北方农牧交错区不同土地利用方式下土壤呼吸动态特征
2018-09-03杨新明
杨新明,韩 磊,庄 涛*
(1.济南市环境研究院土壤研究所,济南 250100;2.中国农业大学资源与环境学院,北京 100193)
土地利用方式的改变能够显著影响陆地生态系统与大气之间的碳交换过程[1],是导致大气CO2浓度不断升高的主要原因之一[2]。开垦和放牧是草地生态系统主要的土地利用方式,已有研究表明草地开垦成农田后,施肥、耕作等相关的农业措施将使土壤微环境条件、微生物群落的组成和活性等发生改变,进而改变土壤呼吸[3-8]。Qi等[4]在内蒙古典型草地研究表明,开垦30年后的农田土壤呼吸累积量仅为草地的68%~77%;巴西中部的热带草原转变为农田以后,玉米-大豆轮作地土壤呼吸累积量达草地的2倍,可能是农田大量施肥所致[8]。放牧对草地土壤呼吸的影响的研究结果也不一致[9-11]。马涛等[9]在内蒙古锡林河流域的研究结果表明,放牧导致羊草土壤呼吸年总量降低34%;在北美温带草地,长期重度放牧和中度放牧草地土壤呼吸没有显著差异。这可能是两个样地土壤物理化学性质的不同引起的[11]。因此,由于草地生态系统的微环境条件、有机质输入、土壤微生物等因素的差异,土壤呼吸对土地利用方式转变的响应也是不同的。
我国北方农牧交错区是华北湿润-半湿润农区和内蒙古干旱-半干旱草地接壤的过渡地带,这个区域对我国农牧业生产和生态环境保护具有重要的意义。长期以来,过度的开垦和放牧导致此区域的草地大面积退化。近十多年来,在北方农牧交错区,禁牧、适度放牧等集约化的草地管理方式已经普遍用来恢复这些退化草地[12],管理方式的改变也导致土地利用方式发生变化。然而,在北方农牧交错区,土地利用方式转变对土壤呼吸影响尚缺乏定量的认识。本研究以河北省沽源县农牧交错区为例,选择当地典型的土地利用类型,研究不同土地利用方式间土壤呼吸的动态变化特征及其与水热条件的响应关系,旨在进一步深化对土壤呼吸的时空变化规律的认识,对揭示北方农牧交错区生态系统碳循环具有重要的意义。
1 材料和方法
1.1 研究区概况及试验设置
试验地点位于河北沽源国家草地生态系统野外观测研究站(41°46′N,115°41′E),平均海拔 1380 m。该区域位于华北农牧交错带中部,属于半干旱大陆季风气候,年均温1℃,最低月(1月)平均气温-18.6℃,最高月(7月)平均气温17.6℃,年平均降水量380 mm,主要集中于7—9月,占全年降水量79%。该区域冬季冰雪覆盖时间达半年以上,生长季一般为从6月开始到9月结束,主要土壤类型为栗钙土。
本研究选择三种土地利用类型。①自由放牧草地(FG):面积大约为15 hm2,2008年开始适度放牧,放牧强度为每年1.0只羊,之前为中度退化草地,优势种为羊草(Leymus chinensis,占物种组成的73%),其他物种有克氏针茅(Stipa krylivii,10%)和星毛委陵菜(Potentilla acaulis,8%);②围封草地(UG):面积大约为3 hm2,自从1997年开始围封禁牧,围封之前为中度退化草地,目前植被恢复良好,覆盖度约为90%,禁牧草地的优势种为羊草(Leymus chinensis,占物种组成的90%),其他物种有克氏针茅(Stipa krylivii,8%)、星毛委陵菜(Potentilla acaulis,2%);③莜麦农田(AN):属于雨养农田,1995年由草地开垦而来,以种植莜麦(Avena.nuda)为主,管理比较粗放,农田作物在6月初播种,9月下旬收获。播种之前施用农家肥做基肥(约30 kg N·hm-2和97 kg C·hm-2)。放牧草地、围封草地、莜麦农田的地上部生物量分别为56、426、336 g·m-2,根系生物量分别为1392、1933、737 g·m-2。三个样地0~20 cm土壤的基本性质见表1。
1.2 土壤呼吸速率及环境因子的测定
在放牧草地、围封草地、莜麦农田三个样地中,选择地势平坦,植物长势均匀的区域均设置5个面积1 m×1 m的样方进行土壤呼吸的测定,样方之间的距离为5 m。每个样方内布置1个聚氯乙烯环(PVC环,内径为20 cm)用来测定土壤呼吸,每个样地内5个PVC环的测定结果平均值代表该样地的土壤呼吸。每次测定时提前一天沿地面剪去环内的植物。
于2015年6月1日至9月30日的整个生长季期间,采用LI-8100A土壤呼吸通量测定系统(Li-COR,Lincoln,NE)测定土壤呼吸,Offset(PVC环露出地面的高度)设定为5 cm。每天9:00—10:00按顺序完成所有样地的测定,每个PVC环测定的时间为2 min。在没有发生强降雨的情况下,土壤呼吸速率的测定频率为每周2次,当有强降雨发生时,每天测定1次土壤呼吸,持续3~5 d。7月10日、8月10日、9月29日在每个样地中选择固定的PVC环进行土壤呼吸速率的日变化观测,白天每2 h、夜晚每3 h测定一次,持续时间24 h。

表1 不同土地利用方式样地0~20 cm土壤基本性质Table 1 Soil properties of 0~20 cm depth in study sites for different land uses
测定土壤呼吸的同时,分别用LI-8100A测定系统的长杆土壤温度计和土壤水分测定仪(ThetaKit,Delta-T Devices,Cambridge,UK)测定附近10 cm土壤温度和0~6 cm土壤体积含水量。土壤水分测定仪预先在实验室内进行了标定,用来保证土壤含水量测定的准确性。
1.3 数据分析
用SigmaPlot 10.0软件描述不同土地利用方式下土壤呼吸速率动态变化和日变化图。用SPSS 10.0软件拟合土壤呼吸速率(F)与10 cm土壤温度(T)、0~6 cm土壤含水量(W)之间的单变量土壤呼吸模型。土壤温度是影响土壤呼吸的主导因素,两者之间的关系常用指数方程来表示[7]。用非线性模型来描述土壤水分与土壤呼吸的相应关系[13]:

式中:F为土壤呼吸速率,μmol·m-2·s-1;T为10 cm土壤温度,℃;W为0~6 cm土壤湿度,cm3·cm-3;Q10是温度每增加10℃所造成的呼吸速率改变的倍数,表示土壤呼吸对温度的敏感性。A、B、a、b、c都是拟合常数。
整个生长季土壤呼吸的累积量参考Salimon等[14]所用的线性内插法进行计算。
2 结果与分析
2.1 不同土地利用方式下土壤温度和含水量的变化
由图1a可知,放牧草地(FG)、围封草地(UG)、莜麦农田(AN)三种土地利用类型中,10 cm土壤温度的变化趋势基本一致。在生长季期间,7月和8月土壤温度较高,6月和9月土壤温度较低。在观测期间,三个样地土壤含水量的波动主要与降雨事件有关(图1b)。三个样地平均温度分别为15.2、14.6、13.0℃,其中6月份农田和草地的差异最大,9月份差异最小。而土壤平均含水量分别为 0.24、0.22、0.14 cm3·cm-3;两个草地样地土壤含水量明显高于农田。造成这种差异的原因与地表植被覆盖有关,农田地表植被覆盖度低,能够接受的太阳辐射多,导致土壤温度较高,同时地面蒸发大,导致土壤含水量较低。
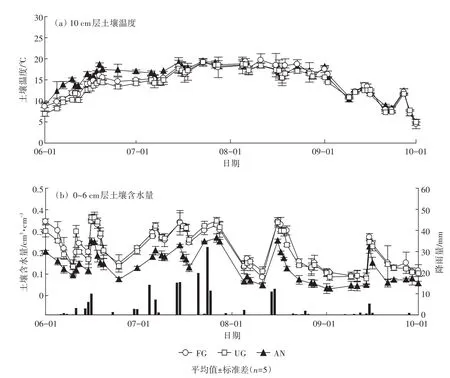
图1 放牧草地(FG)、围封草地(UG)、莜麦农田(AN)三个样地土壤温度及含水量的变化动态Figure 1 Dynamic trends of soil temperature and soil moisture in FG,UG and AN sites under three land uses
2.2 不同土地利用方式下土壤呼吸速率的变化
由图2可知,生长季期间,三个样地的土壤呼吸动态变化一致。7月和8月,土壤水热条件适宜,植物生长旺盛,根系和微生物活性增强,土壤呼吸速率较高;6月和9月,土壤温度和含水量相对较低,尤其从8月下旬开始,随着土壤温度的降低,根系活力和微生物活性降低,土壤呼吸速率开始快速下降。生长季期间,FG、UG、AN土壤呼吸速率的动态变化范围分别为0.54~9.04、1.26~7.68、1.12~9.38 μmol·m-2·s-1,土壤呼吸速率均值分别为3.41、4.06、3.58 μmol·m-2·s-1。
降雨导致了土壤呼吸的波动,尤其是强降雨。以6月15—16日为例:两天降雨量为15.9 mm,降雨前,FG、UG、AN 土壤呼吸速率分别为 2.09、3.06、2.88 μmol·m-2·s-1;降雨后1 d,土壤呼吸速率出现不同程度的抑制,下降至0.54、1.62、1.12 μmol·m-2·s-1;接下来3~4 d内,土壤呼吸速率快速增加并出现峰值,FG、UG、AN土壤呼吸速率峰值分别为4.24、4.88、4.14 μmol·m-2·s-1。再以8月14日—15日的23.1 mm的降雨为例,降雨前,FG、UG、AN土壤呼吸速率分别为4.48、4.52、3.67 μmol·m-2·s-1,降雨后土壤呼吸速率明显地增加直至达到峰值,FG、UG、AN土壤呼吸速率分别达到9.04、6.60、8.21 μmol·m-2·s-1。
2.3 土壤呼吸速率的日变化特点
由图3和图4可知,土壤呼吸速率的日动态均呈单峰型曲线,一般在10:00—14:00达到最大值,而夜间0:00—6:00之间出现最低值。以7月10日为例:FG和UG的最大值出现在12:00,日变化幅度分别为3.88(3.74~7.62)、2.44(4.21~6.65)μmol·m-2·s-1,AN在14:00 达到最大值,变化幅度为 2.61(3.58~6.19)μmol·m-2·s-1。
土壤呼吸速率与10 cm土壤温度日变化动态均出现不同步的现象。以7月10日为例:FG、UG、AN的土壤呼吸速率分别在12:00、12:00、14:00达到最大值,而相应土壤温度都在16:00达到最大值。由图4可知,9月29日,FG、UG、AN的土壤呼吸速率都在10:00达到最大值,而相应土壤温度分别在14:00、16:00、14:00达到最大值。9月29日0:00和9月30日6:00,AN土壤呼吸速率出现负值,分别为-0.15、-0.08 μmol·m-2·s-1。
2.4 土壤呼吸速率与环境因子的关系
土壤呼吸速率与土壤温度、土壤含水量的拟合结果如表2所示。土壤温度能够解释土壤呼吸速率变化的 43%~69% 。FG、UG、AN 的 Q10值分别为 3.8、3.0、4.7,并且三个样地Q10值存在显著性差异(p<0.05),可见UG土壤呼吸速率对土壤温度变化的敏感性最低。与温度相比,土壤含水量对土壤呼吸速率的影响相对较低,FG土壤含水量仅可解释土壤呼吸速率变化的8%,UG和AN土壤含水量则可以解释土壤呼吸速率变化的27%。
2.5 不同土地利用方式下土壤呼吸累积量
三种土地利用方式下土壤呼吸累积量如图5所示。整个生长季,FG、UG、AN三个样地土壤呼吸累积量分别为644、766、661 g·m-2,其中UG土壤呼吸累积量显著高于FG和AN(p<0.05),而FG和AN土壤呼吸累积量之间没有显著差异。


图3 7月10日和8月10日三种样地放牧草地(FG)、围封草地(UG)、莜麦农田(AN)土壤呼吸速率的日变化动态Figure 3 Diurnal variation of soil respiration(a,b)and soil temperature at 10 cm depth(c,d)in FG,UG,AN sites during July 10,August 10

图4 放牧草地(FG)、围封草地(UG)、莜麦农田(AN)土壤呼吸速率和10 cm土壤温度的日变化动态(9月29日)Figure 4 Diurnal variation of soil respiration(a)and soil temperature at 10 cm depth(b)in FG,UG,AN sites during September 29

表2 土壤呼吸速率与土壤温度和湿度的拟合参数Table 2 Parameters of relation model of soil respiration with soil temperature and moisture under three types of land-use

图5 生长季期间放牧草地(FG)、围封草地(UG)、莜麦农田(AN)土壤呼吸累积量的比较Figure 5 Comparison of accumulative soil respiration under three types of land-uses during the growing season
3 讨论
3.1 土壤呼吸速率变化和日变化
观测期间草地和农田土壤呼吸速率的变化趋势呈现出单峰型特点,与许多研究结论相似[15-17]。尽管土地利用方式、测定方法和时间、气候的差异等因素导致土壤呼吸出现的峰值时间有所不同,但是整个生长季的动态变化基本一致。
相关研究结果表明,土壤呼吸速率的日动态与土壤温度基本保持同步[10,18],然而我们发现土壤呼吸速率比土壤温度提前2~3 h达到最大值。土壤温度的日变化与土壤呼吸速率不同步,相似的现象[19-20]已有报道,目前对这种不同步的现象还没有明确的结论,可能的解释是土壤呼吸的不同组分对温度的敏感性不同[19]。土壤呼吸包括自养呼吸(主要是根系呼吸)和异养呼吸(主要是微生物分解有机质产生CO2)[21],大多数情况下,异养呼吸的日变化主要受土壤温度的调控,因为微生物对有机质分解与温度有关[22]。自养呼吸对温度的敏感性受到光合同化产物的影响[23],光合同化产物很大程度上受到太阳辐射的调控[24]。根据当地气象站的观测数据,太阳辐射一般在上午11:00左右达到最大值,光合产物转运到根系需要一定的时间,自养呼吸可能在11:00之后会达到最大值。异养呼吸与土壤温度同步在16:00达到最大值,因此土壤呼吸最大值出现在太阳辐射和土壤温度的峰值之间,即11:00—16:00之间的时段内。
在9月30日的土壤呼吸日动态中,农田(AN)的土壤呼吸速率在凌晨出现负值,这种现象在其他研究中也出现过[25]。其原因可能是:凌晨土壤温度很低(低于5℃),土壤微生物和根系呼吸基本停止,导致土壤孔隙中的CO2浓度低于大气中CO2浓度,在大气与土壤CO2浓度梯度的驱动下,大气中的CO2向土壤扩散并被土壤固定。此种现象提醒我们在估算区域土壤呼吸总量时,必须注意低温期负通量的影响。
3.2 降雨对土壤呼吸的影响
降雨对土壤呼吸的影响机制非常复杂,陈全胜[13]等把降雨机制分为替代效应、阻滞效应、微生物刺激效应、微生物生物量的激增效应。我们观察到6月15—16日的强降雨短时间抑制了土壤呼吸,张红星[26]等在我国黄土区观测到了强降雨发生后土壤呼吸速率减弱的情况。这是因为阻滞效应,水分取代了土壤中CO2占据的空间,同时土壤的通透性也会变差,CO2在土壤中的扩散阻力因此增大,导致雨后实际测定的土壤CO2排放量减少。然而8月14—15日的强降雨后土壤呼吸速率则是明显地增强,土壤呼吸速率在降雨后增强的原因可能存在于两方面:其一是替代效应,降雨填充了土壤孔隙,把土壤中的CO2气体置换出来,土壤CO2释放在水分改善后的几分钟内就会发生,可能持续几个小时;其二,长期干旱导致土壤微生物活性降低,土壤呼吸较弱,降雨增加了对基质的利用率,激活了微生物活性,进而促进土壤呼吸[27]。本试验采用手动间歇方式观测土壤呼吸,可能无法观测到替代效应,为深入探讨降雨对土壤呼吸的影响机制,原位高频率观测土壤呼吸是非常必要的。
3.3 环境因子对土壤呼吸的影响
土壤呼吸温度敏感性Q10值是反映土壤呼吸对温度变化敏感性的重要指标。本研究中农田土壤呼吸对温度的敏感性大于草地,与马骏等[18]的研究结论一致,表明在未来气候变暖的背景下,农牧交错区草地开垦为农田可能会加速土壤有机碳的损失。除了FG外,土壤含水量对土壤呼吸的解释作用为26%~27%并达到极显著水平,说明土壤含水量同样能够影响土壤呼吸。在内蒙古草地生态系统中,马涛等[9]研究表明0~10 cm土壤含水量能够解释围栏封育样地土壤呼吸变异的87%,放牧样地10~20 cm和20~30 cm土壤含水量可以解释放牧地土壤呼吸变异74%。
3.4 土地利用方式对土壤呼吸累积量的影响
本研究结果表明,在北方农牧交错区,开垦和放牧降低草地土壤呼吸累积量,与其他的研究结论一致。马涛等[9]研究表明,放牧草地土壤呼吸总量比围封草地少的主要原因是放牧减少了地下生物量及根系分泌物,降低了根系呼吸和微生物呼吸。Frank等[3]发现草地开垦为农田后,农田土壤呼吸总量仅为草地的57%,同时也认为土壤微生物量碳、根系生物量的差异是造成农田和草地土壤呼吸差异的主要原因。本研究中农田的土壤呼吸累积量要低于围封草地,一方面是由于围封草地的根系生物量是农田的2.6倍,根系呼吸和微生物呼吸都要高于农田[9],另一方面,围封草地土壤含水量是农田的1.7倍,有利于增强土壤呼吸。围封草地的地上部、地下部生物量分别是放牧草地的7.6、1.4倍,放牧草地土壤呼吸累积量低于围封草地,主要原因可能是放牧减少了地表植被量和地下部生物量,降低土壤呼吸[28]。
4 结论
(1)三种土地利用方式下土壤呼吸速率呈现出相似的动态变化趋势,土壤温度是主要的调控因子。降雨导致土壤呼吸的波动,而降雨的效应不同,6月15—16日的强降雨短时间抑制了土壤呼吸,8月14—15日降雨增加了土壤呼吸速率。
(2)三种土地利用下土壤呼吸速率的日动态呈现出单峰型变化,土壤温度的日变化要滞后土壤呼吸速率2~3 h达到最大值。在9月30日凌晨,农田土壤呼吸速率出现负值的现象,提醒我们在估算区域土壤呼吸总量时,必须注意负通量的影响。
(3)土壤温度和土壤含水量都能够显著影响土壤呼吸的动态变化。农田样地的Q10值要大于其他两个草地样地,说明未来气候变暖情景下,草地开垦成农田会加快农田土壤有机碳损失量。
(4)在北方农牧交错区,开垦和放牧降低土壤呼吸累积量。农田的土壤呼吸累积量要低于围封草地,主要是两者之间根系生物量和土壤含水量的差异导致的,而放牧草地土壤呼吸低于围封草地,主要原因可能是放牧减少地表植被量和地下部生物量,降低土壤呼吸。
