TaGAMyb-B等位变异与小麦株高和第二节间细胞长度的相关性
2018-07-27李淑芬刘进英冯玉梅
刘 梦,王 增,李淑芬,刘进英,冯玉梅,杨 燕
(内蒙古农业大学生命科学学院植物生物技术功能实验室/内蒙古自治区植物逆境生理与分子生物学重点实验室,内蒙古呼和浩特 010018)
小麦株高不仅影响产量,同时还与抗倒伏性密切相关[1]。在育种实践中,一般认为株高分布在75~85 cm的小麦品种不易发生倒伏,且能发挥高产潜能[2]。小麦[3-4]、大麦[5]和燕麦[6]的植株茎秆直径和茎壁厚度与抗倒伏性呈负相关,即抗倒伏品种较易倒伏品种表现出较大的基部直径和较厚的秆壁[7]。小麦茎秆基部第一和第二节间长度与倒伏指数呈显著负相关,且基部节间对小麦倒伏性的影响表现为第二节间>第一节间>第三节间[8]。李金才等[9]的研究也验证了小麦茎秆基部第二节间长度与倒伏性密切相关。在由表皮、维管束、厚壁组织和髓腔组成的小麦茎秆横切面中,厚壁组织的细胞壁坚硬而起到支持作用,与植株倒伏性状密切相关[10]。
赤霉素作为植物重要的内源性生长调节剂,在植物整个生长发育过程中起到重要的调节作用。GAMyb是参与GA信号转导途径的正向调节因子,能够促进种子成熟,胚乳和花的发育、茎秆伸长等过程[11-13]。目前,GAMyb同源基因已经从大麦、水稻、小麦禾本科植物中鉴定出来,并确定其为单拷贝基因,通过结合到一个高度保守的由21个碱基对组成的GA应答元件来诱导与发芽相关的水解蛋白酶和细胞壁降解酶的合成[14-18]。同时GAMyb也与植株的茎秆伸长密切相关[19]。在拟南芥中,GAMyb却是一个具有三个拷贝的家族,并促进叶柄和节间伸长[20]。近些年对小麦TaGAMyb的研究较广泛和深入,TaGAMyb已经被克隆并定位在小麦染色体的3AL3-0.42-0.78、3BL7-0.63-1.00和3DL3-0.81-1.00位置上,且B和D基因组TaGAMyb的核酸多态性较保守的A基因组来说更丰富[20]。TaGAMyb基因在花药形成和茎秆的伸长中起重要作用[19,21]。进一步研究发现,在杂交小麦后代中TaGAMyb基因对茎伸长的调节作用可能是因为TaGAMyb基因上调,导致GAMyb调节靶基因表达的增强[22]。这些数据初步表明,TaGAMyb促进小麦茎的伸长,但其在节间伸长中的作用需要进一步的验证。刘进英等[23]在不同株高和不同遗传背景材料中没有发现TaGAMyb-A和TaGAMyb-D基因存在等位变异,只发现TaGAMyb-B存在两种等位变异TaGAMyb-Ba和TaGAMyb-Bb;与TaGAMyb-Ba基因相比,TaGAMyb-Bb基因在第一内含子上存在一个84 bp反向重复序列的插入。本研究在本课题组前期工作[23]的基础之上,进一步探讨TaGAMyb-Ba和TaGAMyb-Bb与小麦株高和第二节间细胞长度的关系,以期深入了解这两种等位变异对小麦茎秆生长发育的调节作用。
1 材料与方法
1.1 试验材料
供试材料为93份春小麦育种材料的自然群体,由内蒙古巴彦淖尔市农科所提供。2015年和2016年播种于内蒙古呼和浩特市,行长2 m,株距15 cm,行距20 cm,设置两个重复。
1.2 株高测定
在小麦腊熟期,对93份春小麦育种材料进行株高测量,每个重复统计10株。
1.3 引物的设计
根据TaGAMyb-A、TaGAMyb-B和TaGAMyb-D基因序列差异位点[22]以及RT-qPCR引物片段适中性原则设计引物对TaGAMyb-Bup/TaGAMyb-Bdown(表1),对第二节间的TaGAMyb-B基因进行RT-qPCR定量检测,同时用小麦管家基因β-actin表达量作为TaGAMyb-B基因相对表达量的参照标准,引物为Actin up/Actin down(序列见表1)。用检测TaGAMyb-B等位基因的引物GAMyb-B2 up/GAMyb-B2 down(表1)[23]对93份春小麦材料进行多态性检测,所用引物序列均由华大基因合成。
1.4 TaGAMyb-B的RT-qPCR检测分析
随机选取分别属于TaGAMyb-Ba和TaGAMyb-Bb等位基因类型的材料各三份,分别是08冬中788、08冬中1146和08冬中275(TaGAMyb-Bb类型)以及格兰尼、拉繁8和青春37(TaGAMyb-Ba类型),在小麦开花后30 d取基部第二节间为试验材料,进行TaGAMyb-B基因RT-qPCR检测分析。利用MiniBEST Plant RNA Extraction Kit(TaKaRa)提取总RNA,并取2 μL进行1%琼脂糖凝胶电泳分析检测总RNA的完整性,用核酸定量仪(Thermo Nanodrop-2000)测定核酸的浓度和检测总RNA的纯度。以总RNA为模板,利用反转录试剂盒RTaseM-MLV(TaKaRa)(200 U·μL-1)合成第一条cDNA链。以反转录得到的第一条cDNA链为模板,利用小麦管家基因β-actin检测反转录过程的完整性。反应体系(15 μL):ddH2O 10.55 μL、10×PCR Buffer 1.5 μL、dNTP Mix(2.5 mmol·L-1) 1.2 μL、上下游引物(20 mmol·L-1)各0.3 μL、LA Taq酶(5 U·μL-1,TaKaRa)0.15 μL和模板cDNA 100 ng。反应条件:94 ℃预变性5 min;94 ℃变性30 s,62 ℃退火30 s,72 ℃延伸30 s,35个循环;72 ℃延伸10 min。PCR产物用1%琼脂糖凝胶检测。
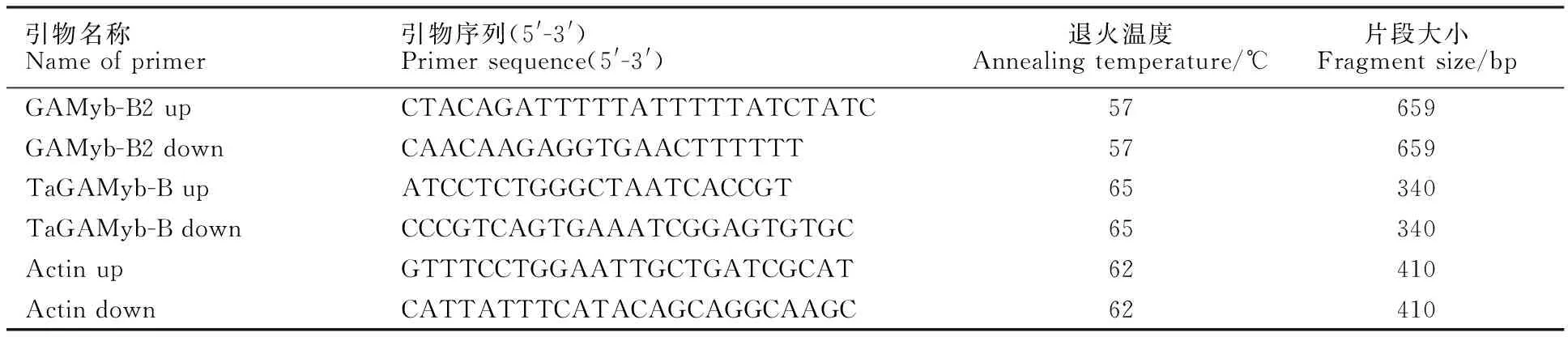
表1 本研究引物信息Table 1 Information of primers in this study
RT-qPCR分析反应体系为20 μL,包括SYBR Premix Ex Taq Ⅱ 10 μL,上下游引物(20 mmol·L-1)各1 μL,ddH2O 6 μL,cDNA(500 ng·μL-1)2 μL。每个材料进行三次生物学重复,反应条件为:94 ℃预变性5 min,94 ℃ 20 s,56 ℃ 20 s,循环40次。电脑自动分析计算CT值。
样品中基因表达量变化倍数运用2-△△CT法计算[24]。其中△△CT=△CT(参照)-△CT(处理),△CT=CT(目标基因)-CT(内参基因)。
1.5 冷冻切片样品的制备与观察
选取小麦开花后30 d茎秆基部第二节间作为试验材料。样品剪成约0.5 cm的小段,用LEICA OCT包埋剂进行包埋。放于-20 ℃冰箱冷藏2 h,然后放于-80 ℃备用。用LEICA CM3050S冷冻切片机进行切片,厚度为20 μm。用小毛刷将切片展平,用粘附载玻片粘样,最后放于Nikon eclipse Ti下进行观察,选用绿色滤光片。
1.6 数据的分析
用Microsoft Excel 2010对株高数据进行差异显著性分析,用SPSS对株高数据进行相关性分析。
2 结果与分析
2.1 春小麦株高表型的差异
测定结果显示,两年中,93份春小麦株高分布范围为51.25~97.97 cm,平均值为70.49 cm。其中,株高最高材料为拉繁8(97.97 cm),株高最低材料为08冬中788(51.25 cm)。93份春小麦材料中,株高在75~85 cm间的有16份材料,占比为17.2%(表2)。该群体在2015年和2016年的株高呈极显著正相关(R=0.719,P<0.01),说明两年的株高表现是一致的。
2.2 TaGAMyb-B等位基因的鉴定结果及相应材料株高分布
用引物GAMyb-B2 up/GAMyb-B2 down对93份春小麦材料进行多态性检测,在93份材料中检测出TaGAMyb-Ba和TaGAMyb-Bb两种等位基因(图1和表2),其中,有31份小麦材料属于TaGAMyb-Ba等位基因型,62份小麦材料属于TaGAMyb-Bb等位基因型。TaGAMyb-Ba等位基因型株高分布范围为51.45~97.97 cm,最高株高材料为拉繁8(97.97 cm),最低株高材料为宁春10号(51.45 cm);分布在最适种植株高75~85 cm范围的有8份材料,分别是05cm178、鉴56、沈太1号、拉2676-9、青春37、宁资08A751、34-206和沈免210848,占比为25.80%,株高分别为77.28、83.96、82.45、80.51、79.44、76.81、75.96和75.89 cm。TaGAMyb-Bb等位基因型的株高分布范围为51.25~91.49 cm,最高株高材料为中国春(91.49 cm),最低株高材料为08冬中788(51.25 cm);分布在75~85 cm范围的也有8份材料,分别为九三大穗/山东7788、甘春20号、小冰32、内麦17号、08H274、G47、05-5371和张春11,占比为12.90%,株高分别为83.44、81.63、80.27、79.89、79.44、77.93、76.88和75.08 cm。由此可见,该自然群体中TaGAMyb-Ba等位基因型材料在最适种植株高75~85 cm范围中的占比大于TaGAMyb-Bb等位基因型材料。
2.3 TaGAMyb-Ba和TaGAMyb-Bb与小麦株高的相关性
2015和2016年TaGAMyb-Ba基因型的株高平均值分别为70.00±9.36和76.86±9.57 cm,两年均以拉繁8最高,宁春10号最低;TaGAMyb-Bb基因型的株高平均值两年分别为65.79±7.81和72.07±9.74 cm,两年分别以野猫和河套3号最高,以08冬中550和08冬中788最低。方差分析表明,同一年内两种基因型间株高差异均显著(P2015=0.024,P2016=0.027);两个基因型的两年平均值分别为73.43±8.62和68.93±8.18 cm,也存在显著差异(P=0.016)。
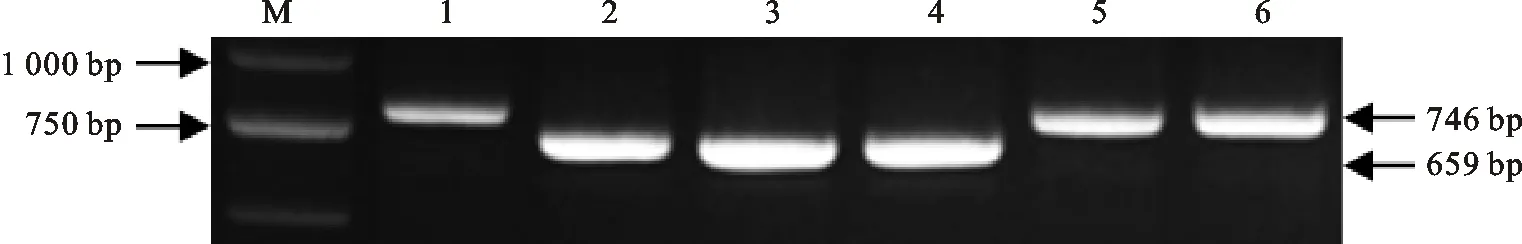
M:Marker DL2000;1、5和6均为TaGAMyb-Bb等位基因型材料(08冬中788、08冬中1146和08冬中275);2、3和4均为TaGAMyb-Ba等位基因型材料(格兰尼、拉繁8和青春37)。
M:Marker DL2000; 1,5 and 6 areTaGAMyb-Bballele materials(08 dongzhong 788,08 dongzhong 1146 and 08 dongzhong 275); 2,3 and 4 are cultivars withTaGAMyb-Baallele(Gelanni,Lafan 8 and Qingchun 37).

图1 GAMyb-B等位基因在93份春小麦材料中的部分扩增结果
(续表2Continuedtable2)
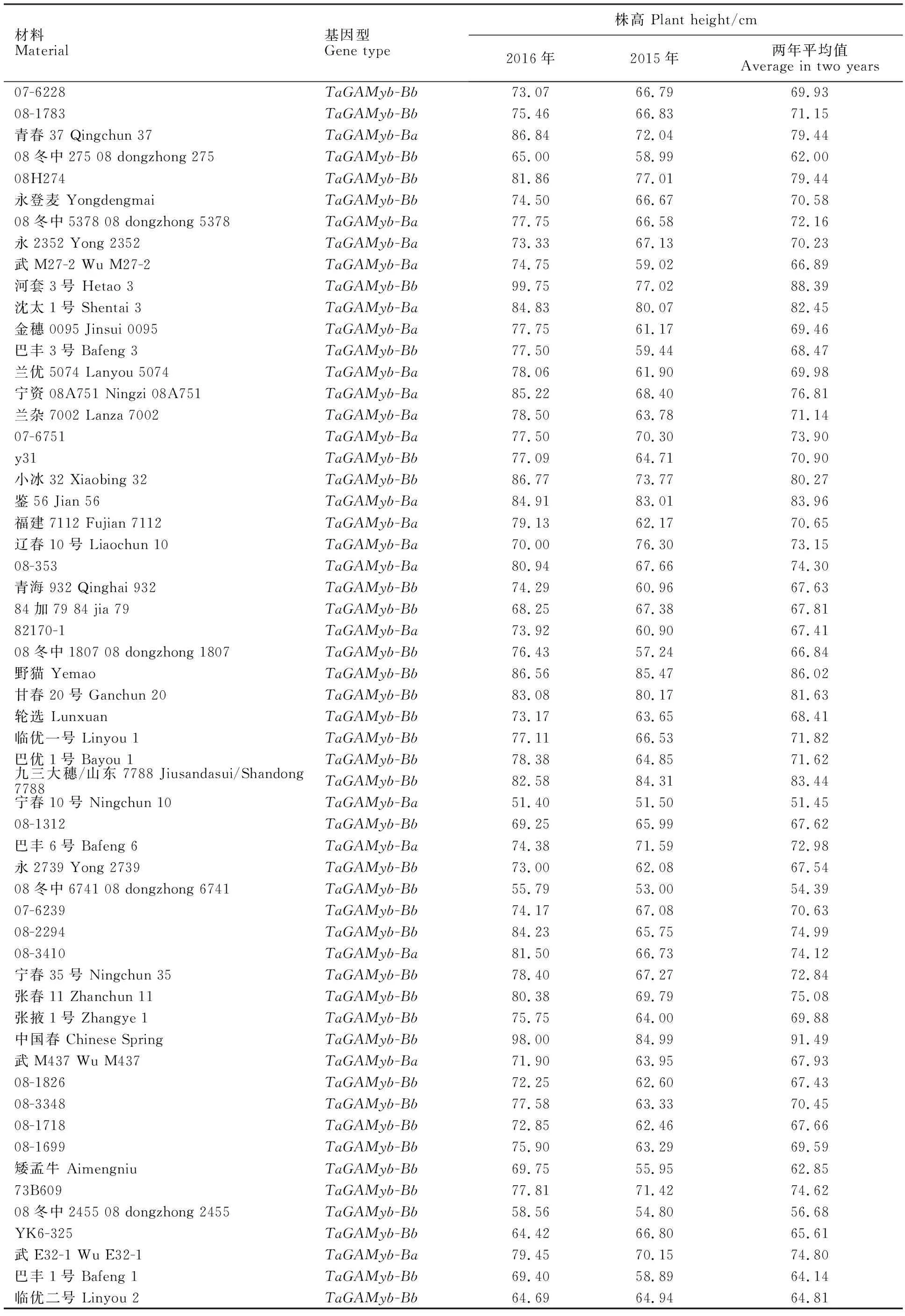
材料Material基因型Gene type株高 Plant height/cm2016年2015年两年平均值Average in two years07-6228TaGAMyb-Bb73.07 66.79 69.93 08-1783TaGAMyb-Bb75.46 66.83 71.15 青春37 Qingchun 37TaGAMyb-Ba86.84 72.04 79.44 08冬中275 08 dongzhong 275TaGAMyb-Bb65.00 58.99 62.00 08H274TaGAMyb-Bb81.86 77.01 79.44 永登麦 YongdengmaiTaGAMyb-Bb74.50 66.67 70.58 08冬中5378 08 dongzhong 5378TaGAMyb-Ba77.75 66.58 72.16 永2352 Yong 2352TaGAMyb-Ba73.33 67.13 70.23 武M27-2 Wu M27-2TaGAMyb-Ba74.75 59.02 66.89 河套3号 Hetao 3TaGAMyb-Bb99.75 77.02 88.39 沈太1号 Shentai 3TaGAMyb-Ba84.83 80.07 82.45 金穗0095 Jinsui 0095TaGAMyb-Ba77.75 61.17 69.46 巴丰3号 Bafeng 3TaGAMyb-Bb77.50 59.44 68.47 兰优5074 Lanyou 5074TaGAMyb-Ba78.06 61.90 69.98 宁资08A751 Ningzi 08A751TaGAMyb-Ba85.22 68.40 76.81 兰杂7002 Lanza 7002TaGAMyb-Ba78.50 63.78 71.14 07-6751TaGAMyb-Ba77.50 70.30 73.90 y31TaGAMyb-Bb77.09 64.71 70.90 小冰32 Xiaobing 32TaGAMyb-Bb86.77 73.77 80.27 鉴56 Jian 56TaGAMyb-Ba84.91 83.01 83.96 福建7112 Fujian 7112TaGAMyb-Ba79.13 62.17 70.65 辽春10号 Liaochun 10TaGAMyb-Ba70.00 76.30 73.15 08-353TaGAMyb-Ba80.94 67.66 74.30 青海932 Qinghai 932TaGAMyb-Bb74.29 60.96 67.63 84加79 84 jia 79TaGAMyb-Bb68.25 67.38 67.81 82170-1TaGAMyb-Ba73.92 60.90 67.41 08冬中1807 08 dongzhong 1807TaGAMyb-Bb76.43 57.24 66.84 野猫 YemaoTaGAMyb-Bb86.56 85.47 86.02 甘春20号 Ganchun 20TaGAMyb-Bb83.08 80.17 81.63 轮选 LunxuanTaGAMyb-Bb73.17 63.65 68.41 临优一号 Linyou 1TaGAMyb-Bb77.11 66.53 71.82 巴优1号 Bayou 1TaGAMyb-Bb78.38 64.85 71.62 九三大穗/山东7788 Jiusandasui/Shandong 7788TaGAMyb-Bb82.58 84.31 83.44 宁春10号 Ningchun 10TaGAMyb-Ba51.40 51.50 51.45 08-1312TaGAMyb-Bb69.25 65.99 67.62 巴丰6号 Bafeng 6TaGAMyb-Ba74.38 71.59 72.98 永2739 Yong 2739TaGAMyb-Bb73.00 62.08 67.54 08冬中6741 08 dongzhong 6741TaGAMyb-Bb55.79 53.00 54.39 07-6239TaGAMyb-Bb74.17 67.08 70.63 08-2294TaGAMyb-Bb84.23 65.75 74.99 08-3410TaGAMyb-Ba81.50 66.73 74.12 宁春35号 Ningchun 35TaGAMyb-Bb78.40 67.27 72.84 张春11 Zhanchun 11TaGAMyb-Bb80.38 69.79 75.08 张掖1号 Zhangye 1TaGAMyb-Bb75.75 64.00 69.88 中国春 Chinese SpringTaGAMyb-Bb98.00 84.99 91.49 武M437 Wu M437TaGAMyb-Ba71.90 63.95 67.93 08-1826TaGAMyb-Bb72.25 62.60 67.43 08-3348TaGAMyb-Bb77.58 63.33 70.45 08-1718TaGAMyb-Bb72.85 62.46 67.66 08-1699TaGAMyb-Bb75.90 63.29 69.59 矮孟牛 AimengniuTaGAMyb-Bb69.75 55.95 62.85 73B609TaGAMyb-Bb77.81 71.42 74.62 08冬中2455 08 dongzhong 2455TaGAMyb-Bb58.56 54.80 56.68 YK6-325TaGAMyb-Bb64.42 66.80 65.61 武E32-1 Wu E32-1TaGAMyb-Ba79.45 70.15 74.80 巴丰1号 Bafeng 1TaGAMyb-Bb69.40 58.89 64.14 临优二号 Linyou 2TaGAMyb-Bb64.69 64.94 64.81
2.4 TaGAMyb-B基因在小麦茎秆第二节间中的表达
在小麦茎秆第二节间中,TaGAMyb-Ba基因的表达量均高于TaGAMyb-Bb基因,但差异不显著(P=0.120)(图2)。
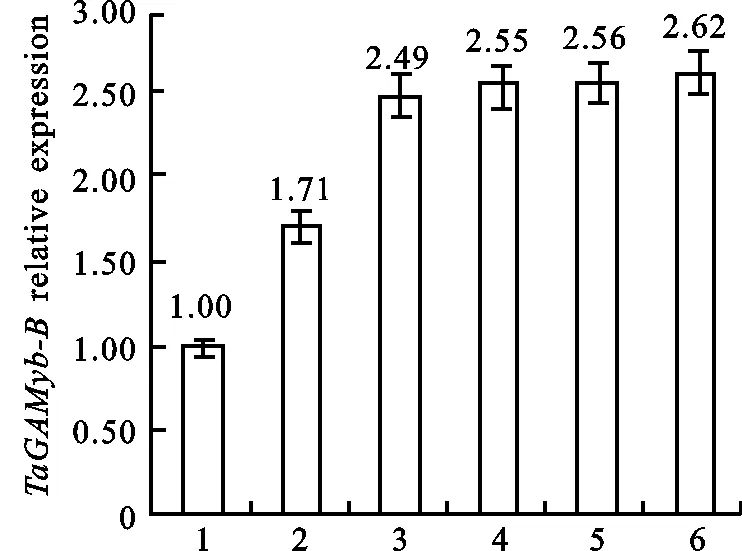
1:08冬中788(TaGAMyb-Bb);2:08冬中1146(TaGAMyb-Bb);3:08冬中275(TaGAMyb-Bb);4:格兰尼(TaGAMyb-Ba);5:拉繁8(TaGAMyb-Ba);6:青春37(TaGAMyb-Ba)。相对表达量以1(08冬中788)为标准进行均一化处理。
1:08 dongzhong 788(TaGAMyb-Bb); 2:08 dongzhong 1146(TaGAMyb-Bb); 3:08 dongzhong 275(TaGAMyb-Bb); 4:Gelanni(TaGAMyb-Ba); 5:Lafan(TaGAMyb-Ba); 6:Qingchun 37(TaGAMyb-Ba). The relative expression was homogenized with 08 dongzhong 788 as the standard.
图2TaGAMyb-B在小麦茎秆第二节间中的表达量
Fig.2ExpressionofTaGAMyb-Binthesecondinternodeofwheatstem
2.5 小麦茎秆第二节间厚壁组织的变异
对6份小麦材料茎秆第二节间横切面(图3)的厚壁组织厚度进行测量,结果表明,属于TaGAMyb-Bb基因型的08冬中788、08冬中1146和08冬中275的厚壁组织厚度分别为990.52、563.68和904.31 μm;属于TaGAMyb-Ba基因型的格兰尼、拉繁8和青春37的厚壁组织厚度分别为1 006.68、854.65和853.66 μm。进一步分析发现,两种基因型材料间厚壁组织厚度无显著差异(P=0.57)。对这6份材料第二节间进行纵切,除拉繁8没能切出完整细胞外,其他5份材料均有完整细胞(图4)。统计每种材料中连续分布的30个厚壁细胞的长度,结果表明,TaGAMyb-Ba基因型材料格兰尼和青春37的厚壁细胞平均长度分别为240.67和267.45 μm,二者间差异不显著;属于TaGAMyb-Bb基因型的08冬中788、08冬中1146和08冬中275厚壁组织细胞的平均长度分别为212.70、202.47和185.28 μm,三者间差异不显著。TaGAMyb-Ba和TaGAMyb-Bb基因型材料的厚壁组织细胞平均长度分别为254.06和200.15 μm,二者间差异显著(P=0.03)。

图3 两种等位基因类型小麦的第二茎间横切面
3 讨 论
本研究中,TaGAMyb-Ba基因型小麦两年平均株高(73.43 cm)显著高于TaGAMyb-Bb基因型小麦(68.93 cm)(P<0.05)。然而田间表型鉴定中,有部分材料却与此结论相悖。例如在TaGAMyb-Ba基因型小麦中,交原356和宁春10号的平均株高分别为58.10和51.45 cm;在TaGAMyb-Bb基因型小麦中,中国春、野猫和河套3号的平均株高分别为91.49、86.02和88.39 cm。这些材料的株高均与相应变异类型的平均值差异较大,原因可能是株高作为一个多基因控制的数量性状,在其建成过程中,除受TaGAMyb-B基因调控之外,还受 Rht8、Rht-B1、 Rht-D1、GA20ox、GA3ox等其他遗传因素[17,24]以及环境因素的共同影响[27]。对6份材料茎秆第二节间的TaGAMyb-B相对表达量与相应株高的相关分析结果显示,两者之间没有显著的相关性(R=0.768,P=0.074>0.05),可能是由于试验样本少造成了结果的局限性。青春37(TaGAMyb-Ba)茎秆第二节间的厚壁细胞最长,并且在6份材料中TaGAMyb-Ba的表达量最高,但其平均株高却不是最高。这一结果进一步体现出小麦株高性状是一个多基因控制的复杂数量性状,影响株高的因素还需要更加深入的探讨。结合TaGAMyb-B在基部第二节间相对表达量TaGAMyb-Ba类型材料的转录本表达量均高于TaGAMyb-Bb类型材料,证明TaGAMyb-B是影响株高建成的主效基因之一。这一研究结果将会在重组自交群体(两亲本株高差异大,且分别属于TaGAMyb-Ba和TaGAMyb-Bb两种等位变异类型)中进一步进行验证;并且通过转基因的途径来研究TaGAMyb-Ba和TaGAMyb-Bb两种等位变异类型对小麦株高的影响。

图4 两种等位变异类型小麦的第二茎间纵切面
小麦茎秆结构由外及里依次为表皮、厚壁组织、维管束、薄壁组织和髓腔,其中厚壁组织起到支撑茎秆的作用。组织学观察表明,TaGAMyb-Ba和TaGAMyb-Bb两种基因型材料间茎秆第二节间厚壁组织厚度没有明显差异,但厚壁组织的细胞长度存在显著差异,且TaGAMyb-Ba基因型材料细胞长度显著长于TaGAMyb-Bb基因型材料,进一步证明了TaGAMyb-B是影响细胞伸长的一个重要因子,TaGAMyb-Ba对小麦第二节间的细胞伸长具有促进作用,有助于株高的建成,不利于抗倒伏。
与TaGAMyb-Ba相比,等位基因TaGAMyb-Bb在第一内含子中存在一个反向重复的84 bp 插入序列,该序列与小麦中国春的一个脚手架基因(GenBank登录号为HG670306.1)有100%的同源性[23]。反向重复序列在双链DNA中可能引起十字形结构的形成,存在于某些转座子末端,发挥重要作用[25]。对转座子的研究表明,不同的转座子产生不同的小RNA,对不同基因的表达进行调控[26]。结合茎秆第二节间纵切结果,推测TaGAMyb-Bb等位基因相对于TaGAMyb-Ba等位基因来说,第一内含子中存在的84 bp插入可能扮演转座子的角色,产生小RNA来调节细胞伸长相关基因的表达,从而造成细胞间长度的差异,影响株高。
