理性。Crosby生长法则认为,Crosby指数大于10%表明分龄指标的分组不合理[31]。同时将各龄幼虫头壳宽度与相应龄数进行曲线拟合,获得二者之间的函数关系,计算回归方程以进一步验证分龄的合理性。

其中,Xn和Xn-1分别表示n龄和n-1龄幼虫各个测量指标的平均值;bn和bn-1分别表示第n,n-1个Brooks指数。
通过研究发现,梨小食心虫幼虫头壳宽度(y)与幼虫龄数(x)的函数关系为开口向上的二次函数y=ax2+bx+c,具体定义域为 1≤x≤5(4)的上升区间,且x(虫龄)为整数。进一步对函数方程进行求导获得导数方程 y′=2ax′+b,其中,y′表示各龄头壳宽度的增长速率,带入龄数(x):1≤x′≤5(4),即可计算函数各龄数分布点的理论增长速率,导数的斜率2 a即为该生长环境下的增长速率的变化率。
2 结果与分析
2.1 幼虫头壳宽度频次分布与龄数的初步划分

根据测得的头壳宽度,将取食不同寄主植物的梨小食心虫在不同世代的幼虫头壳宽度绘制频次分布直方图。在第1代幼虫发生期,桃和苹果饲养的幼虫均具有5个明显的分布区和聚集区。在第2代幼虫发生期,取食桃的幼虫具有4个明显的聚集区,而取食苹果的幼虫具有5个或者4个明显的聚集区。在第3代和越冬代幼虫发生期,取食桃和苹果的幼虫均具有5个明显的聚集区,而取食梨的幼虫具有4个明显的聚集区(图2~4)。
根据Dyar定律,鳞翅目幼虫各龄间的头壳宽度是按一定的几何级数增长的,结合频次分布直方图的聚集区域和Dyar定律,对相邻龄数头壳宽度的几何级数进行划分,并对不同世代取食不同寄主植物的幼虫龄数进行初步划分。结果表明,取食桃的第1代、第3代和越冬代幼虫龄数可初步划分为5龄,取食桃的第2代幼虫龄数可初步划分为4龄。取食苹果的第2代幼虫可初步划分为4龄或者5龄,其余3代幼虫龄数可初步划分为5龄。取食梨的第3代和越冬代幼虫龄数可初步划分为4龄(表 3)。



表3 梨小食心虫幼虫在不同寄主植物上龄数
2.2 验证幼虫龄数划分的合理性
根据初步划分的幼虫龄数,对不同寄主植物饲喂不同世代的幼虫所获得的头壳宽度均值进行计算。其中,苹果上幼虫的头壳宽度整体较大,梨上幼虫的头壳宽度较小。桃和苹果上第1代和越冬代幼虫的头壳宽度大于其余2代。
初步划分的幼虫龄数是否合理需要通过Crosby指数和线性回归来进行验证。结果显示,Crosby指数均小于10%,表明在取食不同寄主植物的不同世代幼虫的龄数划分是合理的(表4)。
将不同寄主植物饲养所得幼虫的各龄数头壳宽度与幼虫龄数进行回归分析,再次验证幼虫分龄的合理性。不同世代下,3个种类的寄主植物饲养所得幼虫头壳宽度与其龄数均呈现显著的二次函数关系,R2值均大于0.9。进一步证明幼虫的龄数划分是合理的(表5)。

表4 室内不同寄主植物饲养梨小食心虫幼虫所得头壳宽度及其统计参数
2.3 头壳宽度增长速率与龄数的关系
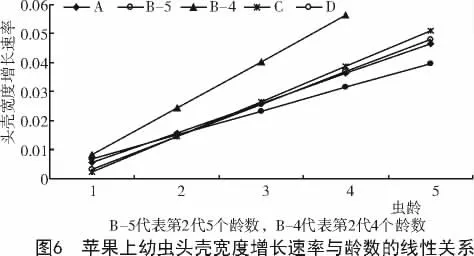
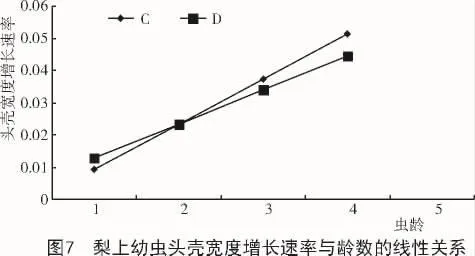
对头壳宽度的增长速率进行计算发现,第4龄幼虫的头壳宽度增长速率与幼虫的龄数分布密切相关,增长速率较大的寄主植物包括:梨小食心虫第2代发生期的桃,第3代和越冬代发生期的梨,以及第2代的苹果(4个龄数)。幼虫在这些寄主植物上的龄数均为4龄。在其余寄主植物上饲养的幼虫,头壳宽度增长速率较小,最终发育为5个龄数(表6)。

表5 幼虫头壳宽度与幼虫龄数的回归关系

表6 幼虫头壳宽度增长速率与龄数的线性关系
此外,第2代发生期的苹果(4个龄数)可使幼虫的头壳宽度增长最快,线性函数的斜率达到0.016,其次为第2代发生期的桃和第3代发生期的梨,线性函数的斜率分别为0.015和0.014。越冬代发生期的苹果使幼虫的头壳宽度增长最慢,线性函数的斜率为0.008 2(图5~7)。

3 讨论
本试验选择幼虫的头壳宽度为测量指标,在不同世代对不同寄主植物上的梨小食心虫幼虫龄数进行划定,并且对其划定的龄数进行验证。结果发现,桃上的幼虫在第1代(5月中旬)、第3代(8月中旬)和越冬代(9月上旬)存在5个龄数,在第2代(7月中旬)存在4个龄数。苹果上的幼虫4个世代均存在5个龄数,但在第2代同时存在4个和5个龄数。梨上的幼虫在第1代(5月中旬)和第2代(7月中旬)无法存活,在第3代(8月中旬)和越冬代(9月上旬)存在4个龄数。这一结果与RUSSEL等[17]和PETERSON等[18]所得田间幼虫龄数较为相似。但后两者的试验是在田间环境下进行的,无法排除如温度等其他因素的影响。本试验排除了田间温度及其他因素可能产生的影响,结合梨小食心虫世代的发生时间,将测定的时间进行了细致的划分,说明寄主植物对梨小食心虫幼虫龄数的影响除寄主种类外,还明显受寄主成熟度的影响。
本研究表明,同一寄主植物既有能力使幼虫龄数变为5龄,也有能力使幼虫龄数变为4龄,因此,时间带来的成熟度对幼虫龄数的影响高于寄主植物种类的影响。根据不同寄主植物的物候期,可以发现桃的成熟期最早,7月桃果开始成熟,而此时为梨小食心虫幼虫发生第2代,幼虫龄数从第1代的5个龄数转变为第2代的4个龄数,龄数缩短。而梨的成熟期较晚,从8月开始梨果逐渐变为梨小食心虫可取食的状态。幼虫龄数受苹果物候期的影响较小。因此,在对寄主植物与梨小食心虫幼虫龄数之间关系的研究中,寄主植物的状态随时间的变化值得重点考虑。
头壳宽度的增长速率与幼虫龄数密切相关,是造成幼虫龄数不同分布规律的原因。从生物的外部形态上代表了幼虫生长发育速度的快慢[16]。对于不适合在室内饲养或观察较为困难的昆虫,无法准确的记录发育历期,可以通过田间采集的方法来研究头壳宽度与龄数的关系,从而对其生长发育的速度进行比较。较快的生长发育速度是适合度高的表现。因此,结合头壳宽度的增长速率与幼虫的龄数可以帮助判断寄主植物的适合度。本试验中适合度较高的寄主植物为梨小食心虫第2代发生期的桃,第3代和越冬代发生期的梨,以及部分第2代发生期的苹果。苹果整体适合度低于桃和梨。这一结果也与梨小食心虫对不同寄主材料的偏爱性和田间实际为害情况相符合,梨小食心虫对第2代的桃以及第3代和越冬代的梨较为偏爱,田间为害情况也较重,而对苹果的偏爱性始终较低。
不同的寄主植物可以充分影响多寄主类昆虫各阶段的生长发育,从而形成以季节为基础的生态上的优势,这是形成不同的寄主型或寄主表型的前提。寄主植物驱动植食性昆虫形成新的寄主型或寄主表型已在多种蛾类害虫上被证实[10-12],这一理论可以帮助多寄主类害虫针对不同的寄主植物种类制定田间防治策略。对于梨小食心虫幼虫而言,本研究认为,寄主植物时间上的作用高于种类上的作用,因此,在制定田间防治策略时,应注重寄主植物在不同生长发育时间段中的寄主状态,即寄主植物时间上的变化。
