3种冷季型草坪草对持续干旱、高温及其互作的生理生态响应
2018-08-23宋娅丽
宋娅丽, *, , ,
(1. 西南林业大学生态与水土保持学院, 云南 昆明 650224; 2. 北京林业大学水土保持学院, 北京 100083)
随着水资源短缺和全球气候变暖,部分区域发生旱灾频率和严重性增加[1-2],全球气温也被预测到21世纪末将上升1.8~4.0oC[3]。干热地区的冷季型草坪草面临着干旱和高温的双重胁迫,两者共同影响着草坪草的生长、发育和生理生态过程,这种自然环境极大地限制了绿地草坪的普及和推广,并严重影响其建坪质量后期产量和效率[4-5]。干旱胁迫下质膜透性增加,细胞内电解质渗透增多,电解液相对电导率升高;同时植物体内可以积累渗透调节物质可溶性糖,冠层同化物向根系分配增多,促进根系生长[6-7]。高温胁迫下植物水分亏缺,光合速率降低,生长缓慢,活性氧代谢失调,膜脂过氧化发生,质膜透性增加,但同时植物通过降低蒸腾和增加内可溶性有机物质的浓度来减缓水分缺失[8-10]。
黑麦草(LoliumperenneL.)、高羊茅(FestucaarundinaceaL.)和早熟禾(PoapratensisL.)为多年生冷季型草坪草,最适温度范围为15.6~23.9℃[11],常作为建坪的先锋草种,适合在我国过渡地带及其冷凉地区生长[12]。已有的关于草坪草抗逆性的研究大多集中于抗旱、抗寒、抗高温、抗病虫害、耐盐碱、耐酸铝性等方面,且主要集中在单一胁迫对草坪草的影响,而缺乏干旱高温互作或复合胁迫对不同冷季型草坪草生理生态的研究资料。研究表明,干旱和高温协同发生比干旱或高温单一胁迫对植物生长构成的危害更大[13],而这方面的研究将有助于全面理解不同逆境胁迫影响冷季型草坪草的内在生理机制。由于夏季持续干旱高温的影响,冷季型草坪草的生长受到了限制,但其伤害机制尚不明确。因此,本试验选择3种典型冷季型草坪草,设置干旱、高温及其互作处理,对其幼苗进行持续胁迫,观测并掌握各品种在胁迫环境中的主要生理生态指标变化,拟揭示3种冷季型草坪草对持续干旱高温环境的生理适应性,并为夏季持续干旱高温区域进行冷季型草坪草的引种和养护管理提供理论依据。
1 材料与方法
1.1 试验材料
供试材料为北京克劳沃公司进口的3种多年生冷季型草坪草:黑麦草-辉煌(LoliumperenneL.)、高羊茅-火凤凰2号(FestucaarundinaceaL.)、早熟禾-雪狼(PoapratensisL.),均为价值较高的优良品系。
1.2 试验设计
试验地位于云南省西南林业大学生态与水土保持学院温室大棚,2016年4月初采用盆栽方法进行播种,种子种植于装有多孔粘土(沙土比为1:1)的PVC管中(直径 10 cm,深 40 cm),灌水后将土压实,放于玻璃温室内进行预培养。温室中采用自然光照,不加温,且材料的培养的处理主要参考Qian和Fu[14]的方法。待种子发芽后每2 d浇一次水,每4 d用Hoagland营养液浇灌。在培养期间,每周修剪植物以保持平均高度控制在10 cm。
播种3个月后将植物转入能够控制环境因子的步入式恒温培养箱中生长。在恒温培养箱中进行2周的适应性培养:温度设定为白天21oC,晚上18oC(12 h光照,12 h黑暗),控制空气相对湿度为70%,光照强度为660 μmol m-2·s-1。2周后将叶片高度统一修剪为10 cm,进行干旱、高温及其互作处理,共4周。各处理分为4个:对照(CK)、干旱处理(D)、高温处理(H)或干旱和高温互作处理(D+H)。CK的温度设置为白天20oC,晚上17oC,植物每2 d浇一次水,浇水量为100%水分蒸散量(ET)(浇水量用称重法获得);D处理温度与CK相同,植物同样每2 d浇一次水,但浇水量减小为50% ET;H处理下的植物温度设定为白天35oC,晚上32oC,浇水量与CK处理相同;D+H处理温度与H处理相同,浇水量与D处理相同。
不同品种及不同处理各3个重复。为减小与避免不同恒温培养箱中环境条件不同对样品的影响,将样品在培养箱内随机放置。每个星期将植物重新放在不同恒温培养箱中。其他条件包括:70%的相对湿度、750 μmol m-2·s-1的光照强度,以及12小时的光周期。按照温室中的培养方式对植物浇水和施肥。胁迫的第0、7、14、21、28天分别测定草坪外观质量(turf quality,TQ)、叶片萎蔫系数(leaf wilting score, LWS)、叶片相对含水量(leaf relative water content, LRWC)、光合速率(photosynthetic rate,Pn)、暗呼吸速率(dark respiration, R)、细胞质膜透性(leaf relative conductivity, LRC);试验结束后测定地上部分干重(shoot biomass, SB)、根系干重(root biomass, RB)、脯氨酸含量(prolin content, Pro)和可溶性糖含量(soluble sugar content, SS),所有指标测定均重复3次。
1.3 试验方法
草坪外观质量基于试验过程中草坪草地上部分生长情况按照1到9的数值来评价,1是密度低、长势差、质地差(完全枯萎和棕色)的草坪,而9是密度高(充满整个PVC管)、长势好、均匀整齐、质地好(水分充足、颜色鲜绿)的草坪[15];叶片萎蔫系数是基于萎蔫状况和叶片颜色按照1~9的数值来评价,1是完全萎蔫、枯黄的草坪,而9是水分状况好、颜色鲜绿的草坪;叶片相对含水量测定采用常规烘干法[16];生物量测定采用常规烘干法[17];光合速率和暗呼吸速率使用便携式红外线分析仪(LI 6400, LI-COR Inc., Lincoln, NE, USA)测定;细胞膜透性采用电导率法(DDS-308,上海精密科学仪器有限公司)[18-19];脯氨酸含量采用酸性茚三酮比色法测定[20];可溶性糖含量采用蒽酮法测定[21]。
1.4 数据分析
相关性分析时各指标均采用不同处理时间相同处理的平均值,采用SPSS 19.0进行分析,并用Microsoft Office Excel 2007绘制数据图。
耐受性评价方法采用隶属函数法[9,22],评价时3个品种的各指标均使用胁迫第28天时的数据。
(1)各指标的隶属函数值
当指标与植物耐受性呈正相关时:
U(Xi)=(Xi-Xmin)/(Xmax-Xmin)
当指标与植物耐受性呈负相关时:
U(Xi)=1-(Xi-Xmin)/(Xmax-Xmin)
(2)平均隶属值
X=∑Xi/n
式中,U(Xi)为各指标的隶属值,U(Xi)越大耐受性越强;Xi为某一指标的测定值;Xmax和Xmin分别为某一指标的最大值和最小值;X为平均隶属值。
2 结果与分析
2.1 持续干旱、高温及其互作对3种冷季型草坪草生长外观的影响
持续干旱、高温及互作处理下,3种冷季型草坪草的外部形态均发生了明显变化,且表现不同,受到影响的包括地上部分均匀度、整齐度以及叶片颜色。CK的3种草坪草均生长正常,叶色鲜绿,草坪外观质量(TQ)均保持在8以上。胁迫第7 d时,3种草坪草TQ在D、H和D+H处理下平均值较CK分别下降了3.7%,13.0%和51.6%;除D处理下的高羊茅在第7 d与CK差异性不显著外,3种草坪草在各处理下均在第7 d开始显著下降(P<0.05)(图1)。从影响程度看,在7 d,14 d和21 d,D+H胁迫对TQ的影响大于D或H单独胁迫,干旱和高温互作效应明显,说明干旱和高温互作对TQ的影响具有叠加效应。当胁迫持续到28 d时,H处理对黑麦草和早熟禾的影响程度与D+H无显著差异,说明随着时间的持续D+H的叠加效应逐渐减弱。28 d时,D+H胁迫下黑麦草、高羊茅、早熟禾与CK相比分别下降了73.8%,75.2%和85.0%,此时早熟禾完全干枯。
各胁迫下3种草坪草的叶片萎蔫系数(LWS)与TQ总体上变化一致,均随胁迫时间的持续而显著下降(图2)。各胁迫处理7 d时,3种草坪草的叶片萎蔫系数均显著下降,且D和D+H胁迫的下降程度显著高于H(P<0.05);胁迫持续到28 d,3种草坪草D、H和D+H胁迫的平均LWS与对照相比下降了73.2%,50.8%和71.7%,D和D+H处理无显著差异,说明H胁迫对LWS影响较小,D起到关键作用。28 d时,D+H胁迫下黑麦草、高羊茅、早熟禾与CK相比分别下降了68.9%,63.3%和83.0%。
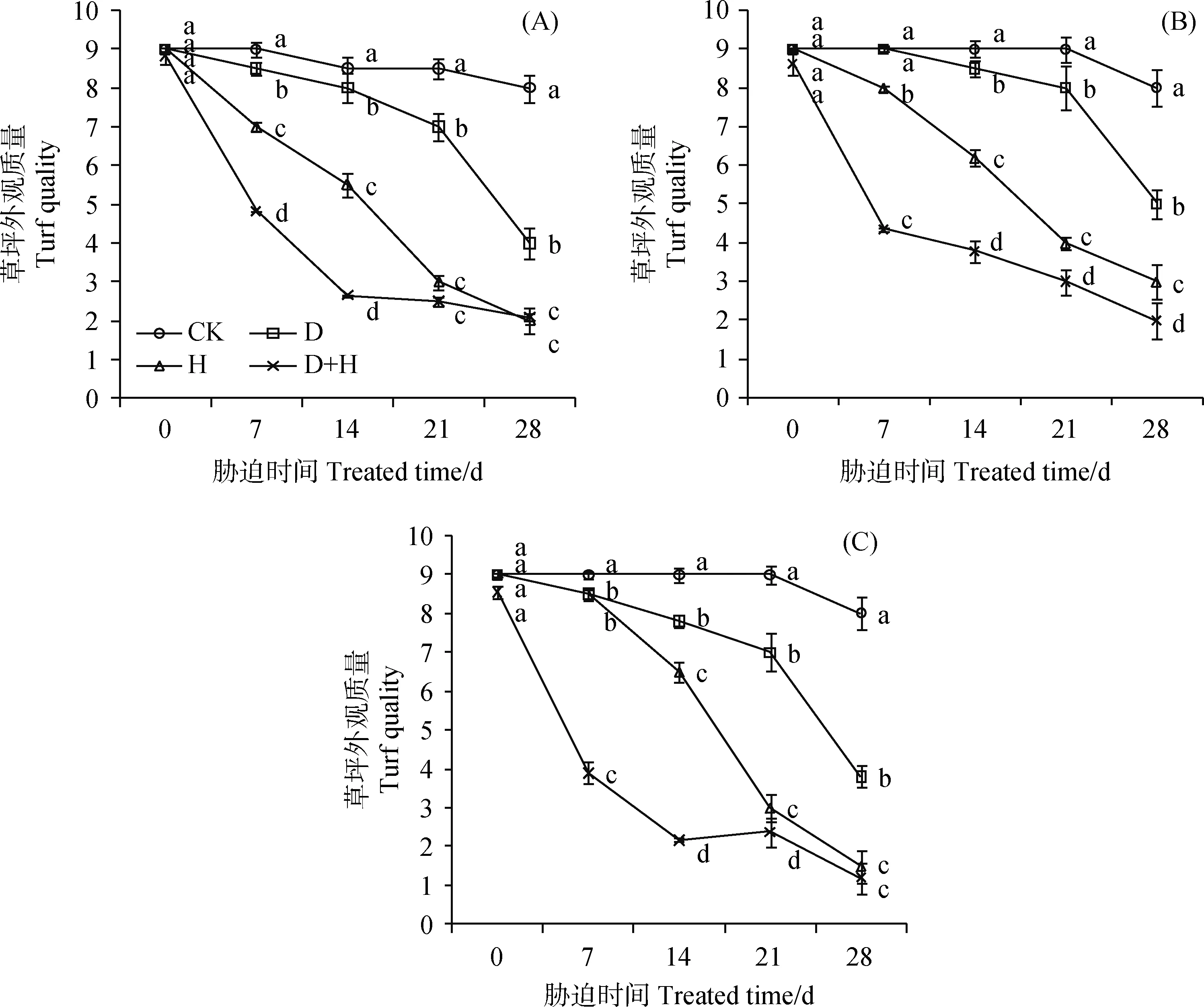
图1 干旱、高温及其互作胁迫对黑麦草(A)、高羊茅(B)和早熟禾(C)草坪外观质量TQ的影响(平均值±标准差)Fig.1 Effect of drought stress, heat stress and their interaction on turf quality of perennial ryegrass (A), tall fescue(B) and kentucky bluegrass (C) (mean±SD)注:图中不同小写字母表示同一处理时间不同胁迫之间差异显著性(P<0.05)Note:Different lower-case letters meant significant difference among different treatments in the same time at the 0.05 level

图2 干旱、高温及其互作胁迫对黑麦草(A)、高羊茅(B)和早熟禾(C)叶片萎蔫系数LWS的影响(平均值±标准差)Fig.2 Effect of drought stress, heat stress and their interaction on leaf wilting score of perennial ryegrass (A), tall fescue (B) and kentucky bluegrass (C) (mean ± SD)
2.2 持续干旱、高温及其互作对3种冷季型草坪草叶片相对含水量的影响
随着胁迫时间的持续,3种草坪草叶片相对含水量呈下降的趋势,胁迫时间越长对叶片相对含水量的影响越严重(图3)。各处理下黑麦草和早熟禾LRWC第28 d均低于50%,比胁迫第21 d分别下降了36.2%和46.9%。各胁迫在第21 d时各处理下LRWC与CK相比均达到显著水平(P<0.05),28 d时呈极显著降低(P<0.01)。胁迫28 d时D、D+H胁迫下的LRWC无显著差异(P<0.05),黑麦草、高羊茅和早熟禾仅为CK的47.4%~56.4%、64.8%~71.3%、31.1%~32.8%;各胁迫下黑麦草、高羊茅和早熟禾的LRWC平均值分别下降44.9%,28.2%和61.7%。这表明逆境胁迫明显导致草坪草叶片水分亏缺,高羊茅叶片持水能力强于黑麦草和早熟禾,早熟禾叶片脱水速度最快。
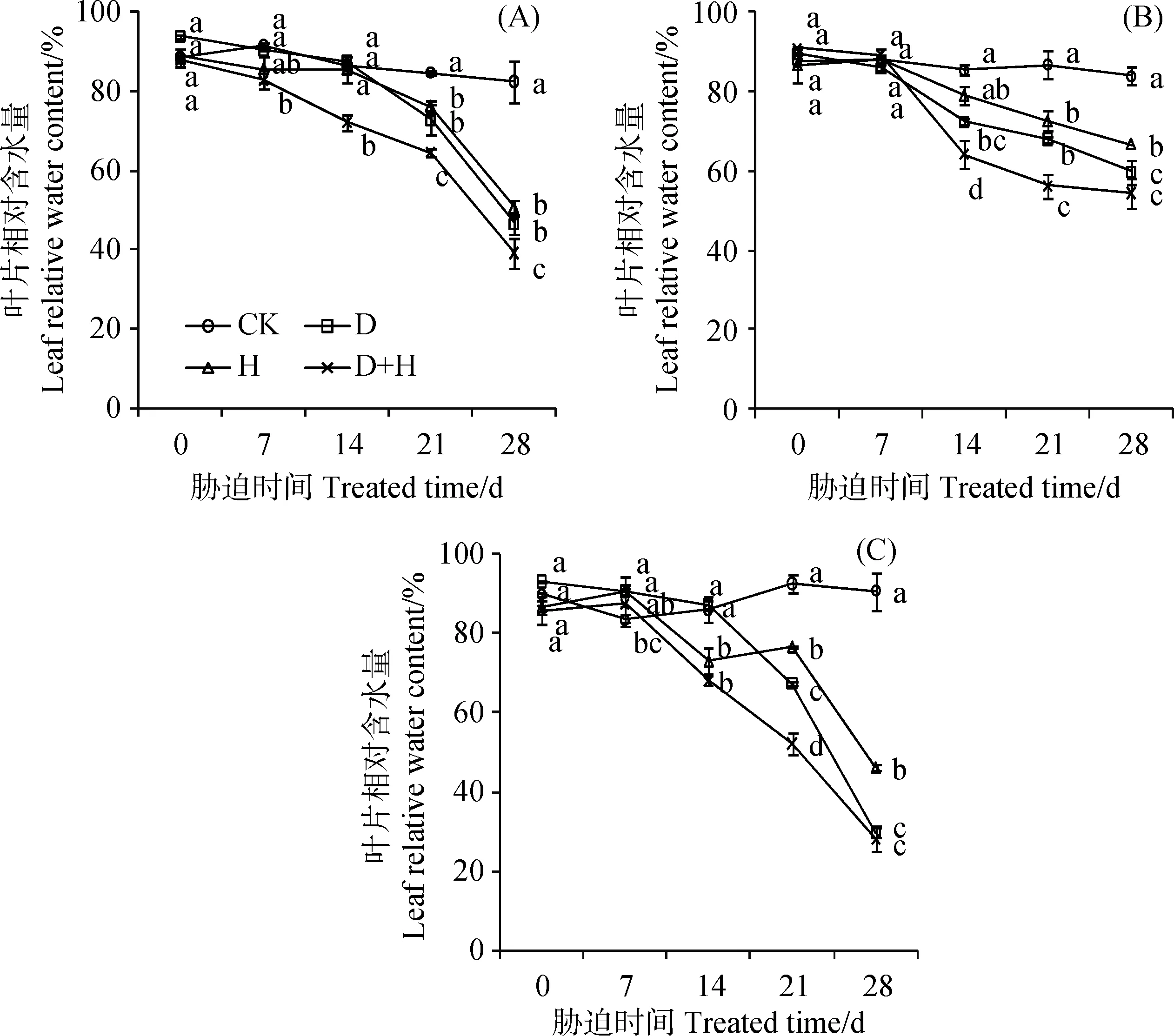
图3 干旱、高温及其互作胁迫对黑麦草(A)、高羊茅(B)和早熟禾(C)叶片相对含水量LRWC的影响(平均值±标准差)Fig.3 Effect of drought stress, heat stress and their interaction on leaf relative water content of perennial ryegrass (A), tall fescue (B) and kentucky bluegrass (C) (mean ± SD)
2.3 持续干旱、高温及其互作对3种冷季型草坪草生物量及根冠比(R/S)的影响
D、H和D+H胁迫均使3种草坪草地上部分生物量(SB)、根系生物量(RB)显著降低(P<0.05)(表1)。地上部分生物量、根系生物量最小值均出现在D+H胁迫下,D、H和D+H胁迫下3种草坪草的地上部分生物量与CK相比,平均下降幅度分别为21.5%,41.0%和50.3%,根系生物量平均下降幅度分别为28.1%,42.2%和55.6%,说明干旱和高温互作对根系生物量的影响大于对地上部分生物量的影响。3种草坪草相比较,在D+H胁迫下早熟禾的下降幅度高于黑麦草和高羊茅,地上部分和根系生物量分别达到80.8%和80.3%,说明干旱和高温互作对早熟禾生长的抑制作用高于黑麦草和高羊茅。各处理对3种草坪草根冠比的影响不尽相同,黑麦草和高羊茅在D+H胁迫下分别下降了16.7%和14.7%,早熟禾提高了9.3%。
2.4 持续干旱、高温及其互作对3种冷季型草坪草叶片光合速率(Pn)和暗呼吸速率(R)的影响
3种冷季型草坪草的叶片光合速率(Pn)均随时间的持续呈显著下降的趋势,且各胁迫处理显著降低了Pn(图4)。黑麦草和高羊茅Pn在胁迫7 d时与0 d有显著差异,早熟禾直到14 d才显著降低(P<0.05)。28 d时各胁迫下Pn与CK相比呈极显著下降趋势(P<0.01),D+H胁迫下Pn几乎接近于0 μmol CO2·m-2·s-1。28 d 时各胁迫下Pn与CK相比下降幅度表现为:D+H>H>D;D+H下黑麦草、高羊茅和早熟禾的Pn分别为CK的8.7%,11.0%和2.5%,下降幅度表现为:早熟禾>黑麦草>高羊茅,这表明早熟禾在干旱高温互作胁迫下叶片光合能力受到严重抑制。

表1 干旱、高温及其互作胁迫对黑麦草、高羊茅和早熟禾生物量和根冠比的影响(平均值±标准差)Table 1 Effect of drought stress, heat stress and their interaction on biomass and root/shoot rate of ryegrass, tall fescue and kentucky bluegrass (mean±SD)
注:同行不同小写字母表示同一测定指标下不同胁迫处理之间差异显著(P<0.05),同列不同大写字母表示同一测定指标下相同胁迫处理不同草坪草间差异显著(P<0.05)
Note:Different lower-case letters meant significant difference among different treatments under same grasses at the 0.05 level, different capital letters meant significant difference among different grasses under same treatment at the 0.05 level
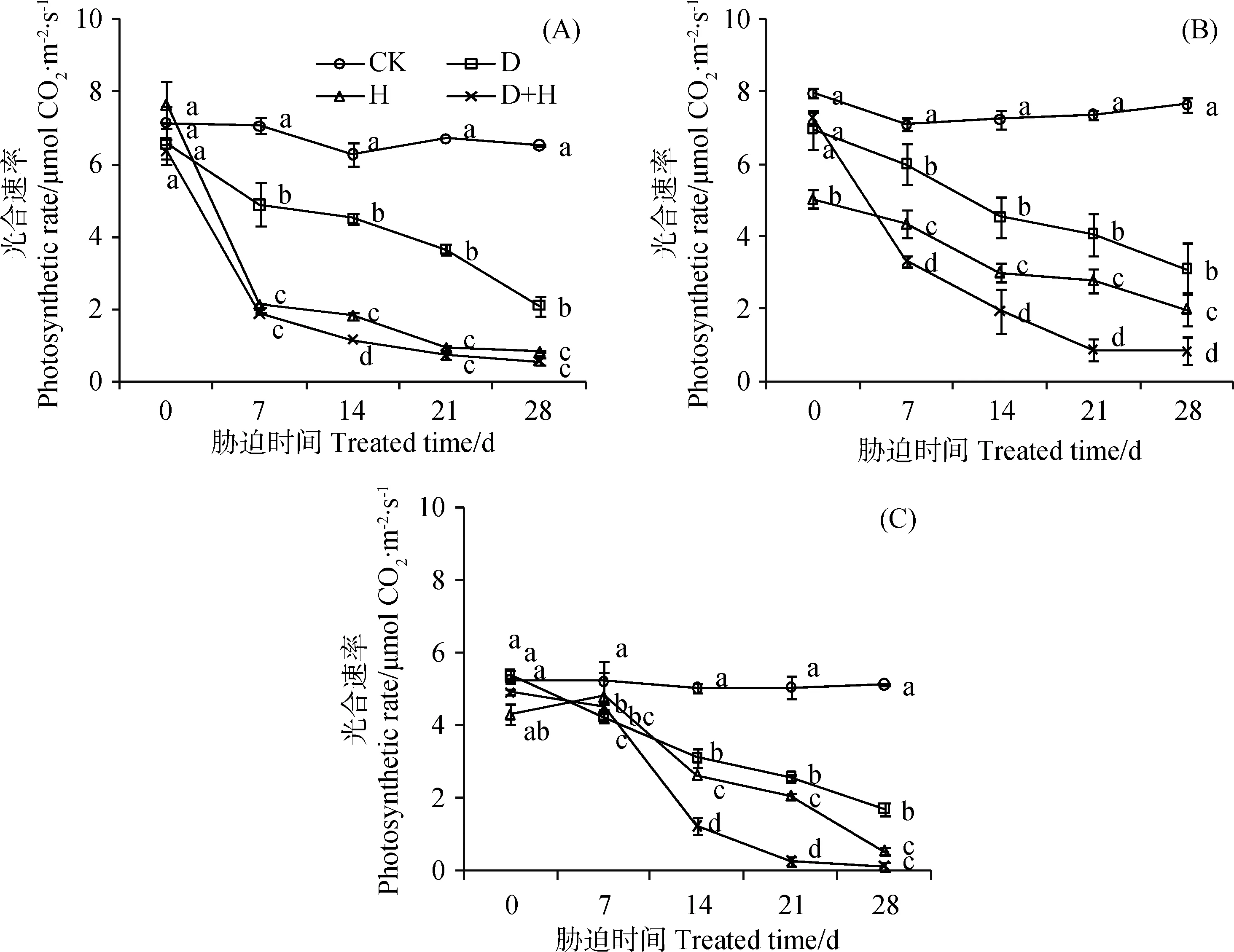
图4 干旱、高温及其互作胁迫对黑麦草(A)、高羊茅(B)和早熟禾(C)叶片光合速率Pn的影响(平均值±标准差)Fig.4 Effect of drought stress, heat stress and their interaction on photosynthetic rate of ryegrass (A), tall fescue (B) and kentucky bluegrass (C) (mean ± SD)
各胁迫下叶片暗呼吸速率(R)与Pn表现相反(图5)。H和D+H胁迫下3种冷季型草坪草的R均随时间的持续呈显著增加的趋势,在28 d时最高,且各胁迫显著增加了R。黑麦草R在H和D+H胁迫7 d时与0 d相比显著升高,高羊茅和早熟禾在7 d无显著差异,直到14 d才显著升高(P<0.05)。28 d 时R与CK相比胁迫增加幅度表现为:D+H>H>D。28 d时H和D+H下的黑麦草、高羊茅和早熟禾R分别比CK增加了137.5%和258.9%,132.1%和169.1%,361.6%和446.9%,升高幅度中早熟禾最高,这表明早熟禾在干旱高温互作胁迫下叶片耐受性和碳代谢状态受到严重抑制。
2.5 持续干旱、高温及其互作对3种冷季型草坪草细胞质膜透性的影响
叶片相对电导率是反映植物膜系统状况的一个重要生理生态指标,植物细胞膜是控制小分子物质进出细胞的屏障,植物在受到逆境胁迫时细胞膜蛋白受损伤,导致胞质的胞液外渗而使相对电导率增大(LRC)。持续干旱、高温及其互作胁迫下黑麦草和高羊茅的LRC变化趋势基本相同(图6),均是在胁迫初期直至胁迫14 d无显著变化,21 ~ 28 d时显著上升(P<0.05);而早熟禾随着胁迫时间的持续,LRC变化不明显。D+H胁迫下,28 d时3种草坪草LRC均显著高于CK。

图5 干旱、高温及其互作胁迫对黑麦草(A)、高羊茅(B)和早熟禾(C)叶片暗呼吸速率R的影响(平均值±标准差)Fig.5 Effect of drought stress, heat stress and their interaction on dark respiration rate of ryegrass (A), tall fescue (B) and kentucky bluegrass (C) (mean ± SD)

图6 干旱、高温及其互作胁迫对黑麦草(A)、高羊茅(B)和早熟禾(C)叶片相对电导率LRC的影响(平均值±标准差)Fig.6 Effect of drought stress, heat stress and their interaction on relative conductivity of ryegrass (A), tall fescue (B) and kentucky bluegrass (C) (mean ± SD)
2.6 持续干旱、高温及其互作对3种冷季型草坪草渗透调节物质的影响
脯氨酸(Pro)是反映植物抗逆性的重要生理生态指标,植物在正常条件下,Pro含量很低;但植物在受干旱高温等逆境胁迫时,Pro会大量积累以进行渗透调节,其积累指数可在一定程度上反映植物受逆境伤害的程度;植物体内Pro积累量越多,则抗逆能力越强。3种冷季型草坪草在受到干旱、高温及其互作胁迫时,体内Pro含量均有不同程度的增加;在H和D+H下呈极显著增加(P<0.01)(图7)。胁迫28 d时,D胁迫下的早熟禾根系Pro与CK相比无显著变化;黑麦草和高羊茅与CK差异显著(P<0.05),叶片和根系平均值分别增加了104.3%和312.9%,说明高羊茅在干旱胁迫下渗透调节能力优于黑麦草和早熟禾。H胁迫下,黑麦草、高羊茅和早熟禾Pro与CK相比增幅分别为268.3%,118.1%和315.1%;D+H胁迫下Pro分别增加362.8%,300.4%和832.9%。D、H和D+H胁迫下根系脯氨酸与CK相比增幅显著高于叶片,平均高出86.0%,39.2%和60.9%,说明在干旱胁迫下根系所能发挥的渗透调节能力高于高温及两者互作条件下。

图7 干旱、高温及其互作胁迫对3种冷季型草坪草叶片脯氨酸含量(A)和根系脯氨酸含量(B)的影响(平均值±标准差)Fig.7 Effect of drought stress, heat stress and their interaction on leaf proline content (A) and root proline content (B) of three kinds of cool season turfgrasses (mean ± SD)
可溶性糖(SS)是植物生长发育的重要渗透调节物质,是植物胁迫诱导的小分子溶质之一,随胁迫程度增加而增加,有利于维持植物体细胞膨压,能减缓由胁迫造成的植物生理代谢不平衡问题。在胁迫28 d植物收获后,3种草坪草各处理下的SS与CK相比均有不同程度的上升(图8)。在D、H和D+H胁迫下,黑麦草叶片和根系平均SS与CK相比分别增加了35.8%,33.1%和46.6%;高羊茅分别增加了54.5%,10.0%和40.8%;早熟禾分别增加了50.6%,53.9%和65.2%。高羊茅在D胁迫下SS上升幅度最高,黑麦草和早熟禾上升幅度最高的表现在D+H互作下,说明高羊茅在H胁迫下渗透调节性能较差。

图8 干旱、高温及其互作胁迫对3种冷季型草坪草叶片可溶性糖含量(A)和根系可溶性糖含量(B)的影响(平均值±标准差)Fig.8 Effect of drought stress, heat stress and their interaction on leaf soluble sugar content (A) and root soluble sugar content (B) of three kinds of cool season turfgrasses (mean ± SD)
2.7 持续干旱、高温及其互作对3种冷季型草坪草各生理生态指标的相关性分析
对干旱、高温及其互作胁迫下3种冷季型草坪草各生理生态指标进行相关性分析(表2),以求反映3种冷季型草坪草在干旱、高温及其互作胁迫下的生理生态指标变化情况。结果表明,TQ与LWS、LRWC、Pn呈极显著正相关;与R、L-Pro、R-Pro、LRC呈极显著负相关;与R/S、R-SS分别呈显著正负相关。LWS与LRWC、Pn呈极显著正相关;与R、LRC呈极显著负相关;与R/S呈显著正相关;与L-Pro、R-Pro呈显著负相关。RWC与Pn呈极显著正相关;与R、L-Pro、R-Pro、LRC、R-SS呈极显著负相关。Pn与R、LRC呈极显著负相关;与L-Pro、R-Pro呈显著负相关;R与L-Pro、LRC呈极显著正相关;与R-Pro呈显著正相关。说明持续干旱、高温及其互作使叶片缺水、光合作用升高、呼吸作用降低的同时,对Pro含量含量的增加有一定的诱导作用。

表2 干旱、高温及其互作胁迫下3种冷季型草坪草个生理生态指标相关性分析Table 2 Correlations between physiological and ecological indices in three kinds of cool season turfgrasses under drought stress, heat stress and their interaction
注:TQ:草坪外观质量;LWS:叶片萎蔫系数;LRWC:叶片相对含水量;SB:地上部分干重;RB:根系干重;R/S:根冠比;Pn:光合速率;R:暗呼吸速率;LRC:叶片相对电导率;L-Pro:叶片脯氨酸;R-Pro:根系脯氨酸; L-SS:叶片可溶性糖;R-SS:根系可溶性糖;*表示α=0.05水平下相关性达到显著;**表示α=0.01水平下相关性达到显著;下同
Note: TQ: turf quality; LWS: leaf wilting score; LRWC: leaf relative water content; SB: shoot biomass; RB: root biomass; R/S: root to shoot; Pn: photosynthetic rate;R: dark respiration; LRC: leaf relative conductivity; L-Pro: leaf proline content; R-Pro: root proline content; L-SS: leaf soluble sugar; R-SS: root soluble sugar. *indicates significant correlation at the level of alpha=0.05); **indicates significant correlation at the level of alpha =0.01; the same as below
2.8 持续干旱、高温及其互作对3种冷季型草坪草的耐受性评价
利用模糊数学的隶属函数法对3种冷季型草坪草的9项生理生态指标进行耐受性的综合评价(表3),用每种草坪草各项指标隶属度的平均值作为不同草坪草抗逆性综合鉴定标准,该值越大抗逆性则越强。结果表明,黑麦草、高羊茅和早熟禾在D、H、D+H胁迫下的隶属函数平均值分别为0.521,0.827和0.224,表明3种草坪草抗干旱、抗高温、抗干旱与高温互作能力顺序均为高羊茅>黑麦草>早熟禾。
3 讨论
干旱和高温是两个主要限制冷季型植物生长的环境因子,持续的干旱高温胁迫会通过影响生理过程而抑制植物的正常生长发育[23]。这种抑制作用最初体现在植物叶片外观质量变化上。本研究发现早熟禾受持续干旱高温胁迫的伤害最大,在处理28 d时整株叶片枯黄,比黑麦草和早熟禾草坪外观质量值低42.9%和40.0%。从叶片萎蔫系数可知,干旱高温对高羊茅的伤害同样轻于黑麦草和早熟禾,其通过调节自身的生理机制做出积极且持久的抗逆反应,耐受性最强;黑麦草介于高羊茅和早熟禾之间。14 d后,D、H及D+H胁迫下,3种草坪草的叶片相对含水量显著下降,下降的幅度随着时间的持续而逐渐增大,平均值从高到低依次为高羊茅>黑麦草>早熟禾,高羊茅的叶片相对含水量显著高于其他草坪草,这与叶片外观质量和叶片萎蔫系数表现一致。叶片水分相对含水量在28 d时迅速降低,说明随着胁迫时间的持续,蒸腾速率增大,草坪草体内水分亏缺加剧。生物量在根系和地上部分之间的分配对于植物生长是决定性因素,这种分配作用在胁迫下更明显[24-25]。逆境胁迫下,3种草坪草根系生物量的下降幅度高于地上部分,说明干旱和高温对根系生长的影响比地上部分更敏感。这个结果同样出现在其他种类的草坪上[26]。从生物量可知,干旱和高温胁迫对早熟禾生长的抑制作用高于黑麦草和高羊茅。

表3 三种冷季型草坪草对干旱、高温及其互作胁迫的耐受性综合评价Table 3 Comprehensive evaluation on drought stress, heat stress and their interaction of three kinds of cool season turfgrasses
研究表明,植物受到一定程度的干旱胁迫时,植物的水分利用效率将提高[27]。本研究中,当干旱胁迫持续到第7 d,叶片相对含水率并没有显著降低,这可能是由于干旱胁迫下草坪草由气孔导度减小而导致降低的光合速率速度小于或滞后于蒸腾速率的下降速度,导致叶片水分利用效率增加。也有研究表明,叶片相对含水量小于50%时,光合机构的正常运转受到影响[28]。本试验中,28 d时干旱和高温互作胁迫下黑麦草和早熟禾的LRWC均小于50%,光合速率趋向于0,低于干旱或高温单一胁迫下。3种草坪草的光合作用对干旱和高温互作胁迫下的响应强于单一胁迫下。此时草坪草通过积累大量脯氨酸和可溶性糖(比CK高207.0%~1 102.5%)来防止植物水分散失,主动降低渗透势提高渗透调节能力,降低质膜热受害程度,以保证草坪草在逆境下的生存。
干旱会伴随高温同时发生,并加剧高温对植物的光合影响,光合作用是对高温反应最敏感的生理过程之一[29]。当温度超过植物自身调节的控制范围后导致植物出现热害症状,甚至萎蔫死亡[30]。本研究中在高温胁迫导致3种草坪草的光合速率显著降低,造成光合速率大大下降的原因可能是干旱高温导致水分散平衡细胞质与液泡间的渗透势失和植物细胞脱水,光合色素含量急剧下降,造成光合机构的伤害,气孔导度降低使叶绿体内CO2的供应受阻[8]。同时,本研究中耐热性强的高羊茅光合速率下降幅度小于耐热性弱的黑麦草和早熟禾,这也体现在其他植物中,如新疆榛(Corylusheterophylla×C.avellan)[31]、玉米(Zeamays)[32]和金荞麦(Fogopyrumdibotrys)[33]等,结论与本研究结果一致。但值得注意的是,本研究中14 d时在各处理下草坪草并未死亡,说明此时气孔并未完全关闭,相反必须保持一定的开度,使强烈的蒸腾带走过多的热量,以适应逆境。干旱和高温互作胁迫会抑制冷季型草坪草的光合作用,加剧光合速率降低,但在此实验中耐干旱高温的高羊茅比不耐干旱高温的草种具有更高的Pn,这可能与其叶片内具有较高的抗氧化酶活性和叶绿素合成速率有关[34-35]。这些生理代谢的变化有助于减缓逆境对其光合系统的破坏。同时,其自身又通过各种生理机理的变化以增强自己对极端干旱和高温环境的适应。
持续干旱、高温导致3种草坪草通过增加呼吸速率来适应环境的改变,呼吸作用的增强消耗了大量的糖类等能源物质,在28 d时平均增加132.1%~446.9%,在干旱高温互作胁迫下增加高于单一胁迫,说明干旱高温互作下植物光合产量仍在进一步减少,最终导致CO2吸收的终止,比单一胁迫对植物造成的伤害更为严重。干旱和高温互作胁迫下高羊茅的呼吸速率显著低于黑麦草(15.2%)和早熟禾(30.5%),说明逆境生长条件下的高羊茅具有较好的干旱高温耐受性和碳代谢状态。细胞膜是防止细胞外物质自由进入细胞的屏障,它能保证细胞内环境的相对稳定,使各种生化反应能够有序运行。植物在受到逆境胁迫时,质膜透性增大已被许多研究证实[9,33,36]。但本研究中干旱高温互作胁迫下的质膜透性增大幅度并不显著高于干旱或高温单一胁迫。这可能是由于单一胁迫已导致膜结构失去活性,膜对细胞内外物质交换和控制能力消失,外渗液电导趋近死亡电导率[37]。逆境胁迫会抑制冷季型草坪草的光合作用,提高呼吸作用,但较为耐热的草种比不耐热的草种具有更高的光合速率,和更低的呼吸速率。这些生理代谢的变化有助于缓解高温对其光合系统的破坏[38]。
脯氨酸和可溶性糖是植物体内的主要渗透调节物质,两者均能通过降低细胞原生质的渗透势来减轻渗透胁迫,促使细胞从外界吸收水分,使植物保持一定的含水量和膨压,防止膜质和蛋白质的过氧化[39]。本试验中,3种草坪草叶片和根系脯氨酸含量均在各胁迫下显著增加,这与李力等[9]的研究结果一致。研究表明,植物体内脯氨酸的积累程度与植物的抗逆性强弱有一定关联[34,40],本试验在胁迫28 d时,D+H下早熟禾叶片和根系Pro的含量最高,Pro积累程度为早熟禾>黑麦草>高羊茅,结果与草坪外观质量和叶片萎蔫系数变化一致,表明脯氨酸是草坪草应对干旱高温胁迫的主导性渗透调节物质,这与史燕山等[41]和韩春梅[42]的研究结果一致。在受到逆境胁迫时,脯氨酸可通过最有效的渗透作用来调节细胞水分环境的变化,抵抗外界渗透胁迫,从而减轻干旱高温胁迫对植物体内水分亏缺的影响。持续干旱高温胁迫可诱导3种冷季型草坪草体内大量形成可溶性糖,以提高植物的抗逆性。本试验中,3种草坪草的可溶性糖含量在各胁迫处理下均有不同程度的上升,早熟禾的可溶性糖含量变化幅度均比黑麦草和高羊茅高,且对干旱高温互作处理表现的更为敏感,而黑麦草和高羊茅的敏感程度差异较小,因此早熟禾对干旱高温的响应速度最快。较高的可溶性糖含量不仅可以提高细胞的持水力,提高其渗透调节作用,还能增强原生质弹性,有效的保护细胞的膜结构,保证植株的正常生长[43]。
干旱高温胁迫对植物的影响是多方面的,植物也可以通过自身营养物质、光合呼吸速率、渗透调节物质等多种生理机制的共同作用来减轻或减缓伤害的发生。干旱高温胁迫下,3种冷季型草坪草通过各种生理生态反应以低于逆境,维持正常生长,各种抗干旱高温指标的变化趋势总体相同,不同胁迫处理和不同品种各指标的变化规律存在差异,3个品种的耐干旱高温能力也存在一定区别。本试验中,草坪外观质量、叶片萎蔫系数、叶片相对含水量、根冠比、光合速率、暗呼吸速率、质膜透性、脯氨酸含量和可溶性糖含量对干旱高温胁迫处理比较敏感,故可作为冷季型草坪草抗逆性生理鉴定指标。3种冷季型草坪草在干旱高温胁迫下各生理指标的变化情况比较复杂,但根据整个胁迫中植株的生长状态,并通过隶属函数法综合分析各种草坪草的耐干旱高温性能,综合分析得出3种冷季型草坪草抗干旱高温能力由强到弱依次为:高羊茅>黑麦草>早熟禾。3种草坪草在干旱高温条件下通过增加细胞渗透调节物质含量,降低水分丧失,平衡细胞质与液泡问的渗透势,来增强其对干旱高温环境的适应能力,干旱高温共同胁迫对草坪草的伤害高于单一胁迫。
4 结论
3种草坪草的草坪外观质量、叶片萎蔫系数和叶片相对含水量在持续干旱、高温及干旱高温互作胁迫下显著下降,下降的幅度随着时间的持续而逐渐增大,平均值从高到低依次为高羊茅>黑麦草>早熟禾;在干旱、高温及干旱高温互作胁迫下草坪草根系生物量的下降幅度高于地上部分,逆境对根系生长的影响比地上部分更敏感。
持续干旱、高温及干旱高温互作胁迫导致3种草坪草的光合速率显著降低,两者互作下的降低幅度高于单一胁迫下,耐热性强的高羊茅光合速率下降幅度小于耐热性弱的黑麦草和早熟禾;相反,干旱、高温及干旱高温互作胁迫导致3种草坪草暗呼吸速率增加,且在干旱高温互作胁迫下增加高于单一胁迫,说明干旱高温互作比单一胁迫对植物造成的伤害更为严重。
3种草坪草叶片和根系脯氨酸和可溶性糖含量均在高温、以及干旱高温互作下显著增加,早熟禾的变化幅度均高于黑麦草和高羊茅,且对干旱高温互作处理表现的更为敏感,而黑麦草和高羊茅的敏感程度差异较小,因此早熟禾对干旱高温的响应速度最快。
3种冷季型草坪草在干旱高温胁迫下各生理指标的变化情况比较复杂,但根据整个胁迫中植株的生长状态,并通过隶属函数法综合分析各种草坪草的耐干旱高温性能,综合分析得出3种冷季型草坪草抗干旱高温能力由强到弱依次为:高羊茅>黑麦草>早熟禾。
