草地早熟禾叶片脂肪酸对秋季低温的响应
2018-08-23张咏梅
张咏梅, , ,
(1. 甘肃农业大学 甘肃省干旱生境作物学重点实验室, 甘肃 兰州730070; 2. 甘肃农业大学草业学院, 甘肃 兰州730070;3. 甘肃农业大学园艺学院, 甘肃 兰州730070)
草地早熟禾(PoapratensisL.)为禾本科早熟禾属多年生草本植物,其坪质优美、质地纤细、绿期长、再生能力和耐修剪性强,喜光耐阴、喜温暖湿润,又具有很强的耐寒能力,适用性广[1],是宝贵的草坪草和牧草种质资源[2-3]。草地早熟禾广泛分布于寒带和北温带湿润的冷凉地区[4-5],是天然草原、草甸以及人工草地植被的重要组成成分[6]。其作为一种典型的冷季型草坪草,具有较强的抗寒能力,夏季炎热时生长停滞,春秋生长茂盛。通常在4—6月生长最旺盛,其次是9—10月初;而在3月和11月受寒冷的限制,7月和8月受干热限制,生长停顿。
细胞膜是细胞损害的最敏感部位。因此,研究逆境对植物生长的影响时往往将细胞膜渗漏[7]、细胞膜电位[8-9]、细胞膜成分脂肪酸[10-12]或蛋白质[13]作为主要的研究内容。使细胞膜的研究一度成为植物抗性生理学研究的热点。植物细胞膜必须要维持一定的流动性和完整性,才能使细胞膜的一系列功能得到发挥,从而维持一个良好的细胞内环境,使细胞得以生存。脂肪酸是一类重要的生物活性物质,不仅是生物体贮藏能量的主要来源,也是细胞膜的重要组成成分,在生命有机体内具有特殊的生物学功能。其中,多不饱和脂肪酸是细胞膜磷脂的重要组分,能够保持细胞膜的流动性和相对变形性[14],对于维护细胞膜的结构和功能有着重要的作用[15];在叶绿体膜中的多不饱和脂肪酸含量影响膜脂的流动性并决定植物适应温度的能力[16-17]。
本研究拟对草地早熟禾夏季、尤其是秋季低温下叶片脂肪酸的种类和含量进行测定,了解随季节交替气温变化,低温对膜脂脂肪酸种类及饱和水平的影响,探索草地早熟禾的耐冷机制,充分利用草坪草的耐冷性,为延长北方草坪的坪用时间或改善坪用质量提供理论依据。
1 材料与方法
1.1 供试材料与处理
供试地兰州市地处黄河上游,黄土高原西部。由于深居西北内陆,海洋温湿气流不易到达,成雨机会少,大部分地区气候干燥,属大陆性很强的温带季风气候。春夏界线不分明,夏季短促,秋季降温快,冬季漫长,年平均气温10.3℃。夏无酷暑,冬无严寒,是著名的避暑胜地。
供试材料为草地早熟禾午夜,(Poapretensis‘Midnight’)。2012年播种于甘肃农业大学校园高尔夫实训基地,生长期4年。测试样品采集于实训基地休闲区旁的无踩踏非击球区的绿化护坡草坪草。清晨喷灌浇水,夏季隔天1次,入秋后每周2~4次;每月剪草1次。选取颜色、生长密度、修剪高度基本一致的草坪草块分别于2016年7月15日、9月1日、10月1日和10月15日下午4:00—5:00采集草坪草叶片10 g左右,随机选 3个地点重复采样。将采集的样品冰盒保存,快速带至实验室,天平称重每份2 g,用锡铂纸包好,迅速投入到液氮中速冻,于-70℃低温箱贮存待测。于2016年7—11月在甘肃农业大学,甘肃省干旱生境作物学重点实验室对样品进行测定。
1.2 脂肪酸提取及甲酯化
将2 g样品加液氮研磨至粉末状,加入14%的三氯化硼-甲醇溶液10 mL,充分混合,90℃水浴回流90 min;冰浴冷却后,转移至50 mL离心管中,加入9 mL正己烷,振荡混匀;室温下4 000 rpm离心10 min,收集正己烷相;正己烷重复萃取2次;合并正己烷相,40℃减压浓缩至近干;加入0.6 mL甲醇,超声波处理,使残渣全部溶解,过0.22 μm有机滤膜,进GC-MS仪器分析。
1.3 气相色谱-质谱条件
脂肪酸的测定使用Agilent 6890 GC-5973 MSD气相色谱-质谱联用仪进行检测。
色谱条件:色谱柱HP-88(60 m×0.25 mm×0.25 m);进样量为1 uL;进样口温度260℃;分流进样,分流比20∶1;柱温箱升温程序:140℃ 保持 4 min,以3℃·min-1上升到 230℃,保持10 min。载气(高纯氦)流速 1.1 mL·min-1。
质谱条件:四级杆温度150℃,离子源温度230℃,电子轰击能70 eV,扫描范围 15~550 m·z-1,辅助温度230℃。
1.4 脂肪酸不饱和指数和不饱和度
将文库中检索到的所有脂肪酸计算膜脂脂肪酸不饱和指数(IUFA),公式为:
式中,Si:膜脂不饱和脂肪酸相对含量;ti:该不饱和脂肪酸所含不饱和键的数量。
膜脂脂肪酸不饱和度=不饱和脂肪酸相对含量(UFA)/ 饱和脂肪酸相对含量(FA)
1.5 数据分析
定性分析:化合物经计算机检索同时与标准品和 NIST Library相匹配,仅报道匹配度和纯度大于800(最大值1 000)的鉴定结果。
定量分析:用气相色谱-质谱(GC-MS)联用技术分析其脂肪酸组成,并用面积归一化法测定了各种成分的质量分数。
采用IBM SPSS Statistics 21统计分析软件进行单因素方差分析。
2 结果与分析
2.1 草地早熟禾总脂肪酸种类分析
对草地早熟禾叶片脂肪酸进行提取和甲酯化样品前处理,利用GC-MS进行定性和定量分析,总离子流色谱图如图1所示。利用NIST97.L质谱库自动检索,共检测出27种脂肪酸,其中13种为饱和脂肪酸,另外14种为不饱和脂肪酸。如表1所示,饱和脂肪酸主要有棕榈酸、硬脂酸、苯甲酸、月桂酸、肉豆蔻酸、癸酸、丁二酸、十五烷酸、十七碳酸、廿碳酸、二十二碳酸、二十三碳酸等。不饱和脂肪酸中,除丁烯二酸、棕榈酸(7-;9-)和油酸4种单不饱和脂肪酸外,其他10种均为多不饱和脂肪酸,尤以亚油酸、亚麻酸、廿碳二烯酸、廿碳三烯酸、廿碳四烯酸、二十二碳四烯酸等18碳、廿碳和二十二碳多不饱和脂肪酸为主。

图1 草地早熟禾脂肪酸甲酯总离子流色谱图Fig.1 The total ion current chromatogram for fatty acid methyl ester in Poa pratensis L.

表1 草地早熟禾主要脂肪酸出峰时间与种类Table 1 Retention time and species of main fatty acid in Poa pratensis L.

峰编号Peak No.出峰时间Retention times/min化合物Compounds英文名English name中文名Chinese name化学式Formula2420.966,9,12-Octadecatrienoic acid, methyl ester6,9,12-亚麻酸甲酯(C18:3)C19H32O22118.418,11-Eicosadienoic acid, methyl ester8,11-廿碳二烯酸甲酯(C20:2)C21H38O22320.738,11,14-Eicosatrienoic acid, methyl ester, (Z,Z,Z)-8,11,14-廿碳三烯酸甲酯(C20:3)C21H36O21817.0411,14,17-Eicosatrienoic acid, methyl ester11,14,17-廿碳三烯酸甲酯(C20:3)C21H36O22521.11Methyl (Z)-5,11,14,17-eicosatetraenoate5,11,14,17-廿碳四烯酸甲酯(C20:4)C21H34O22621.256,9,12,15-Docosatetraenoic acid, methyl ester6,9,12,15-二十二碳四烯酸(C22:4)C23H38O269.5413-Tetradecynoic acid, methyl ester十四炔酸甲酯C15H26O2
2.2 草地早熟禾夏、秋季脂肪酸成分和含量分析
据天气网公布的数据[18-21],兰州市夏季日均温14~27℃,秋季是11~23℃。根据这一气候特点及历年的气温数据,实验设计于7月15日、9月1日、10月1日和10月15日,对兰州市区甘肃农业大学校园高尔夫实训基地生长4年的午夜草地早熟禾,采样进行脂肪酸检测。这4个采样时间最高温分别为32℃、28℃、27℃和22℃,最低温为18℃、15℃、12℃和8℃,日温差为13~15℃,能够代表西北黄土高原盛夏、秋季气温变化特点。

图2 兰州市2016年7月至10月气温图Fig.2 The temperature figure during July to October in 2016 in Lanzhou city
对7月15日、9月1日、10月1日和10月15日采集的草地早熟禾样品进行脂肪酸检测。如表2所示,草地早熟禾中饱和脂肪酸约占脂肪酸质量总数的17%,以棕榈酸(约占14%)和硬脂酸(约占1.4%)为主,而苯甲酸、月桂酸、肉豆蔻酸、癸酸、丁二酸、十五烷酸、十七碳酸、廿碳酸、二十二碳酸、二十三碳酸等饱和脂肪酸总质量不超过2%;不饱和脂肪酸约占83%,以亚麻酸(约占61.7%)、亚油酸(约占13.5%)、油酸(约占4.2%)和棕榈油酸(约占2.3%)为主。草地早熟禾叶片内稳定存在的主要有肉豆蔻酸、十五烷酸、棕榈酸、棕榈油酸、十七烷酸、硬脂酸、油酸、亚油酸、亚麻酸、廿碳三烯酸、二十二碳烷酸、二十碳四烯酸,共计13种脂肪酸,约占98.206%~99.196%;在夏季出现反丁烯二酸和十四炔酸2种独特的脂肪酸;进入秋季以后,随着气温逐渐转凉,叶片内又产生了苯甲酸、月桂酸、癸酸、丁二酸、廿碳烷酸、庚二烯、二十三碳烷酸、二十二碳四烯酸,及不同双键位的棕榈油酸、亚麻酸和廿碳三烯酸等,共计12种,在质膜中含量微小,仅有0.804%~1.794%。
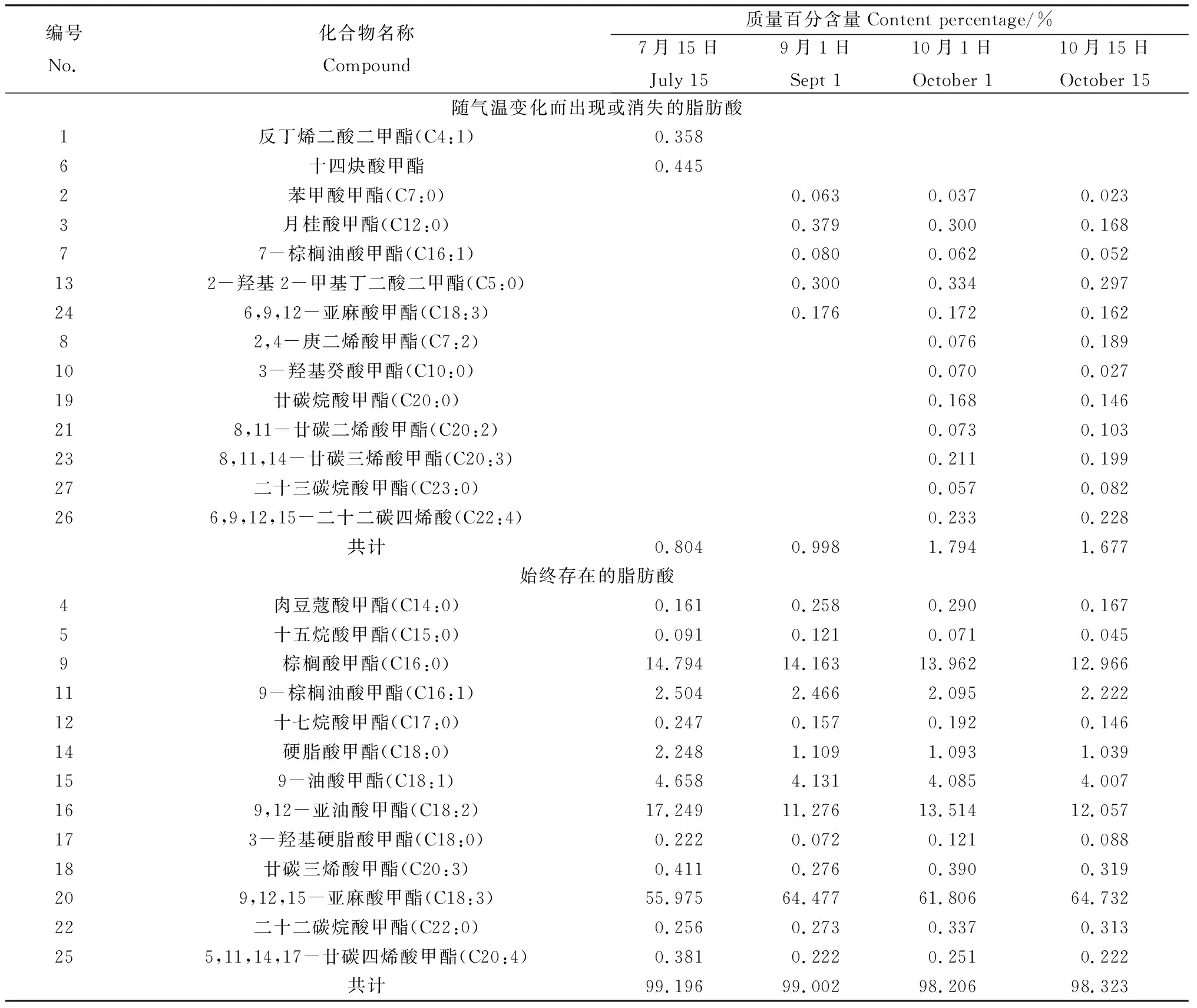
表2 草地早熟禾叶片夏、秋季脂肪酸含量变化Table 2 The changes of fatty acid content in leaf of Poa pratensis L. during summer and autumn
对夏季7月15日采集的样品和入秋后3个时间段采集的样品脂肪酸结果进行归纳总结和数据分析发现,随气温变化,不仅脂肪酸的种类发生着改变,而且各种脂肪酸的百分含量也发生了变化。饱和脂肪酸与单烯不饱和脂肪酸夏、秋季交替,均随温度的逐渐降低,在膜质中所占质量百分含量也明显降低;但入秋以后,无显著变化。随着气温的持续下降,二烯脂肪酸百分含量大幅降低,三烯脂肪酸的含量显著增加(图3)。从而使三烯脂肪酸与二烯脂肪酸的比例发生改变,比值大大增加。在盛夏,二烯脂肪酸含量远高于秋季,三烯脂肪酸含量则相反,凉爽的秋季高于酷热的夏季。

图3 草地早熟禾脂肪酸组分变化Fig.3 The changes of fatty acid composition in Poa pratensis L.注:同一组柱上不同字母表示显著差异(P<0.05)Note: Different letters in the same group of column indicate significant difference at the 0.05 level
2.3 夏、秋季草地早熟禾膜脂脂肪酸不饱和指数与不饱和度分析
对检测到的脂肪酸不饱和指数(IUFA)和不饱和度进行分析。如图4 所示,夏季草地早熟禾IUFA较低为213.14,进入秋季以后,随着气温的逐渐降低,IUFA开始升高,到深秋的10月15日,IUFA上升至229.02,显著高于盛夏;相应的脂肪酸不饱和度也呈上升趋势,由盛夏的4.55增至深秋的5.45。

图4 草地早熟禾膜脂脂肪酸不饱和指数和不饱和度变化Fig. 4 The change of double bond index and unsatuation degree of fatty acid in Poa pratensis L.注:图上不同字母表示显著差异(P<0.05)Note: Different letters indicate significant difference at the 0.05 level
3 讨论
3.1 秋季低温对草地早熟禾脂肪酸种类的影响
脂肪酸是生物体基本组成之一,具重要生理功能。它们既是细胞膜脂的主要成分,又是重要的能源物质,还是一些信号分子的前体。此外,它还与细胞识别、特异性和组织免疫等有密切关系[22]。据报道,含两个以上双键的多不饱和脂肪酸是机体生长发育、构成细胞脂膜必不可少的成分。由于多不饱和脂肪酸中双键位置的不同,其构型则完全不同,功能也不同[14]。
对草地早熟禾叶片质膜脂肪酸进行检测发现,长期稳定存在的主要有棕榈酸、棕榈油酸、硬脂酸、油酸、亚油酸、亚麻酸这6种主要脂肪酸种类以外,还有小比例存在的肉豆蔻酸、十五烷酸、十七烷酸、廿碳三烯酸、二十二碳烷酸、二十碳四烯酸,共计13种脂肪酸,约占总质量的98.2%~99.2%。随着季节交替、气温变化,有含量甚少的脂肪酸种类出现或消逝。在夏季出现反丁烯二酸和十四炔酸2种独特的脂肪酸;反丁烯二酸又称富马酸或延胡索酸,是植物中最简单的不饱和二元羧酸,作为一种短链的挥发性脂肪酸,其存在可能与草坪草特有的芳草味有关。进入秋季以后,随着气温逐渐转凉,叶片内又产生了饱和脂肪酸:苯甲酸、月桂酸、癸酸、丁二酸、廿碳烷酸、二十三碳烷酸,不饱和脂肪酸:庚二烯、二十二碳四烯酸,及不同双键位的棕榈油酸、亚麻酸和廿碳三烯酸等,共计12种,在质膜中含量微小且逐渐增多,由最初的0.998%增长为1.677%。由于这些脂肪酸在植物中相对含量微小和变化性,因而目前无更深入的研究。在植物组织和细胞中, 脂肪酸绝大部分为结合形式,极少数以游离形式存在[22]。结合草地早熟禾夏季生长缓慢、秋季生长旺盛的生长发育特点和秋季凉爽的气候特点,推测入秋后植物体内出现的这12 种脂肪酸可能与其它物质一起,分布于机体表面,防止机械损伤和热量散失等有关[22]。
3.2 气温对脂肪酸不饱和性的影响
脂肪酸分为饱和脂肪酸和不饱和脂肪酸。构成细胞膜磷脂双分子层中的脂肪酸成分及其饱和水平与膜稳定性密切相关。细胞膜的稳定性得益于细胞膜膜质的流动性,其流动性的大小与以下两因素有关:
一方面,在于细胞环境温度的高低。周围温度愈高,脂质双层膜的流动性愈大[23]。反之,在低温下,细胞膜经历了相变,从一个高度流动的液晶相变为一个更为僵硬的凝胶相。在凝胶相时,脂质紧密排列,高度有序,这阻碍了膜的正常生理功能,可能导致膜渗漏,甚至膜破裂[24]。
另一方面,取决于构成脂质双层膜的脂质分子的脂肪酸链的饱和程度的大小。不饱和的程度愈大,流动性愈大[23]。饱和脂肪酸碳原子间缺乏双键,而不饱和脂肪酸在顺式或反式结构中有一个或多个双键,它通过在脂酰链中引入弯曲或蜷缩,从而抑制相邻脂质分子的紧密排列[24-25],增加膜的流动性;另外,脂质的不饱和度和种类能显著地影响膜的相变温度范围[24]。细胞膜磷脂中饱和脂肪酸棕榈酸(C16:0)和硬脂酸(C18:0)的融点温度分别是62.9℃和69.5℃,单不饱和脂肪酸棕榈油酸(C16:1)和油酸(C18:1)分别是-0.1℃和13.4℃,多不饱和脂肪酸亚油酸(C18:2)和亚麻酸(C18:3)分别是-5℃和-10℃[26]。不饱和脂肪酸尤其是多不饱和脂肪酸可降低细胞膜相变温度,被认为有助于保持膜呈流动状态,这对于膜的生物学功能是必需的[27]。夏明[28]等对20种苜蓿在低温下叶片膜脂脂肪酸的组分和变化进行分析,认为亚麻酸和棕榈油酸要比其它不饱和脂肪酸在抗寒力方面具有更大的贡献,也从实验的角度验证了不饱和脂肪酸种类与植物抗寒能力间的关系。
膜质脂肪酸不饱和指数(IUFA)能反映膜的流动性。脂肪酸不饱和度(UFA/FA)可直接反映脂肪酸的构成比例的变化。对草地早熟禾夏、秋季叶片脂肪酸的不饱和性进行研究,数据显示,随着气温的逐渐降低,IUFA开始升高,到深秋的10月15日,IUFA上升至229.02,显著高于盛夏的213.14;相应的脂肪酸不饱和度也呈上升趋势,由盛夏的4.55增至深秋的5.45。说明入秋以后,虽然环境温度降低了质膜的流动性,但脂肪酸不饱和程度大大升高,增加了质膜的流动性,在总体上保证了秋季细胞膜的稳定性,有利于维系低温下膜结构正常的生理功能和避免膜脂固化造成的膜伤害。此结论与刘慧民等[29]对低温与脱锻炼处理下2种绣线菊膜脂脂肪酸变化;孙汉洲等[30]筛选抗寒性巨桉,马程等[31]对油松针叶细胞膜脂肪酸组成的季节性变化;张婷等[32]对哈密瓜果实耐冷性与细胞膜脂肪酸关系的研究;Li等[33]对秋季低下4种菊花叶片和根中膜脂肪酸成份比较的研究结论基本一致:植物叶中不饱和脂肪酸含量随气温降低而逐渐升高,低温有助于不饱和脂肪酸的形成和IUFA的升高。植物叶片中膜脂不饱和脂肪酸含量提高,膜脂相变温度会降低,植物的低温抗性相应提高。因此,不饱和脂肪酸具有防止植物遭受低温损伤的作用[17]。
Falcone和Ogas[34]认为,植物细胞膜随温度的变化,三烯脂肪酸与二烯脂肪酸的比例将发生不同的变化。许多植物的三烯脂肪酸含量随温度的降低而升高,随温度的升高而降低。本研究中,在盛夏二烯脂肪酸的含量远高于秋季;而三烯脂肪酸含量则相反,在凉爽的秋季高于酷热的夏季。此结论与Wang等[35]在转基因水稻和Zhou等[36]在转基因杨树的研究,低温胁迫下三烯脂肪酸含量上升;高温环境中三烯脂肪酸含量降低的结论相近。此外,本研究发现草地早熟禾叶片的饱和脂肪酸与单烯不饱和脂肪酸入秋后,随气温降低,含量无显著变化。与Cyril等[27]对雀稗在低温下2种饱和脂肪酸棕榈酸和硬脂酸含量无变化,而三烯不饱和脂肪酸亚麻酸含量显著增加的结论较一致。结合Yuuki等[37]对编码叶绿体omega-3脂肪酸脱饱和基因(FAD7)的转基因烟草耐高温的研究结果,转基因烟草T15和T23比野生型烟草耐高温的唯一不同之处在于转基因烟草植株的叶绿体中含有较低水平的三烯脂肪酸和较高水平的二烯脂肪酸;以及Sukit等[38]对脂肪酸体外试验的结果,不饱和脂肪酸油酸和亚油酸低毒,且对膜流动性影响较小。这些证据进一步明确表明三烯脂肪酸的积累与植物对温度的耐受性有关。除此以外,三烯脂肪酸对于植物在干旱、盐等逆境条件下生存也具有极其重要的作用[12,32]。
3.3 植物脂肪酸检测方法
脂肪酸在生物体中普遍存在,与人类生活息息相关,多年来一直受到研究者的高度重视。相应地也涌现出大量的脂肪酸检测方法。比较了近年来有关植物脂肪酸的检测方法,大多数只能检测出5~8种主要的脂肪酸种类[29,39-40];在进行定量分析时,多以十七碳脂肪酸作为内标。本研究所采用的植物脂肪酸甲酯化及提取方法和气相色谱-质谱上机分离检测条件,共检出27种脂肪酸,此外,本研究检出脂肪酸碳原子数有四碳、五碳、七碳、十碳、十二碳、十四碳、十五碳、十六碳、十七碳、十八碳、廿碳、二十二碳和二十三碳,因此,以十七碳脂肪酸作为内标对植物脂肪酸进行定量分析是不严谨的。
4 结论
草地早熟禾中饱和脂肪酸约占脂肪酸质量总数的17%,不饱和脂肪酸约占83%,叶片质膜含有较高的不饱和脂肪酸,说明草地早熟禾原本具有较强的耐冷性;秋季随着气温的降低,脂肪酸种类和数量均发生了改变;且组成质膜的脂肪酸不饱和度也发生了改变,二烯脂肪酸含量下降,三烯脂肪酸增加。研究结果表明,草地早熟禾为了抵御秋季冷凉气温,其叶片脂肪酸不饱和度增加,进一步增强了对冷害的抗性。
