基于生长模型的速生阔叶树种林分碳汇潜力研究
2018-08-10朱丽艳吴雪琼
吴 恒,朱丽艳,吴雪琼
(国家林业局昆明勘察设计院,云南 昆明 650216)
森林是陆地生态系统最重要的碳库,在全球气候变暖的背景下森林碳汇在森林生态效益中的作用日益凸显[1]。速生阔叶树种由于其速生性和丰产性在森林碳汇中发挥着重要的作用,发展速生阔叶树种碳汇贸易对改善生态和发展经济具有积极的意义。森林碳储量估计方法是评价森林碳汇生态效益的基础,现今国内外普遍运用的碳汇计量方法有生物量法[2]、蓄积量法[3]、生物量清单法[4]、涡旋相关法等[5]。以生长模型[6-7]或生长过程[8]为基础的碳汇潜力分析可作为森林经营管理的决策工具[9]。但长期以来速生阔叶树种生长模型研究的缺失,限制了速生阔叶树种在碳汇经营中的效用。
桤木、银荆、蓝桉和直干桉等是我国亚热带地区主要的退耕还林[10-11]和碳汇造林[12]阔叶树种。对桤木[13]、银荆[14]、蓝桉和直干桉[15]林分生物量的研究缺少了不同年龄阶段的生物量变化动态分析,且未进行林分碳储量的比较。立地质量和林分密度是决定森林生产力的重要因素,缺少在相同立地条件和林分密度条件下林分生物量和碳储量的生产力比较,不能全面反映不同树种的生产力和碳汇潜力。沈彪等[16]对秦岭中段南坡锐齿栎林碳密度的研究表明,地形因子与森林碳密度关系密切,地形因子作为立地质量评价的重要指标影响着林分的生长,进而影响林分碳储量的生长过程。不同立地条件下相同树种的林分生产力存在差异,在相同立地条件不同林分密度下林分生长力也存在差异。对林分生长过程中碳储量进行量化分析,能为速生阔叶树种碳汇潜力评价提供依据[17]。
碳汇经营[18-19]是森林多功能经营的目标之一,适宜的树种选择是碳汇经营的前提[20],比较不同速生阔叶树种各发育阶段碳汇潜力能为碳汇造林提供依据[21]。本研究将林分断面积生长模型和单木生物量模型与器官含碳率链接,比较不同立地条件和林分密度下林分碳储量生长过程差异,分析各速生阔叶树种碳汇潜力,旨在为碳汇造林提供决策依据。
1 材料与方法
1.1 数据特征
采用昆明市第4次森林资源二类调查数据,选取主要速生阔叶树种桤木、银荆、蓝桉和直干桉组成系数大于6成的相对纯林林小班作为研究对象,以小班为建模样本单元,测定各龄阶林分平均林龄、平均树高、平均胸径、密度、断面积为准,以±3倍标准差为界剔除该龄阶范围内的异常点后进行样本量、平均值、中位数和标准差的描述性统计(表1)。
1.2 研究方法
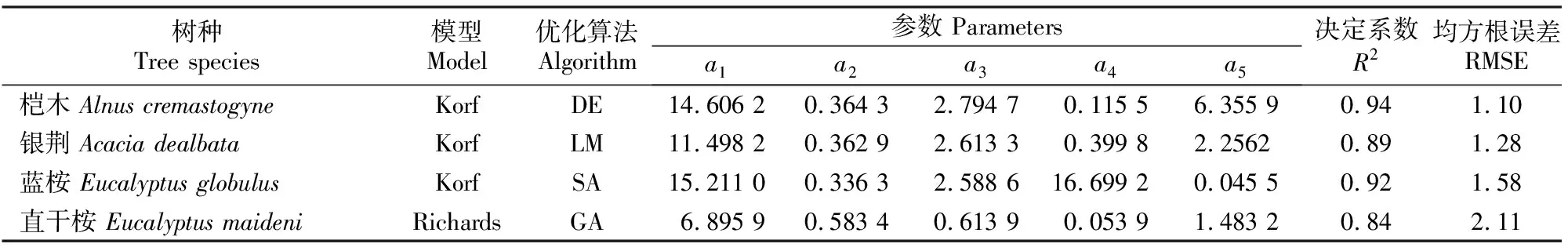
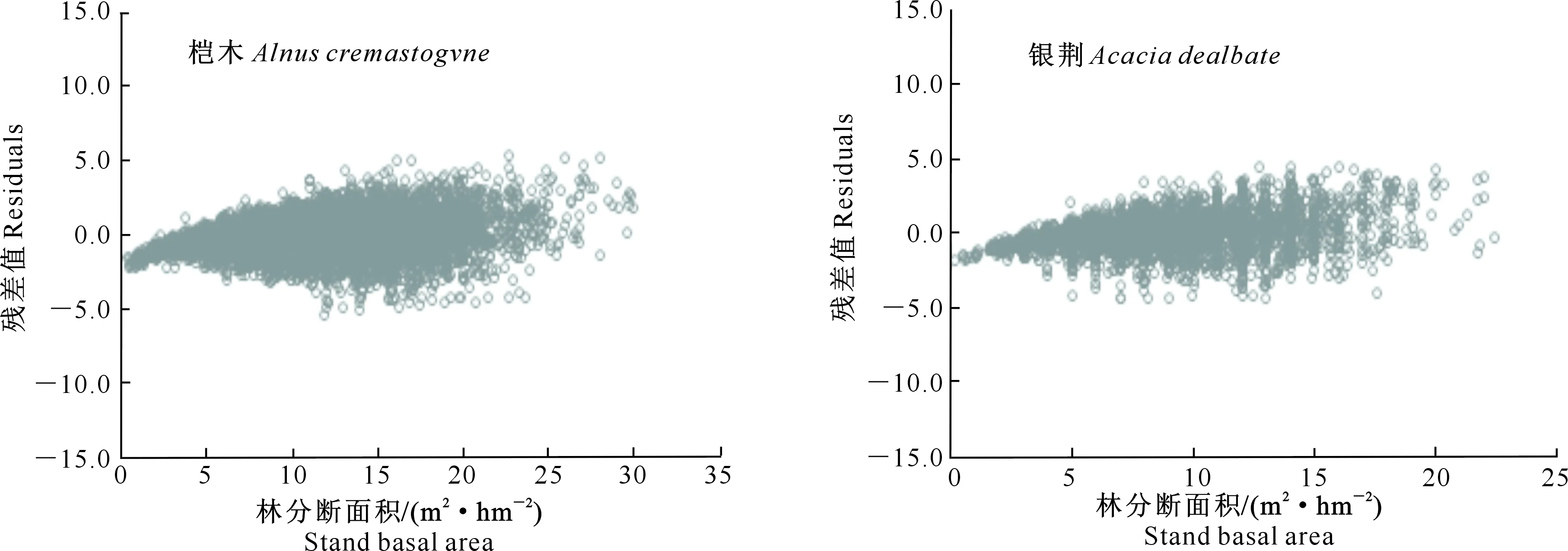
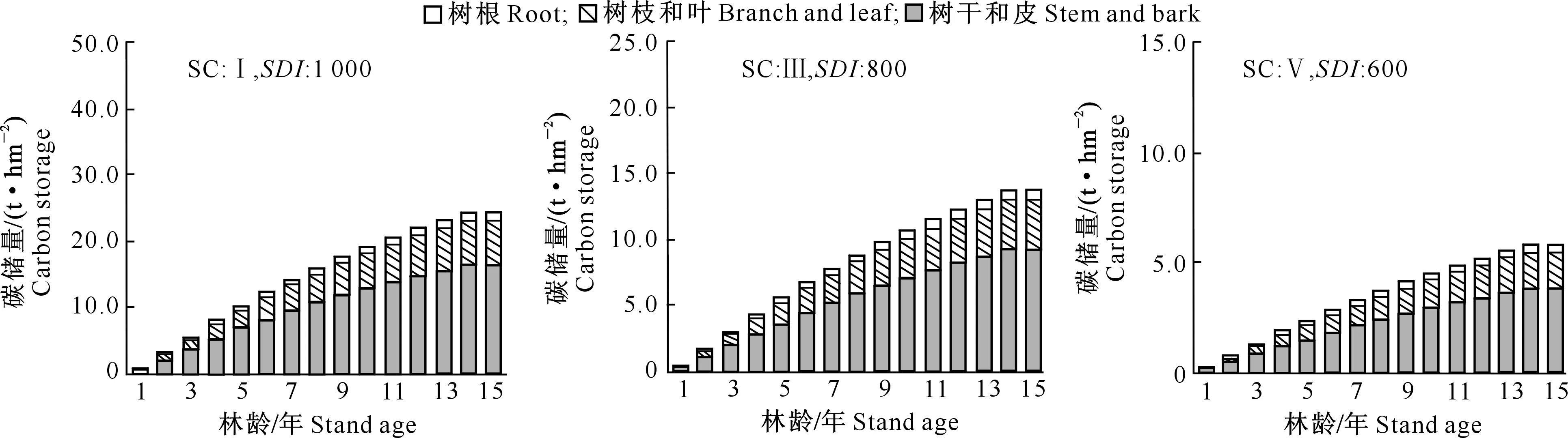
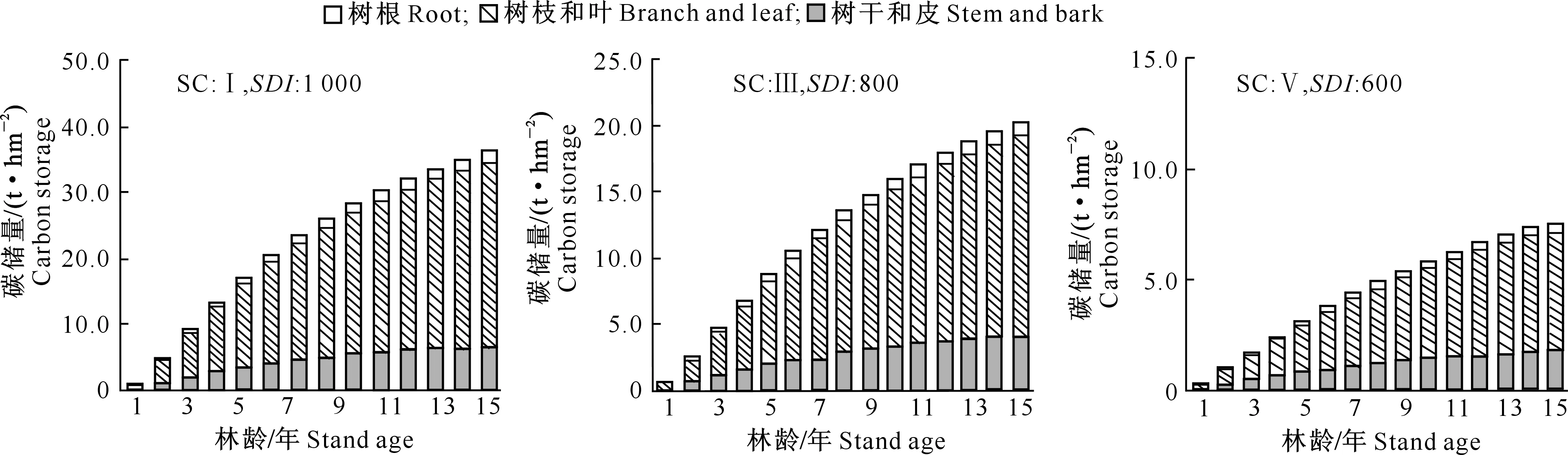
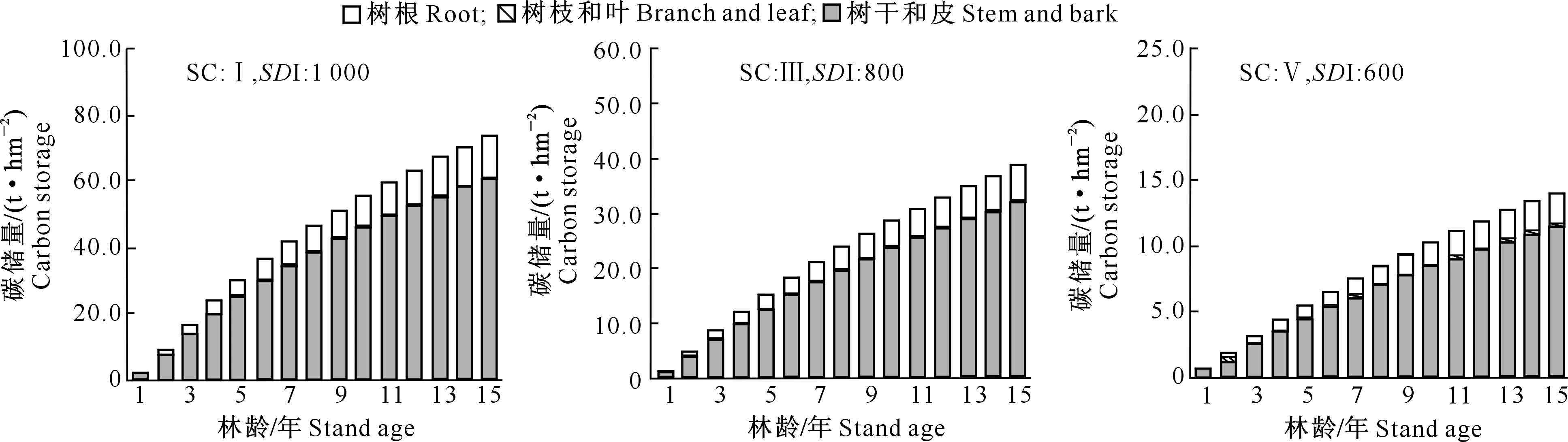
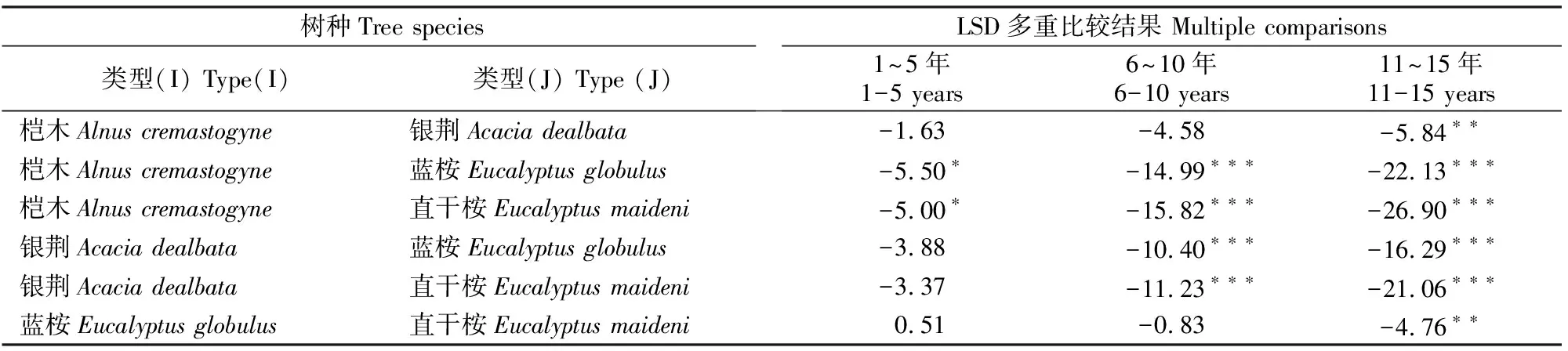
1.2.1 断面积生长模型构建 在全林分生长模型中,森林的生长和收获取决于林龄、立地、林分密度等3个主要因子。立地质量评价采用Schumacher、Richards和Compertz拟合林龄-树高生长导向曲线,建立4个速生阔叶树种地位级指数(SCI)模型。采用胸径(Dg)和林分密度(N)的全部样本观测值对回归方程进行拟合,估计得到a1和b1值后,删除lnN (1) 式中:N为现实林分密度,D0为标准平均胸径(本研究D0=20 cm),Dg为现实林分平均胸径,b为完满立木度和平均胸径间方程斜率。 表1 建模速生阔叶树种样本基本特性的描述性统计分析Table 1 Descriptive statistics of fast growing broad-leaf species for modeling 采用SDI作为竞争指标,将其引入到断面积生长模型中,拟合不同立地条件下林分生长模型,其分别是Schumacher(式(2))、Richards(式(3))和Korf(式(4))的变型: (2) (3) (4) 式中:G为林分断面积,A为林分平均林龄,SCI为地位级指数,SDI为林分密度指数,a1、a2、a3、a4和a5为参数。 采用麦夸特算法(LM)、差分进化算法(DE)、遗传算法(GA)、模拟退火算法(SA)和粒子群算法(PSO)极小化离差平方和,运用Matlab2014b进行最优值求解,得到不同优化算法下林分断面积生长模型参数,根据拟合决定系数(R2)、均方根误差(RMSE),综合考虑拟合参数稳定性,从中筛选一组参数估计值作为模型拟合结果。根据参数求解迭代次数比较优化算法的效能。采用欧氏距离计算各参数拟合结果间的差异,比较不同模型参数的拟合稳定性,欧氏距离计算公式如下: (5) 1.2.2 生长模型与生物量模型链接 林分生物量常采用等段面积分级标准木法进行测定,工作量大且费时耗力。通过林分生长模型与生物量模型,利用已知的林分因子(如胸径和树高)推算林分各器官生物量,累加即得到林分生物量。本研究采用李贵祥等[13]建立的滇中高原桤木生物量模型,张加正等[14]建立的银荆生物量模型,高永能等[15]建立的蓝桉、直干桉生物量模型以及桉树地上部和地下部生物量模型,最终建立桤木、银荆、蓝桉和直干桉断面积生长模型与树高生长模型,进行各器官林分生物量的计算。 1.2.3 速生阔叶树种林分碳储量差异性分析 植被碳储量需要根据植物碳储量进行求算,本研究采用重铬酸钾-硫酸氧化法[12]测定桤木各器官含碳率,计算桤木林分各器官碳储量;采用干烧法[10]测定桉树各器官含碳率,计算蓝桉和直干桉林分各器官碳储量;采用向玮等[7]推算的林分碳储量转化系数0.50,计算银荆林分各器官的碳储量。将各速生阔叶树种生长过程分为1~5,6~10,11~15年3个阶段,进行单因素方差分析和最小显著性(LSD)多重比较,分析各速生阔叶树种碳储量生长过程的差异。比较各速生阔叶树种碳储量平均增长量和连年增长量,分析各树种林分碳汇潜力。平均生长量为林分某一时期的碳储量增长速度,连年增长量为林分某一年的实际碳储量增长速度。 采用优化算法拟合的速生阔叶树种林分断面积生长模型见表2。 表2 速生阔叶树种林分断面积生长模型的拟合结果Table 2 Fitting results of stand basal area growth for fast growing broad-leaf species 注:DE表示差分进化算法,LM表示麦夸特算法,SA表示模拟退火算法,GA表示遗传算法。 Note:DE means differential evolution algorithm,LM means levenberg-marquardt algorithm,SA means simulated annealing algorithm,and GA means genetic algorithm. 从表2可以看出,桤木、银荆和蓝桉均采用Korf模型形式,直干桉采用Richards模型形式。各树种生长模型拟合决定系数介于0.84~0.94,平均值为0.90;生长模型拟合结果均方根误差介于1.10~2.11,平均值为1.52。各优化算法在速生阔叶树种林分断面积生长模型参数求解中的迭代次数依次为麦夸特算法(LM)迭代52次,差分进化算法(DE)迭代1 592次,遗传算法(GA)迭代4 540次,模拟退火算法(SA)迭代4 539次,粒子群算法(PSO)迭代次数1 291次;不同模型形式对优化算法求解参数效能也存在差异,各模型形式参数求解中迭代次数依次为:Richards模型拟合迭代3 060次,Schumacher模型拟合迭代2 127次,Korf模型拟合迭代2 021次。Richards模型各参数拟合结果间欧式距离为314.82,Schumacher模型各参数拟合结果间欧式距离为7.88,Korf模型各参数拟合结果间欧式距离为10.03。采用Schumacher模型和Korf模型拟合参数稳定性优于Richards模型,但Korf模型的拟合优度大于Schumacher模型。 林分断面积生长模型拟合残差分布见图1~2。由图1可知,模型预测值与观测值间的残差散点均匀分布在以0为起点的横轴两侧,不存在系统性偏差。 图1 桤木和银荆林分断面积生长模型拟合残差的分布Fig.1 Residual distribution of growth model of stand basal area for Alnus cremastogvne and Acacia dealbate 图2 蓝桉和直干桉林分断面积生长模型残差的分布Fig.2 Residual distribution of growth model of stand basal area for Eucalyptus globulus and Eucalyptus maideni 桤木、银荆、蓝桉和直干桉模型预测值与观察值间均值检验的T统计量分别为0.02,0.08,0.14和0.08,自由度分别为25 574,7 472,10 222和3 626,显著性P值分别为0.80,0.88,0.84和0.99,即在α=0.10显著性水平上,模型预测值与观测值间没有差异。通过模型拟合残差分析和均值检验表明,本研究建立的速生阔叶树种断面积生长模型具有较好的预测效能,精度能满足林分生长分析的要求。 基于建立的速生阔叶树种生长模型与相应的生物量模型链接,再乘以各器官含碳率即可得到各速生阔叶树种地位级(SC)分别为Ⅰ、Ⅲ、Ⅴ,林分密度指数(SDI)分别为1 000,800,600时碳储量生长过程(图3~6)。以地位级Ⅲ级、林分密度指数800为例,说明各速生阔叶树种林分碳储量生长过程。 图3 不同立地(SC)和密度指数(SDI)条件下桤木林分碳储量生长过程Fig.3 Carbon storage process of Alnus cremastogyne with different site classes and stand densities 图4 不同立地(SC)和密度指数(SDI)条件下银荆林分碳储量生长过程Fig.4 Carbon storage process of Acacia dealbata with different site classes and stand densities 由图3~6可知,各树种林分碳储量随林龄增加呈上升趋势。至林龄为15年时,桤木、银荆、蓝桉和直干桉林分碳储量分别为13.81,20.33,38.89和45.27 t/hm2,蓝桉和直干桉林分碳储量增长速率大于桤木和银荆林分;桤木林分树干和皮碳储量占总碳储量的比例平均值为66.80%,银荆林分树枝和叶碳储量占总碳储量的比例平均值为71.19%,蓝桉林分树干和皮碳储量占总碳储量的比例平均值为81.54%,直干桉林分树干和皮碳储量占总碳储量的比例平均值为79.78%。 图5 不同立地(SC)和密度指数(SDI)条件下蓝桉林分碳储量生长过程Fig.5 Carbon storage process of Eucalyptus globulus with different site classes and stand densities 图6 不同立地(SC)和密度指数(SDI)条件下直干桉林分碳储量生长过程Fig.6 Carbon storage process of Eucalyptus maiden with different site classes and stand densities 根据单因素方差分析表明,林龄为1~5年时各树种林分间碳储量生长不存在差异(α=0.10),林龄为6~10和11~15年时各树种林分间碳储量生长存在极显著差异(α=0.01)。多重比较结果(表3)表明,林龄为1~5年时桤木林分与蓝桉和直干桉林分间碳储量生长存在显著差异(α=0.10),其余各林分间差异不显著;林龄为6~10年时桤木与蓝桉、直干桉林分间,银荆与蓝桉、直干桉林分间存在极显著差异(α=0.01);林龄为11~15年时桤木与银荆林分间,蓝桉与直干桉林分间存在显著差异(α=0.05),桤木与蓝桉、直干桉林分间,银荆林分与蓝桉、直干桉林分间存在极显著性差异(α=0.01)。 表3 速生阔叶树种林分不同阶段碳储量生长差异多重比较结果Table 3 Multiple comparison of carbon storage at different stages of fast growing species 注:*表示在α=0.10水平差异显著,**表示在α=0.05水平差异显著,***表示在α=0.01水平差异极显著。 Note:* means difference (α=0.10), ** means significant difference (α=0.05), and *** means extremely significant difference (α=0.01). 以地位级为Ⅲ级、林分密度指数为800的林分碳储量平均增长量与连年增长量分析各树种碳汇潜力,结果见图7。由图7可以看出,随着林分年龄增加,各树种林分碳储量平均增长量和连年增长量均呈现先快速生长后再缓慢下降的趋势。桤木、银荆、蓝桉和直干桉林分碳储量平均增长量最大值分别为1.14,1.76,3.10和3.11 t/(hm2·年),林分碳储量连年增长量最大值分别为1.34,2.18,3.66和3.41 t/(hm2·年)。林龄为2~6年直干桉林分碳储量年平均增长量与连年增长量小于蓝桉林分,林龄为7~15年直干桉林分碳储量年平均增长量与连年增长量大于蓝桉林分,直干桉和蓝桉林分碳储量年平均增长量与连年增长量均大于银荆和桤木林分,银荆林分碳储量年平均增长量与连年增长量均大于桤木林分。 图7 速生阔叶树种林分碳储量平均增长量和连年增长量(SC:Ⅲ,SDI:800)Fig.7 Mean increment and annual increment of stand carbon storage of fast growing species (SC:Ⅲ,SDI:800) 林分生长模型与单木生物量生长模型的链接能对林分生产力进行较好预测。张加正等[14]以银荆林分为例发现, 6年现实林分生物量生产力为36.71 t/hm2,年平均生物量生产力为6.12 t/(hm2·年)。本研究基于生长模型预测6年现实林分生物量生产力为36.94 t/hm2,年平均生物量生产力为6.16 t/(hm2·年),连年生物量生产力为6.02 t/(hm2·年)。当进行经营调整使银荆林分保留株数达到2 850株/hm2时,林分生物量生产力为48.43 t/hm2,本研究预测的林分生物量生产力为45.49 t/hm2。说明林分生长模型与单木生物量模型链接进行林分生产力预测具有较好的精确性。 森林碳汇估算方法中生物量法[2,22]采用单位面积生物量、森林面积、生物量在树木各器官中的分配比例、树木各器官的平均碳含量等参数进行计算。但林分生物量的积累不仅与树种本身有关,还与立地质量和林分密度等因素有关。蓄积量法[3]根据主要树种的平均容重,再根据生物量与含碳率计算森林碳储量,蓄积量法操作简便、实用性强,但含碳率的运用只考虑了树种差异,仍然存在较大误差[17]。生物量清单法[4]将生态学调查资源和森林普查资料结合计算森林碳储量。用涡旋相关法能够直接长期对森林生态系统进行CO2通量测定,同时能够为其他模型的建立和校准提供基础数据。不同的碳汇计量方法有各自优缺点,链接林分生长模型进行碳汇计量估算[7-8]可以分树种、年龄、立地质量和林分密度等进行,在实际运用中更为直接和精确。陶玉华等[23]对杉木人工林碳储量的研究表明,中龄林杉木总碳储量略高于成熟林,与现实林分的生长状况存在差异,这种差异来源于林分立地质量和林分密度,用林分生长模型与单木生物量模型链接则能较好地解决立地质量和密度对林分碳储量生长过程的影响。李斌等[24]对湖南省杉木林的碳储量和碳密度研究表明,杉木林碳密度和碳储量随着林龄的增加而增高,这与本研究结果一致。 生物量推算碳储量一般采用0.50作为转化系数[7],但含碳率因树种和器官而存在差异。Bert和Danjon[25]对法国西南部海岸松(Pinuspinaster)碳储量估计的研究表明,采用通用转化系数0.50会造成森林碳储量的低估。本研究银荆林分碳储量估算采用了0.50的转换系数,可能导致银荆林分碳储量估计偏差,在后续的研究中需要完善银荆各器官含碳率本土化参数。

2 结果与分析
2.1 速生阔叶树种林分断面积生长模型拟合结果

2.2 速生阔叶树种林分碳储量生长过程
2.3 速生阔叶树种林分碳汇潜力分析

3 讨 论
