唐古特雪莲花部特征及生殖分配的海拔差异
2018-09-10杨亚军王一峰祁如林杨洋
杨亚军 王一峰 祁如林 杨洋


摘 要: 为探究高山植物生殖分配策略以及分析唐古特雪莲花部特征对海拔梯度的响应机制,该研究利用采样调查法和烘干称重法,对分布在青藏高原东缘不同海拔14个居群的唐古特雪莲花部特征和生殖分配进行了研究。结果表明:(1)繁殖分配随个体大小(地上生物量和株高)的增大呈线性递减趋势。(2)花期植株地上生物量、株高、管状小花数目、繁殖器官及营养器官生物量均随海拔升高呈线性递减趋势,管状小花生物量随海拔升高呈线性递增趋势。(3)花期管状小花数量及大小、繁殖器官生物量与营养器官生物量、雄蕊重量和雌蕊重量以及花粉数目与花丝长度之间均存在权衡现象。由此推论:(1)海拔作为外界因子对唐古特雪莲花部特征具有显著的影响,个体大小对其繁殖分配也存在潜在调控作用。(2)在不同海拔梯度上,唐古特雪莲有效地整合了有限的资源,其适应性特征之一就是通过减小个体大小来削弱营养生长以达到促进生殖生长的目的。
关键词: 繁殖生态学, 唐古特雪莲, 花部特征, 海拔, 权衡, 资源分配
中图分类号: Q948.11
文献标识码: A
文章编号: 1000-3142(2018)02-0159-10
Discrepancy caused by various altitudes in both floral traits and reproductive allocation of Saussurea tangutica
YANG Yajun, WANG Yifeng*, QI Rulin, YANG Yang
( College of Life Sciences, Northwest Normal University, Lanzhou 730070, China )
Abstract: To investigate reproductive allocation strategies of alpine vegetation and analyze the response mechanism of floral characteristics to altitude gradient in Saussurea tangutica, the floral characteristics and reproductive allocation of fourteen populations of S. tangutica from different altitudes on the eastern edge of Qinghai-Tibet Plateau were determined through field investigation and drying-weighing method. The results were as follows: (1) Reproductive allocation presented a linear decline with increasing individual size (above-ground biomass and plant height). (2) Above-ground biomass, plant height, the number of tubulous, and the biomass of reproductive and vegetative organs at the flowering stage all showed a linear decline with rising altitude, while the biomass of tubulous increased with rising altitude. (3) There was a common balance between the number and size of tubulous, the biomass of reproductive and vegetative organs, the weight of stamens and pistils, as well as the number of pollen grains and the length of filaments. It can be concluded as follows: (1) As an external factor, altitude significantly affects the floral characteristics of S. tangutica, and individual size also potentially regulates reproductive allocation. (2) S. tangutica at different altitudes can utilize the limited resource effectively and decrease vegetative growth by reducing its size to promote reproductive growth.
Key words: reproductive ecology, Saussurea tangutica, floral characteristics, altitude, trade-off, resource allocation
花部特征作为植物的有性繁殖器官,在植物交配系统的进化中意义重大,人们对此已经有了研究(李新蓉等,2016;杨勇等,2015;Sun et al, 2014)。随着海拔的升高,影響植物生长繁殖的环境因子(如温度、光照)会不断发生变化,这些环境因子的变化会对植物的花部特征产生一定影响,进而影响到植物的繁殖策略(苏梅等,2009)。就虫媒传粉的植物来说,花特征被认为是雌、雄繁殖成功的关键(Wright & Meagher,2004;Han et al,2011)。Kleunen & Ritland(2010)预测了在自然居群中与交配系统相关的花特征的进化趋势,对有花植物而言,特殊的花的多样性结构已经进化形成。通常这种多样性结构能够影响植物对传粉媒介的吸引构件和传粉的效率来影响虫媒传粉植物的繁殖成功(Pires & Freitas,2008)。
生殖分配是植物将所有可利用资源分配给生殖生长的比例,即总资源供给生殖器官的比例,它是植物产生后代、种群得以延续和发展的关键因素。植物的繁殖策略会受到海拔制约下环境因素的潜在调控(张茜等,2013),海拔作为外界环境因子会对植物的生殖分配产生显著影响(张挺峰等,2014;张茜等,2013)。王一峰等(2012a)发现在高海拔地区植物的花部特征产生了转变,花柱和花丝增长,并且将更多资源投入到了传粉吸引结构花瓣,以此来加强该种在高海拔地区对传粉昆虫的吸引能力,从而保证其能在高寒草甸顺利完成有性繁殖。选择典型植物,在海拔差异较大的区域进行海拔与生殖分配的关系研究,将有助于分析海拔对植物生殖生态学的影响及可能机制(马诣欣等,2017)。
青藏高原是中国最大、世界海拔最高的高原,气候寒冷,昼夜温差大,紫外辐射强,生态因素复杂。因此植物可获取的资源总量较低,从而导致投入到繁殖部分的绝对资源减少(王一峰等,2012b)。青藏高原东缘气候条件复杂,唐古特雪莲作为青藏高原东缘风毛菊属雪莲亚属的常见种之一,它可以适应青藏高原的高寒气候。近些年来,国内外对毛茛科(Ranunculaceae)、龙胆科(Gentianaceae)、虎耳草科(Saxifragaceae)等多种植物(梁艳,2009;王赟等,2010)的资源分配有了较多研究,其中也不乏对菊科风毛菊属植物繁殖分配的研究。但对风毛菊属的雪莲亚属中的植物花部特征及生殖分配随海拔的变化以及内部资源权衡的研究鲜有报道,株高可以指示植物的生长势,然而对风毛菊属植物的繁殖分配的相关研究中几乎未涉及植株高度。因此,把植株高度也作为研究指标,来研究高山植物的生殖分配以及高山植物如何调节其生殖分配去适应特定生存环境具有重要的生态学意义(苏智先和钟章成,1998)。基于青藏高原东缘的海拔梯度差异,选择风毛菊属植物开展研究,具有较好的前期基础,将有助于相关机制的揭示。
本研究以分布在青藏高原东缘的风毛菊属植物唐古特雪莲(Saussurea tangutica)为研究对象,主要探讨以下三个问题:(1)唐古特雪莲各部分生物量在不同海拔高度下的差异;(2)唐古特雪莲个体大小(株高和地上生物量)与繁殖分配的关系;(3)唐古特雪莲在不同海拔梯度上资源分配是否存在权衡关系。旨在深入了解唐古特雪莲适应性形态习性,为有效利用该区域分布的风毛菊属植物特别是该属的雪莲亚属这一重要的药用植物资源提供理论依据。
1 材料与方法
1.1 研究区概况
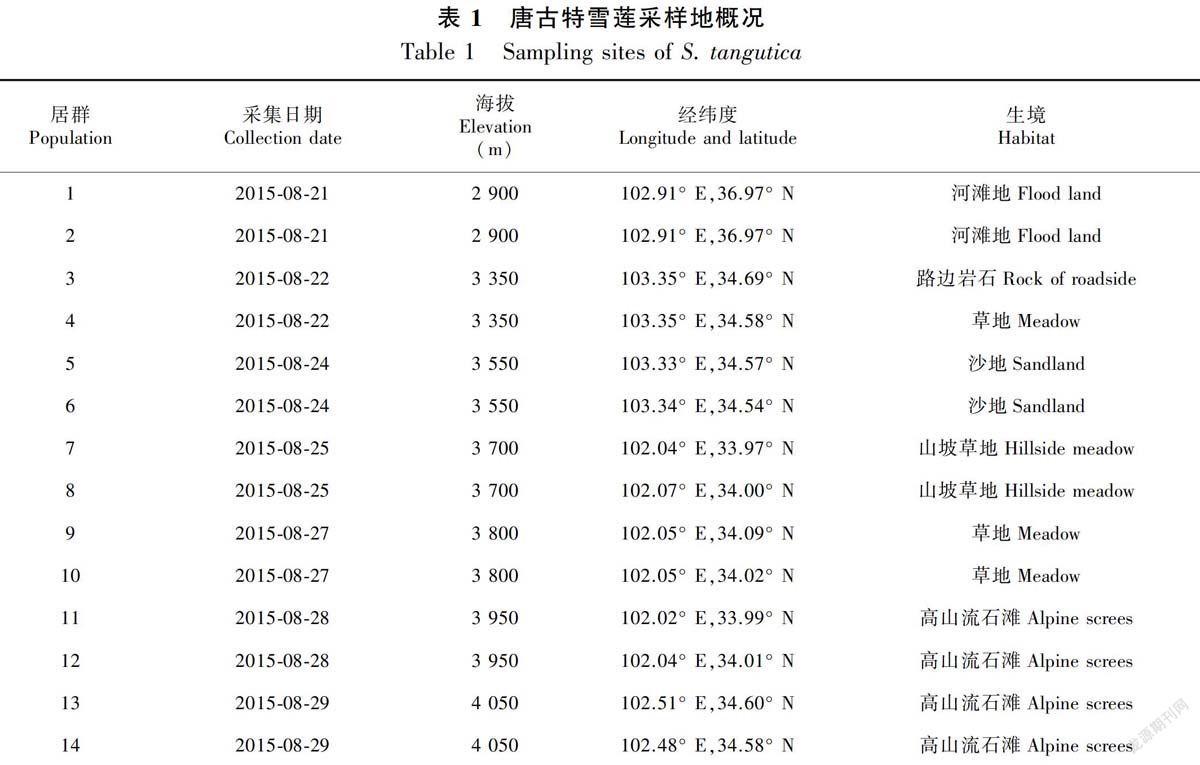
采样点位于青藏高原东缘的甘南藏族自治州合作市,以及夏河、玛曲、卓尼县。样地位置介于102°02′—103°35′ E和33°00′—36°97′ N之间,研究区海拔为1 200~4 800 m,年均气温为1.7 ℃,每年的1月、2月、12月气温较低,7月、8 月气温较高。年降雨量为400~800 mm,无霜期短,日照时间长,是典型的大陆性气候。采样点位于青藏高原东北边缘,高山大川密布,地势险峻多变,地形复杂(王一峰等,2008)。
1.2 研究材料
唐古特雪莲属多年生草本植物,高16~70 cm。根狀茎粗,上部被多数褐色残存的叶柄;茎直立,单生,被稀疏的白色长柔毛,紫色或淡紫色;叶片长圆形或宽披针形,边缘有细齿;头状花序无小花梗,1~5 个,在茎端密集成直径3~7 cm的总花序或单生茎顶。瘦果长圆形,长4 mm,紫褐色。花果期7—9 月。分布于甘肃、青海、四川、云南、西藏。唐古特雪莲种群分布在高海拔地区的风化带和雪线上的石隙,砾石及砂质湿地中,处于潮湿、凉爽以及光照强烈的复杂性气候环境,伴生植物包括草甸雪兔子、小风毛菊、委陵菜等(陈艺林和石铸,1999)。
1.3 研究方法
1.3.1 取样 2015 年8—9月,在甘南藏族自治州从低到高山共选取7 个海拔梯度(2 900、3 350、3 550、3 700、3 800、3 950和4 050 m),每个海拔梯度上选取两个居群,于每个居群内随机挖取25 株完整的样品植株,在野外测量记录植株地上部分高度,同时为了避免野外挖取和运输中对花粉造成损失,每个居群随机选取25 朵未开裂的花药,编号分装入自封袋,然后小心对植株和花药进行分装编号,带回实验室。
1.3.2 生物量的测定 在每个居群25 株完整植株中随机选取25 朵完整小花,经过花部剥离处理后,利用游标卡尺测量并记录每朵小花的花丝和花柱长度,随后计算出每个海拔每个居群中25 朵小花的花丝、花柱长度的平均值,采用血球计数板法对花粉数进行计数,用5 次平均值估算每朵小花的花粉数并记录。将同一植株上的营养器官、管状小花以及去除管状小花的花序轴进行编号。在80 ℃下烘干24 h后用1/10000 电子天平称同一海拔同一居群25 朵管状小花生物量,并计算出一朵管状小花的生物量,同时统计同一植株上的管状小花数目,以及称量每个居群25 朵小花花瓣、雄蕊、雌蕊和茎叶重量。
1.4 统计分析
植株的个体大小以地上部分的总干质量(包括茎、叶、管状小花和花序轴)和株高表示。营养器官生物量为唐古特雪莲茎和叶的总干质量,繁殖器官生物量为唐古特雪莲植株管状小花和花序轴的总干质量。繁殖分配用繁殖器官生物量与地上生物量的比值表示。分别取同一居群25 个植株的地上生物量、繁殖器官生物量、株高、营养器官生物量、同一植株上每个头状花序中的管状小花数目和管状小花生物量的均值用方差分析(ANOVA)分析与海拔间的相关性,同时分析花期管状小花数量及大小、繁殖器官生物量与营养器官生物量、雄蕊重量和雌蕊重量以及花粉数目与花丝长度之间是否存在权衡关系。所有数据均采用Excel 2003和SPSS 20.0软件进行处理和分析,用Origin 8.5作图。
2 结果与分析
2.1 各生物量与海拔的相关性
从图1可以看出,唐古特雪莲的地上生物量与海拔高度呈极显著负相关关系(P<0.01),即随着海拔升高,植株地上生物量变小。一方面由于研究区处于高山低温环境,植物蛋白质新陈代谢及细胞结构合成过程中受到低温限制,成熟细胞结构尤其是细胞壁的合成受阻可能限制了唐古特雪莲的生长发育,从而导致地上生物量降低;另一方面,由于高海拔地区,有效积温低、光合产物积累少,可能也是导致唐古特雪莲的地上生物量与海拔梯度之间呈负相关关系的原因之一。
从图2可以看出,唐古特雪莲的株高与海拔高度均呈极显著负相关关系(P<0.01),即海拔越高,其植株越矮。研究区环境恶劣、生态因子不稳定,唐古特雪莲保持矮小个体更容易被积雪或者碎石保护以避开极端低温环境。
图3结果显示,唐古特雪莲的繁殖器官生物量及营养器官生物量均与海拔呈极显著的负相关关系(P<0.01),海拔升高200 m,唐古特雪莲营养器官生物量平均减少0.212 9 g,繁殖器官生物量平均减少0.009 2 g。这说明随着海拔高度的增加,唐古特雪莲将有限的资源更多的投入到繁殖结构以提高繁殖率。
图4结果表明,唐古特雪莲的管状小花生物量与海拔高度之间呈极显著正相关关系(P<0.01),但管状小花数目与海拔呈极显著负相关关系(P<0.01),海拔升高200 m,唐古特雪莲植株的管状小花数目平均减少3 朵,而管状小花生物量平均增加0.000 317 g。由于研究居群处在高海拔的胁迫环境,唐古特雪莲如果将可获取的有限资源用于形成更多的管状小花,则此过程中不但需要形成繁殖器官还需要消耗额外资源形成总苞片等其他辅助结构,因此唐古特雪莲采取了增加花大小、减少花数量的最优资源分配模式来提高繁殖成功率。
图5结果显示,唐古特雪莲的花瓣重量和花柱长度均与海拔呈极显著正相关关系(P<0.01),即随着海拔升高,花瓣重量越大、花柱越长。唐古特雪莲将更多的资源投入到吸引结构花瓣来增加花期花朵的吸引力,以应对高海拔的不良传粉环境。此外,随着研究区海拔的升高,唐古特雪莲可能是通过增长花柱来提高对传粉昆虫的感受性,有利于传粉昆虫高效传粉。
图6结果表明,唐古特雪莲的花丝长度与海拔呈极显著正相关关系(P<0.01),即海拔越高,花丝越长;花粉数目与海拔呈极显著负相关关系(P<0.01),即随着海拔升高,花粉数目在逐渐减少。一方面,随着海拔的升高,唐古特雪莲可能通过增加花丝长度,将花丝支持的花药托展于更高空间,以利于传粉;另一方面,在高海拔不良环境下,唐古特雪莲将有限的资源更多地投入到花丝,必将导致投资到花粉的资源减少,进而唐古特雪莲可能选择了降低花粉数,提高花粉的质量以更大程度地保证生活史的完成。
2.2 繁殖分配与个体大小的相关性
从图7可以看出,唐古特雪莲的地上生物量与繁殖分配呈极显著负相关关系(P<0.01),即各海拔梯度唐古特雪莲的繁殖分配随着植株地上生物量的增加,呈显著减小趋势。研究区植物可获取的资源量有限,唐古特雪莲对繁殖器官和营养器官的绝对投资均在减少,但对繁殖器官的相对投资多于营养器官,这可能是为了提高生存的概率,将较多的资源用于繁殖来完成生活史。
從图8可以看出,唐古特雪莲在各海拔梯度上的株高与繁殖分配呈显著负相关关系(P<0.01),即随着株高的增大,各海拔梯度唐古特雪莲的繁殖分配呈现出明显的减小趋势。研究区海拔介于2 900~4 050 m,在这个特殊的环境中,唐古特雪莲会受到强风、冰雹等外界因素的干扰,因此,大个体的繁殖代价更高,相应地增加了对繁殖支持结构的投入,从而导致繁殖分配降低。
2.3 各生物量间的权衡关系
从图9可以看出,唐古特雪莲的花期管状小花数目和管状小花生物量呈负相关关系(P<0.01),即唐古特雪莲在管状小花水平上其管状小花数目和管状小花生物量之间存在权衡现象;花期繁殖器官生物量与营养器官生物量呈线性正相关关系(P<0.01),即对营养和繁殖的绝对投资之间存在权衡关系。在一朵小花水平上,唐古特雪莲雌蕊生物量和雄蕊生物量之间呈极显著负相关关系(P<0.01),即唐古特雪莲对雌雄功能间的资源投入存在权衡现象;花粉数与花丝长度呈极显著负相关关系(P<0.01),即花丝越长,花粉数越少,也就是说唐古特雪莲在雄蕊内部对投入到花丝和花粉数的资源也进行了权衡。
3 讨论
3.1 海拔对唐古特雪莲各生物量的影响
植物在生长繁殖过程中受到外界环境资源的限制,通过调节生长与生殖间的资源分配比率来提高适合度以适应不同环境(张婵等,2014),即通过对各器官合理的生物量分配使其更快地适应不同的环境。因此在资源受限环境中,个体会将自身获得的有限资源进行最优化分配以提高适合度(Hutchings,1997)。对不同海拔唐古特雪莲居群的研究显示:随着海拔的升高,植株的个体大小(株高和地上生物量)、繁殖器官生物量、营养器官生物量、管状小花数目均呈线性减小趋势,说明了唐古特雪莲在不同海拔梯度上对资源分配做了适应性调整,与上述理论一致。植物在高海拔地区面临更多复杂且不稳的生态因子的干扰,因此植物的繁殖代价会随着植株个体增大而变高。在环境资源受限的情况下,加强对繁殖支持结构的资源投入,反而会使繁殖分配减小(王一峰等,2012c)。植物越矮小,面临极端环境的干扰就越小。较矮的株高所形成的温暖花部环境由于可以吸引更多的传粉者访花从而大于较高的植物所具备的其它优势(Korner,2001),因此投入到繁殖部分(花)的资源更多(张茜等,2013)。此外,植株距离地面越近,热量在叶冠层的积累就越高(Reekie,1998),所以唐古特雪莲通过减小在高海拔地区植株高度来确保花部的温度,从而提高种子产量并吸引更多传粉昆虫前来访花。对川西风毛菊(S. dzeurensis)8个海拔居群的研究也显示,花期植物个体大小、头状花序数量、繁殖器官及营养器官生物量均与海拔呈线性递减趋势(王一峰等,2015)。
特殊的花部综合特征对提升植物传粉效率并促进异交繁殖成功具有重要意义(Wang,2011)。多数由昆虫传粉的植物都是通过华丽的花展示来吸引传粉昆虫前来传粉,从而完成有性生殖。随着研究区海拔的升高,环境因子复杂多变,传粉昆虫越来越少,唐古特雪莲可以通过加强花朵的吸引力来应对不良的传粉环境。本研究发现,随着海拔的升高,唐古特雪莲的花瓣重量增大,花柱、花丝变长,而花粉数量明显减少。一方面,在高海拔地区,唐古特雪莲将更多地资源投入到吸引结构花瓣以此来加强花朵对传粉昆虫的感受性,相对较长的花柱对于接受花粉越有利(王一峰等,2008),进而加大了花粉的输入和输出,这在一定程度上弥补了随着海拔升高因传粉昆虫数量减少等不利因素带来的影响,从而保证了传粉昆虫对花粉的有效传播;另一方面,在植物可获取的资源受限的情况下,唐古特雪莲通过减少花粉数,提高花粉的质量,最大程度保证花粉的生殖活力,以确保其生活史的顺利完成。
3.2 个体大小对繁殖分配的影响
植株个体增大会导致有限资源更多地投入到繁殖支持结构,相对的繁殖分配会降低(Reekie,1998)。当植物的资源获取受到限制,植物对茎、叶等营养器官的资源投入量增大,就意味着对繁殖器官的相对投入减少,从而会影响繁殖分配及个体的存活。本研究结果表明,个体大小(株高和地上生物量)对各海拔梯度唐古特雪莲的繁殖分配影响显著,即个体越大各海拔唐古特雪莲的繁殖分配越小。随着研究区海拔的升高,唐古特雪莲通过减小个体大小来降低资源消耗,把有限的资源投入到繁殖器官,目的是为了顺利完成有性生殖过程。对长毛风毛菊(S. hieracioides)、柳叶菜风毛菊(S. epilobioides)、尖苞风毛菊(S. subulisquama )、钝苞雪莲(S. nigrescens)等植物繁殖分配与海拔差异研究的结果也支持这一结论(索南措等,2013;王一峰等,2012c)。个体越大,对繁殖器官的绝对投入越低,其相对的繁殖分配也就越低。因此个体较小的植物其繁殖分配反而相对较高(王一峰等,2012a)。研究发现,繁殖分配依赖于植株个体大小(Zhang & Jiang,2002)。如对蔷薇科(Rosaceae)的直立委陵菜(Potentilla recta)(Soule & Werner,1981)和毛茛科的欧洲金莲花(Trollius europaeus)(sam & Staffan,1998)的研究表明,繁殖分配都随个体的增大而减小。造成这种现象的原因可能是由于唐古特雪莲在青藏高原东缘恶劣环境中,植株个体越大越容易受到外界因素如动物踩踏、取食及天气变化的影响。个体越大的植株投入到繁殖支持结构的资源越多,其结果就是使植物的繁殖分配减小。该研究结果符合以往对植物繁殖分配研究的结论。本研究结果也与植物繁殖分配与个体大小间的线性关系模型一致(Weiner & Lovett,1988)。
3.3 各组生物量间的权衡关系
权衡理论的主要内容是植物将可获取的有限资源用于一种功能就不能再用于其他的功能或结构,如植物增加对繁殖器官生物量的资源投资,相应会减少对其他功能和结构的投入(孟丽华等,2011)。对一年生龙胆属(Gentiana)植物研究发现,当用偏相关方法控制植物个体大小后,植物的花大小与花数目之间表现出“此消彼长”权衡关系(陈学林等,2009)。本研究表明,唐古特雪莲在植株总生物量下降的情况下,有选择性的减少管状小花数量、营养器官生物量,将有限的资源投入到每个管状小花的大小和繁殖器官上,有利于完善花部特征的构件以及种群繁衍,尽可能地保证生殖生长的顺利完成。植物花大小—数量权衡机制是植物在异质生境下各功能可塑性对外界环境适应性的表现(王永健等,2010)。
随着海拔的升高,唐古特雪莲对雄蕊和雌蕊的投入存在权衡关系。本研究结果表明,对雌蕊投入增加,是为了使花柱变长,从而使得柱头更多地暴露在环境中,进而会使传粉昆虫更有效地将花粉进行传播,在各种花部特征中,柱头的伸出是通过影响所接受花粉的质量和数量来影响雌性繁殖成功,这或许是自然选择的一个目的(Campbell, 1989)。与传粉者大小相适应的高高伸出的柱头就是为了有效的傳粉,特别是在有管状花的植物中更是如此(Wang et al, 2013)。一方面在高海拔区域,虽然传粉昆虫活动量会减少,但唐古特雪莲通过花柱的伸长,即提高柱头对传粉昆虫的感受性得以补偿;另一方面随着海拔的升高,在外界环境资源有限的情况下,唐古特雪莲为了保证种群的生存和繁衍,如果产生大量的花粉而不能全部被传粉昆虫携带传播,势必会造成有限资源的浪费,因而将有限的资源更多地投入到增加花瓣质量和增长花柱、花丝上,以此来增加传粉昆虫每次访花、携带花粉的几率,使传粉过程能顺利完成,从而有效地提高有性生殖繁殖的成功率。本研究结果符合性分配理论假定的对于其中一个性别资源投入增多,必然会导致对另一种性别资源投入的减少。
4 结论
(1)随着海拔的升高,植物可获取的资源总量降低,唐古特雪莲分配给各个器官的资源总量减少。唐古特雪莲的这种适应性形态特征很有可能就是通过海拔影响植株个体大小变化来完成的。(2)随着海拔的升高,唐古特雪莲通过伸长花柱、花丝,减少花粉数,以及投入更多的资源于传粉吸引结构花瓣来响应随着海拔变化的生境,从而顺利地完成有性繁殖。(3)唐古特雪莲通过对管状小花数量及大小、繁殖器官生物量与营养器官生物量、雄蕊重量和雌蕊重量以及花粉数目与花丝长度之间进行权衡,将可获取的有限资源进行合理分配,从而适应了青藏高原东缘这样恶劣的高山环境。
参考文献:
SAM HP, STAFFAN K, 1998. Altitudinal variation in size effects on plant reproductive effort and somatic costs of reproduction [J]. coscience, 5(4):517-525.
CAMPBELL DR, 1989. Measurements of selection in a hermaphroditic plant: variation in male and female pollination success [J]. Evolution, 43:318-334.
CHEN XL, LIANG Y, WEI QI, et al, 2009. Studies on reproductive allocation, floral size and its trade-off with floral number of annual Gentiana [J]. Acta Pratacult Sin,(5):58-66. [陈学林,梁艳,齐威,等,2009. 一年生龙胆属植物的繁殖分配及其花大小、数量的权衡关系研究 [J]. 草业学报, (5):58-66.]
CHEN YL,SHI Z,1999. Flora Reipublicae Popularis Siniceae [M]. Beijing: Science Press: 78. [陈艺林,石铸,1999. 中国植物志 [M].北京:科学出版社: 78.]
HAN B, WANG FX, HUANG SQ, 2011. Production of male flowers does not decrease with plant size in insect-pollinated Sagittaria trifolia, contrary to predictions of size-dependent sex allocation [J]. J Syst Evol, 49(5):379-385.
HUTCHINGS MJ, 1997. Resource allocation patterns in clonal herbs and their consequences for growth [M]. Plant Resourc Alloc: 161-189.
KLEUNEN MV, RITLAND K,2010. Predicting evolution of floral traits associated with mating system in a natural plant population [J]. J Evol Biol, 17(6):1389-1399.
KORNER C, 2001. Alpine plant life: Functional plant ecology of high mountain ecosystems [J]. Mount Res Dev, 21(2):202-202.
LI XR, TANG X, FU WJ, 2016. Floral syndrome and breeding system of Gymnocarpos przewalskii Maxim [J]. Chin J Ecol, 35(10):2592-2598. [李新蓉,唐欣,付文潔,2016. 荒漠孑遗植物裸果木的花部特征及繁育系统 [J]. 生态学杂志, 35(10):2592-2598.]
LIANG Y,2009. Reproductive allocation strategies of Gentiana in the eastern of Qinghai-Tibetan Plateau [D]. Lanzhou: Northwest Normal University. [梁艳,2009. 青藏高原东缘龙胆属植物繁殖分配对策研究 [D]. 兰州:西北师范大学.]
MA YX, WANG YF, LI YY, et al, 2017. Correlation between reproductive allocation and elevation of Saussurea graminea in eastern margin of the Tibetan Plateau [J]. Bull Bot Res, 37 (1):23-30. [马诣欣,王一峰,李怡颖,等,2017. 青藏高原东缘禾叶风毛菊的繁殖分配与海拔的相关性 [J]. 植物研究,37(1):23-30.]
MENG LH, WANG ZK, LIU CY, et al, 2011. Reproductive allocation of an alpine perennial, polygonum macrophyllum [J]. Acta Bot Boreal-Occident Sin, 31(6):1157-1163. [孟丽华,王政昆,刘春燕,等,2011. 高山植物圆穗蓼的繁殖资源分配 [J]. 西北植物学报,31(6):1157-1163.]
PIRES JPDA, FREITAS L, 2008. Reproductive biology of two tree species of Leguminosae in a Montane Rain Forest in southeastern Brazil [J]. Flora, 203(6):491-498.
REEKIE EG,1998. An explanation for size-dependent reproductive allocation in Plantago major [J]. Can J Bot,76:43-50.
SOULE JD, WERNER PA, 1981. Patterns of resource allocation in plants, with special reference to Potentilla recta L [J]. Bull Torr Bot Club, 108(3):311-319.
SU M, QI W, YANG M, et al, 2009. Altitudinal differences in flower traits and reproductive allocation of Delphinium pylzowii (Ranunculdiculdiceae) at east Qinhai Tibetan Plateau [J]. J Lanzhou Univ(Nat Sci Ed ),45(2):61-65. [苏梅,齐威,阳敏,等, 2009. 青藏高原东部大通翠雀花的花特征和繁殖分配的海拔差异 [J]. 兰州大学学报(自然科学版),45(2):61-65.]
SU ZX, ZHONG ZC, 1998. Studies on the reproductive ecology of gordonia acuminata populationⅡ.The patterns of reproductive allocation on the biomass in the population [J]. Acta Ecol Sin,(4):45-51. [苏智先,钟章成,1998. 四川大头茶种群生殖生态学研究Ⅱ.种群生物量生殖配置格局研究 [J]. 生态学报,(4):45-51.]
SUN MM, KARIN G, FLORIAN PS, 2014. Floral adaptation to local pollinator guilds in a terrestrial orchid [J]. Ann Bot, 113:289-300.
SUO NC, WANG YF, LI M, et al, 2013. Reproductive allocation of Saussurea hieracioides(Asteraceae), a common species in eastern margin of Qinghai-Tibet Plateau [J]. Chin J Ecol, 32(6):1433-1438. [索南措,王一峰,李梅,等,2013. 青藏高原东缘常见种长毛风毛菊的繁殖分配 [J]. 生态学杂志, 32(6):1433-1438.]
WANG F, ZHAO YH, YANG SY, et al. 2013. Floral syndrome and its pollination system of Tamarindus indica [J]. J W Chin For Sci, 42(40):62-72.
WANG L, 2011. Floral syndrome and secondary pollen presentation in Codonopsis clem-atidea(Campanulaceae) [J]. Biodivers Sci, 19(1):24-33.
WANG Y, HU LJ, DUAN YW, et al, 2010. Altitudinal variations in reproductive allocation of Bergenia purpurascens(Saxifragaceae) [J]. Acta Bot Yunnan, 32(3):270-280. [王赟,胡莉娟,段元文,等,2010. 岩白菜(虎耳草科)不同海拔居群的繁殖分配 [J]. 云南植物研究, 32(3):270-280.]
WANG YF, GAO HY, SHI HY, et al, 2008. Adaptive significance of Saussurea parvifloras sexual organs,qinghai-tibetan plateau [J]. Chin J Plant Ecol, (2):379-384. [王一峰,高宏岩,施海燕,等,2008. 小花风毛菊的性器官在青藏高原的海拔变异 [J]. 植物生态学报, (2):379-384.]
WANG YF, JIN J, HOU HH, et al, 2015. Changes in flowering resource allocation of Saussurea dzeurensis with elevations [J]. Chin J Plant Ecol, 39(9):901-908. [王一峰,靳洁,侯宏红,等, 2015. 川西风毛菊花期资源分配随海拔的变化 [J].植物生态学报, 39(9):901-908.]
WANG YF, LI M, LI SX, et al, 2012a. Variation of reproductive allocation along elevations in Saussurea stella on east Qinghai-Xizang Plateau [J]. Chin J Plant Ecol, 36(11):1145-1153. [王一峰,李梅,李世雄,等,2012a. 青藏高原东缘星状风毛菊生殖分配对海拔的响应 [J]. 植物生态学报, 36(11):1145-1153.]
WANG YF, LIU QQ, PEI ZY, 2012c. Correlation between altitude and reproductive allocation in three Saussurea species on Chinas Qinhai-Tibetan Plateau [J]. Chin J Plant Ecol, 36(1):39-46. [王一峰,刘启茜,裴泽宇,等,2012c. 青藏高原3种风毛菊屬植物的繁殖分配与海拔高度的相关性 [J]. 植物生态学报, 36(1):39-46.]
WANG YF, PEI ZY, LIU QQ,2012b. Correlation analysis between altitude and reproductive allocation Floral traits in Saussurea iodostegia Hance [J]. J NW A & F Univ(Nat Sci Ed), (6):195-200. [王一峰,裴泽宇,刘启茜,2012b. 紫苞雪莲花期繁殖分配及花部特征与海拔高度的相关分析 [J]. 西北农林科技大学学报(自然科学版), (6):195-200.]
WANG YJ, FANG X, ZHONG ZC, et al, 2010. Effect of different habitats on floral,fruit and seed traits of Iris japonica Thunb [J]. Acta Ecol Sin, 30(17):4628-4635. [王永健,方兴,钟章成,2010. 不同生境对蝴蝶花花部与果实特征的影响 [J]. 生态学报,30(17):4628-4635.]
WEINER J, LOVETT DJ, 1988. The influence of competition on plant reproduction [J]. Plant Reprod Ecol Patt Strat: 228-245.
WRIGHT JW, MEAGHER TR, 2004. Selection on characters in natural Spanish populations of Silene latifolia [J]. J Evol Biol, 17:382-395.
YANG Y, LUO JT, ZHANG BF, et al, 2015. Studies on floral characteristics and breeding system of Paeonia decomposita [J]. J Plant Resourc Environ, 24(4):97-104. [杨勇, 骆劲涛,张必芳,等,2015. 四川牡丹的花部特征和繁育系统研究 [J]. 植物资源与环境学报, 24(4):97-104.]
ZHANG C,HU LJ,WANG Y, 2014. Reproductive allocation of three sympatric Comastoma(Gentianaceae)species[J].Guihaia, 34
(1):38-45. [张婵,胡莉娟,王赟,2014. 三种同域分布喉毛花的繁殖分配 [J]. 广西植物, 34(1):38-45.]
ZHANG DY, JIANG XH, 2002. Size-dependent resource allocation and sex allocation in herbaceous perennial plants [J]. J Evol Biol, 15(1):74-83.
ZHANG Q, ZHAO CZ, MA XL, et al, 2013. Response of reproductive allocation of Stellera chamaejasme population in alpine grassland to altitude [J]. Chin J Ecol, 32(2):247-252. [张茜,赵成章,马小丽,等, 2013. 高寒草地狼毒种群繁殖分配对海拔的响应 [J]. 生态学杂志, 32(2):247-252.]
ZHANG TF,ZHANG Y, LI YJ, 2014. Flower duration and reproductive allocation in different altitudinal populations of Aconitum gymnandrum(Ranunculaceae) [J]. Acta Bot Boreal-Occident Sin, 34(8):1576-1580. [张挺峰,张勇,李亚珺, 2014. 青藏高原露蕊乌头不同海拔居群的开花持续期与繁殖分配 [J]. 西北植物學报, 34(8):1576-1580.]
