“现在”有多长:空时距知觉的分段性研究
2018-06-06于晓琳,黄希庭,陈有国,邱俊杰
于 晓 琳,黄 希 庭,陈 有 国,邱 俊 杰
(西南大学 心理学部,重庆市 400715)
一、引 言
在日常生活中,人们总会不假思索地使用“现在”这个词,但“现在”的定义是什么?它只是过去与将来的分界线,还是有其自身的时间长度?这引发了哲学家和心理学家们的思考。波佩尔认为:“‘现在’——也就是我们的意识——犹如架在时间这匹骏马上的鞍子,我们坐在上面,像两面人似得前后张望:向着过去,也向着将来。”[1]这种认为“现在”本身具有时间长度的观点得到了普遍认同。那么心理的现在(psychological now)究竟有多久?该问题与时间知觉具有相同的内涵[2]。
时间知觉(time perception)是对客观现象的持续性和顺序性的感知[3],与工作记忆密切相关。工作记忆(working memory)作为唯一可进行有意识加工的记忆阶段,负责暂时保持和处理包括字母、数字、图形、面孔和空间位置等多种类型的信息[4]。时间作为一种特殊的信息,也可在工作记忆中得到暂时保持,这就涉及到“现在”的上限。因为,工作记忆的容量十分有限,从最早发现平均容量为7±2[5]到后来被修正为4[6],都表明工作记忆只能保持一定限度的时间长度,即为“现在”的上限。Fraisse对相关研究进行了总结,认为心理的现在不超过5s,平均在2~3s[7]。波佩尔也提出,只有在约3s的时间范围内,信息才能被整合为一个单元或时间上的“格式塔”,成为心理的现在的内容[1]。可见,该限度以内的一段时间可被人们知觉为现在,超过该限度则被排除在现在之外,成为过去。这导致处于该分界线上下的时间知觉存在分段性,可能对应着不同的时间加工方式。
然而,“现在”的限度并非一成不变,该限度会受哪些因素影响越来越引起研究者们的关注。早期研究虽对此进行了一些有意义的尝试,但限于行为研究的局限性,难以深入到内部表征进行探讨。近来,研究者发现大脑神经振荡活动对维持工作记忆的表征起重要作用[8-10],为该问题提供了新途径。Chen等[11]和Yu等[12]先后采用脑电图(EEG)技术对视、听时距在工作记忆保持过程中的神经振荡特点进行了考察,首次从电生理角度表明“现在”的限度受到感觉通道的影响。其中,人们从视觉通道体验的“现在”的上限是3s[11],听觉通道则为2s[12]。对于该发现,研究者认为其原因是人们对来自不同通道的时距知觉本身存在差异,即听觉信号总是比客观等长的视觉信号知觉为更长,该通道效应在以往研究中被多次证实[13-14]。由此,容量有限的工作记忆所能保持的听觉时距比视觉更短,导致分界线更短。值得注意的是,有研究发现,在时距知觉中,比上述视听知觉差异更明显的效应发生在不同时距类型之间[15],这意味着以不同时距类型呈现的“现在”限度很可能也具有差异,这是本研究主要关注的问题。
时距分为两种类型:以持续形式呈现的为实时距(filled interval),以间隔形式呈现的为空时距(empty interval),即分别由一个简短的感觉事件来标记这段时间的开始和结束,而在这段时间过程中并没有信号呈现的“安静的”时距[16]。与时距知觉的视听通道效应类似,研究者发现对于客观相等的一段时距,实时距会比用两个短音标记的空时距知觉为更长[17],称为实时距错觉(filled-duration illusion)。例如,Zwicker要求被试自行调整一段连续的声音长度使之达到与一段空时距的长度相等,发现人们会感到一段实时距和一段客观上约两倍长于该实时距的空时距等长[18]。Craig也发现,空时距需要比实时距长657 ms才能被知觉为与实时距相等[19]。Wearden等人的研究也表明,一段空时距的长度仅仅被知觉为客观等长的一段实时距的55%~65%,且该错觉随刺激时距的增加越发明显[15]。研究者主要采用时间知觉的内部时钟模型(internal clock model)[20]对实时距错觉进行解释。该模型认为,人脑中存在一个起搏器(pacemaker)和累加器(accumulator),二者之间有开关(switch)相连。当个体开始计时时,起搏器以一定频率发射脉冲,开关闭合,脉冲进入累加器进行整合,形成时间表征。研究者认为,实时距错觉的产生原因可能是与空时距相比,实时距知觉过程中起搏器发射脉冲的速度更快,或开关关闭更快而打开更慢,这都使实时距累加的脉冲更多,进而知觉为更长[15,21]。有趣的是,这与视听时距知觉的通道效应的解释非常相似。那么空时距在工作记忆中的神经表征是否也与实时距存在差异,并且能否用与视听时距相同的方式进行解释,这是本研究所要探讨的问题。
考虑到Yu等[12]已对听觉实时距进行过探讨,本研究采用与以往研究[11-12]相同的matching-to-sample任务,结合脑电图技术对听觉空时距在工作记忆保持过程中的神经振荡特点(theta和alpha波)进行考察,假设如下:首先,“现在”的限度会受到不同时距类型的影响,即空时距在工作记忆中的表征分界线与实时距不同;其次,若引起实、空时距表征分界线差异的原因与视听时距的原因相同,那么与实时距的表征分界线相比(2s)[12],空时距的表征分界线将更长,即体验到的“现在”的上限会长于2s。
二、研究方法
(一)被试
23名本科生自愿参加本实验,其中男生11名,女生12名,年龄为18~26岁(平均年龄20.304±2.055岁)。均为右利手,听力正常,视力或矫正视力正常,没有脑部损伤或神经系统疾病。实验后给予适当报酬。
(二)材料和仪器
听觉空时距刺激采用两个持续50 ms的纯音(1 000Hz,响度适宜约80dB,各有10 ms的淡入淡出)标记时距的开始和结束。时距长度分别为1s、2s、3s、4s。空时距的长度界定为第一个声音消失到第二个声音刚出现之间的时间间隔,声音本身的持续时间不计入时距[22]。由Sennheiser耳机双耳呈现。反应信号为视觉呈现一个白色问号,大小为2cm(1.53°)。电脑距被试约75cm。屏幕刷新频率为85Hz。
(三)实验程序
采用matching-to-sample任务,整个实验共有7个blocks,每个block包括48个trials。每种时距条件包括84个trials。如图1所示,在每个trial中,先后呈现两个听觉空时距刺激(sample刺激和probe刺激),中间间隔3s(保持阶段)。每个空时距刺激的呈现时间为4种时距(1~4s)中随机一种。经过1s间隔后,在屏幕中央出现一个问号(反应信号),随按键消失或经过2s后自动消失。被试需要估计第二个空时距刺激(probe)的时距是短于、等于或长于第一个空时距刺激(sample)。在反应阶段,被试分别使用食指、中指或无名指按键盘上的1、2或3键(1~3键依次表示probe刺激短于、等于或长于sample刺激)。其中,一半被试用左手反应,另一半用右手反应。除休息时间外,要求被试在实验过程不能闭眼,始终注视屏幕中央的注视点。trial之间间隔2s。
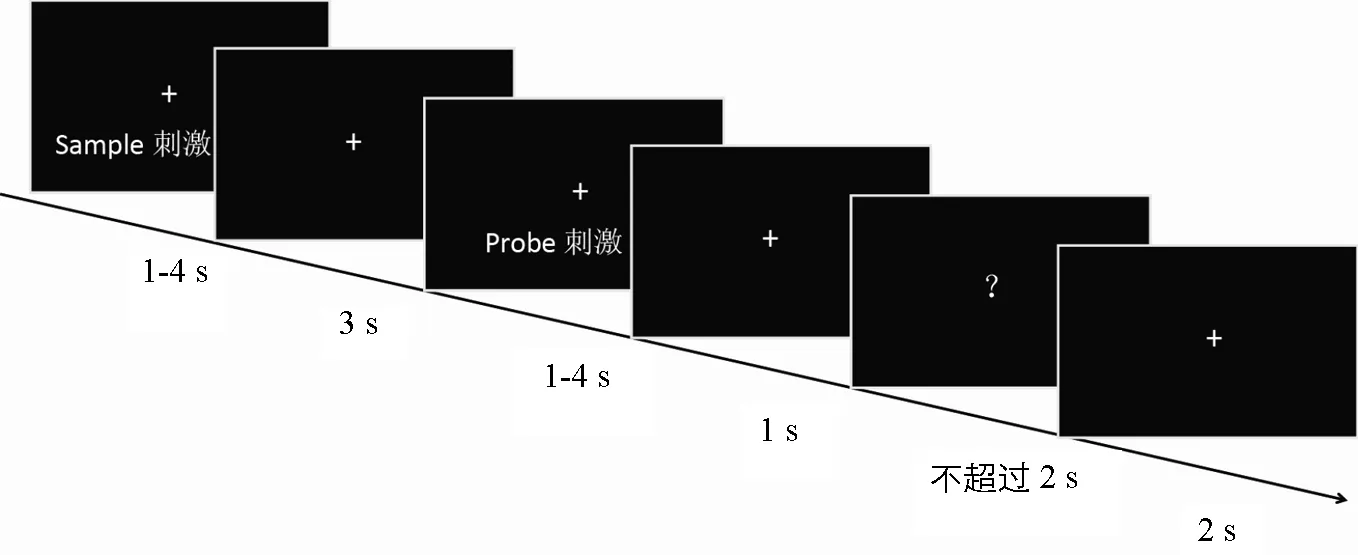
图1 单个trial的时距呈现流程图
(四)脑电记录
采用安置Ag/AgCl电极的电极帽(Brain Products GmbH, Herrsching, Germany)连续记录EEG。按照国际扩展10-20系统安置64导电极记录脑电。另外增加两个电极分别安置于左右乳突。左眼外侧安置电极记录水平眼电(HEOG),右眼下方安置电极记录垂直眼电(VEOG)。采样速度为500 Hz。头皮电阻均小于5 KΩ。
采用EEGLAB[23]和MATLAB(The MathWorks, Inc., Natick, Massachusetts, USA)进行离线分析。连续的EEG数据均采用左右耳后乳突的平均值进行重参考,数字滤波为0.5 Hz高通[11,24-25]。以sample刺激(第一个声音)开始为时间点,对连续数据以-1s到8s的窗口进行分段。每段EEG通过减去sample刺激开始前的平均电压进行基线矫正。波幅超过±100 μV自动标记,经观察确认后手动删除。采用独立成分分析,并根据头皮地图和活动特点[26-27]辨别和删除与眼动等有关的伪迹独立成分。共保留95.76%的trials。重复测量方差分析发现,四种时距条件保留的trials数量无显著差异[F(3,66)=0.427,p>0.05,ηp2=0.019]。
对已分段且无伪迹的数据进行时频分析。采用EEGLAB探讨EEG节律的trial-by-trial事件相关谱功率调制。采用3个周期、频率为3 Hz的汉宁窗正弦小波进行时频分析,直至线性上升为15个周期、频率为30Hz[28]。采用事件相关谱扰动(event-related spectral perturbation, ERSP)指数计算事件相关谱功率反应的变化[29](dB转换以sample刺激开始前-0.4s至-0.1s为基线)。图2为sample刺激的编码、保持阶段以及probe刺激呈现阶段的ERSP,均通过减去sample刺激开始前-0.4s至-0.1s的平均振荡功率进行校正。

图2 1s(A),2s(B),3s(C),4s(D)空时距条件在大脑后部的ERSP总平均图
如图3所示,经频率平均后,从保持阶段开始后1s至3s期间提取出稳定的theta(4-8Hz)和alpha波(8-12Hz)活动。在新坐标系统中,0代表保持阶段的开始。本研究根据Roberts等[25]的研究将电极点分为九个区域,即左-额叶((Fp1,AF3,AF7,F3,F5),中-额叶(Fpz,F1,Fz,F2),右-额叶(Fp2,AF4,AF8,F4,F6),左-中央(FC5,C3,C5,T7,CP5),中-中央(FCz,Cz,C1,C2,CPz),右-中央(FC6,C4,C6,T8,CP6),左-后部(P3,P5,P7,PO3,O1),中-后部(P1,P2,Pz,POz,Oz),右-后部(P4,P6,P8,PO4,O2)。
由于基线和保持阶段之间的时间间隔在四种时距条件下并不相同,为评估统计分析是否受基线和统计时间窗口之间不同时间间隔的污染,额外对所选取的保持阶段开始前-0.5s至-0.1s的时间窗口进行统计。以时距条件(1s,2s,3s,4s)和区域(九组)为自变量,分别对-0.5s至-0.1s和1s至3s的时间间隔的theta和alpha波进行重复测量方差分析。当不符合球形检验时,均采用Greenhouse-Geisser校正方法进行校正[30]。偏eta方(ηp2)用来估计ANOVA的效应量[31]。
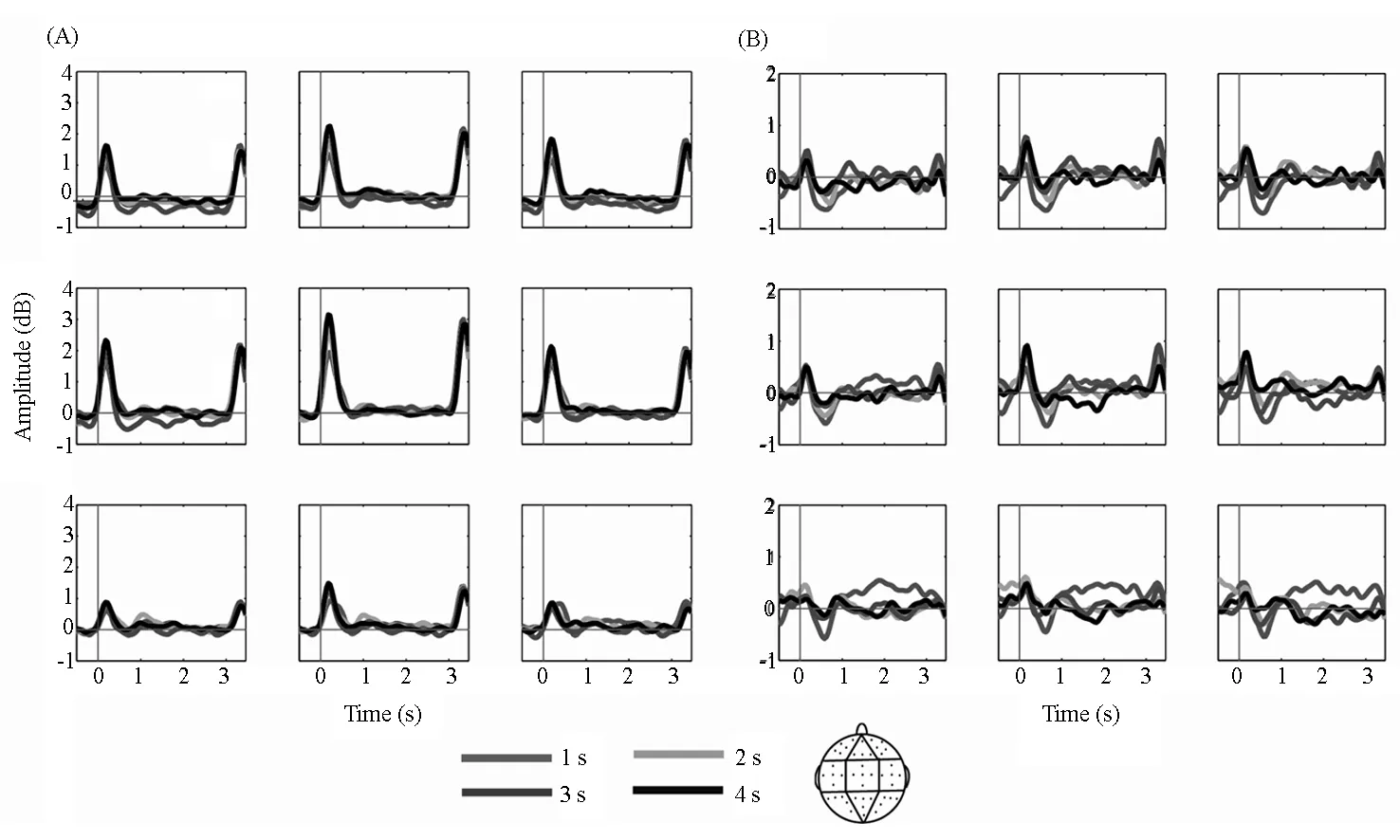
图3 1s,2s,3s,4s空时距条件在保持阶段中的theta波(A)和alpha波(B)平均值
(一)行为结果
以时距条件(1s,2s,3s,4s)为自变量,正确率为因变量进行重复测量方差分析,发现时距条件主效应显著(F(3,66)=50.616,p<0.001,ηp2=0.697)。事后检验发现,1s时距条件的正确率(M±SE:0.867±0.019)显著高于2s(0.783±0.019)、3s(0.708±0.018)和4s(0.628±0.027)条件(ps<0.001),2s时距条件的正确率显著高于3s和4s条件(ps<0.001),3s时距条件的正确率显著高于4s条件(p<0.01)。
以时距条件(1s,2s,3s,4s)为自变量,反应时为因变量进行重复测量方差分析,发现时距条件主效应显著(F(1.779,39.136)=4.308,p<0.05,ηp2=0.164)。事后检验发现,1s时距条件的反应时(495.169±29.163)与2s(508.757±31.672)无显著差异(p>0.05),显著小于3s(522.573±32.861)和4s(524.135±32.396)条件(ps<0.05)。2s时距条件的反应时显著小于3s和4s条件(ps<0.05),3s和4s条件的反应时无显著差异(p>0.05)。
(二)EEG结果
对保持阶段开始前0.5~0.1s内theta波进行重复测量方差分析发现,时距的主效应、时距与脑区的交互作用均不显著(ps>0.05)。对保持阶段开始后1~3s内theta波进行重复测量方差分析发现,时距的主效应不显著[F(3,66)=0.623,p>0.05,ηp2=0.028],脑区的主效应显著[F(2.014,44.298)=3.219,p<0.05,ηp2=0.128],表现为在大脑后部(左-后部(0.078±0.123 dB)、中-后部(0.098±0.114 dB)和右-后部(0.111±0.109 dB)激活相对较大。交互作用的主效应不显著[F(7.140,157.083=1.010,p>0.05,ηp2=0.044]。
对保持阶段开始前0.5~0.1s内alpha波进行重复测量方差分析发现,时距的主效应、时距与脑区的交互作用均不显著(ps>0.05)。对保持阶段开始后1~3s内alpha波进行重复测量方差分析发现,时距的主效应显著[F(3,66)=2.908,p<0.05,ηp2=0.117],表现为1s(0.353±0.225 dB)条件下的alpha波值显著高于2s(0.011±0.220 dB)、3s(-0.005±0.193 dB)和4s(-0.036±0.208 dB)条件(t(22)=2.113—2.641,ps<0.05),而2s、3s和4s条件之间则无显著差异(t(22)=0.106-0.330,ps>0.05)。脑区条件的主效应不显著[F(2.194,48.269)=1.228,p>0.05,ηp2=0.053],时距与脑区的交互作用显著[F(4.898,107.753)=5.166,p<0.01,ηp2=0.190]。进一步简单效应分析表明,时距效应主要在后部三区(左-后部、中-后部和右-后部)上显著[F(3,66)=7.296-7.529,ps<0.01,ηp2=0.249-0.255],而在大脑前部三区和中部三区则均不显著[F(3,66)=0.148-1.227,ps>0.05,ηp2=0.007—0.053]。
如图4所示,在保持阶段开始后1~3s的时间间隔中,theta和alpha波功率在大脑后部功率最大。
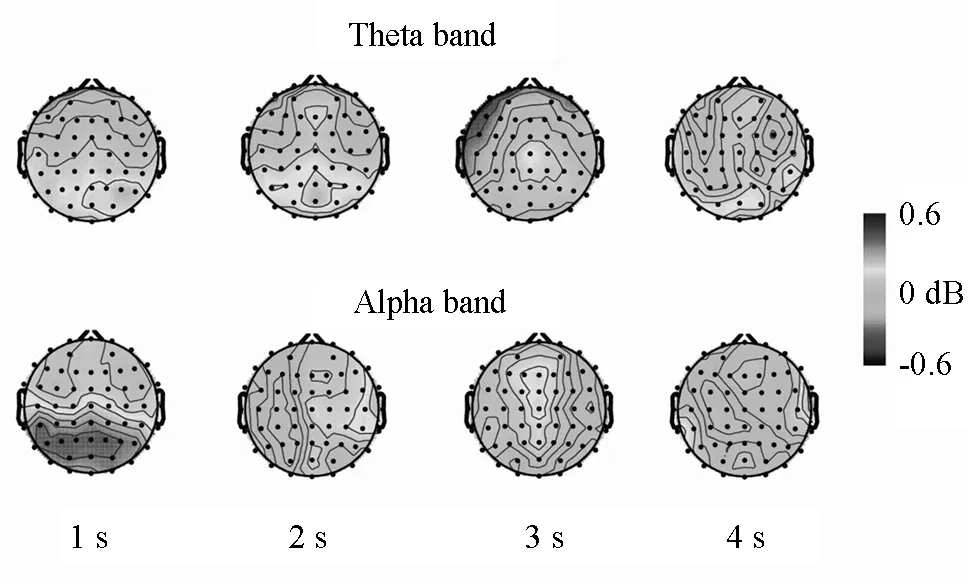
图4 1s,2s,3s,4s时距条件在保持阶段中的theta和alpha波振荡活动的地形图
EEG结果表明,在工作记忆的保持阶段,2s、3s和4s时距条件的alpha波波幅均显著低于1s条件,不同时距条件的theta波波幅却无显著差异,这与以往研究一致[11-12],均发现不同时距条件在alpha波而非theta波上表现出分段性。同时,与实时距的表征分界线相比(2s)[12],本研究发现空时距表征的分界线为1s,这支持本研究的假设一,即空时距在工作记忆中的表征分界线与实时距不同,说明“现在”的限度会受到不同时距类型的影响。然而,空时距的表征分界线短于实时距[12]与假设二不符,说明引起实、空时距表征分界线差异的原因很可能与视听时距分界线差异的原因不同。
如引言所述,时距知觉的视听通道效应与实时距错觉分别描述的是不同通道和不同类型的信息在时距知觉上具有差异的现象。以往研究者倾向采用相似的原因对二者进行解释,但多停留在理论层面,缺少实验证明。本研究首次对空时距的神经表征进行探讨,结果发现空时距的表征分界线为1s,说明工作记忆最多只能保持1s长度的空时距,即体验的“现在”的上限是1s,短于实时距[12],这不能用实时距错觉进行解释。波佩尔认为,“现在”经验的形成有赖于大脑的一种时间整合机制。该整合机制将系列事件融合成知觉单元,一旦两事件间的时间间隔超过一定限度(“现在”的时间长度),该整合单元就会解体[1]。我们认为,实、空时距间的差异不仅仅体现在知觉长度上,二者的呈现方式也具明显差异:与实时距的持续呈现相比,人们更容易将标记空时距开始和结束的两个刺激知觉为两个独立的事件,而非一个整体。这可能使空时距在工作记忆中被保持为一个整体单元的难度更大,最终导致空时距表征分界线更短。研究发现,6岁儿童可与成人一样较准确的复制出1s的实时距,但很难复制出空时距,即只能复制出空时距的其中一个声音标记,或复制出两个快速连续的声音标记。研究者认为,他们很可能只能觉察到空时距两声音标记的相继性(即知觉为两个独立的事件),而难以知觉二者之间标记的时距长度(即整合为一个主观结构)[7]。在本研究中,当两声音标记间隔在1s以内时,人们很容易将先后的两标记整合为一个主观结构,作为一个整体保持在工作记忆中。然而一旦刺激间隔超过1s,人们则无法进行整合,而将两刺激知觉为两个独立的事件,1s即为人们体验到的由空时距形式呈现的“现在”的限度。
本研究从神经表征的层面为理解实、空时距的差异提供了新视角,发现时距类型是影响“现在”的限度的因素之一,也表明不同影响因素之间的内部机制不能一概而论,还需未来研究进行深入探讨。此外,需要说明的是,与EEG结果不同,1~4s时距条件在行为结果上并未表现出分段性。主要原因是由于行为研究的结果会综合受到时距信息的编码、保持和决策阶段影响,而EEG结果却能将上述三阶段进行分离,从而单独考察不同时距条件在保持阶段的分段性,后者显然在考察“现在”的限度这一问题上更具优势[11-12]。
参考文献:
[1] 波佩尔.意识的限度:关于时间与意识的新见解[M].李百涵,韩力,译.北京:北京大学出版社,1995:47.
[2] 黄希庭.心理学导论[M].北京:人民教育出版社,2015:250.
[3] 凤四海,黄希庭.时间知觉理论和实验范型[J].心理科学,2004(5):1157-1160.
[4] BADDELEY A. Working memory[J]. Science, 1992, 255(5044):556-559.
[5] MILLER G A. The magic number seven, plus or minus two: Some limits on our capacity for processing of information[J]. Psychological review, 1956, 63(2): 81-93.
[6] COWAN N. The magical number 4 in short-term memory: A reconsideration of mental storage capacity[J]. Behavioral and brain sciences, 2001, 24(1): 87-185.
[7] FRAISSE P. Cognition of time in human activity. Cognition in motivation and learning[M]. Hillsdale, New York: Lawrence erlbaum, 1981:233-258.
[8] JENSEN O. Maintenance of multiple working memory items by temporal segmentation[J]. Neuroscience, 2006, 139(1): 237-249.
[9] KOENE R A, HASSELMO M E. First-in-first-out item replacement in a model of short-term memory based on persistent spiking[J]. Cerebral cortex, 2007, 17(8): 1766-1781.
[10] LISMAN J E, IDIART M A. Storage of 7 plus/minus 2 short-term memories in oscillatory subcycles[J]. Science, 1995, 267(5203): 1512-1515.
[11] CHEN Y G, CHEN X, KUANG C W, HUANG X T. Neural oscillatory correlates of duration maintenance in working memory[J]. Neuroscience, 2015, 290: 389-397.
[12] YU X L, CHEN Y G, QIU J J, HUANG X T. Neural oscillations associated with auditory duration maintenance in working memory[J]. Scientific reports, 7(1), 5695-5703.
[13] PENNEY T B, GIBBON J, MECK W H. Differential effects of auditory and visual signals on clock speed and temporal memory[J]. Journal of experimental psychology: human perception and performance, 2000, 26(6): 1770-1789.
[14] WEARDEN J H, TODD N, JONES L. When do auditory/visual differences in duration judgements occur?[J]. The quarterly journal of experimental psychology, 2006, 59(10):1709-1724.
[15] WEARDEN J H, NORTON R, MARTIN S, MONTFORD-BEBB O. Internal clock processes and the filled-duration illusion[J]. Journal of experimental psychology: human perception & performance, 2007, 33(3):716-729.
[16] 郭秀艳,聂晶.大学生实时距、空时距估计的比较研究[J].心理发展与教育,2003(3):58-62.
[17] HOOPEN G T, MIYAUCHI R, NAKAJIMA Y. Time-based illusions in the auditory mode. In GRONDIN S (Ed.), Psychology of time[M]. Bingley, UK: Emerald, 2008:139-187.
[18] ZWICKER E. Subjective and objective duration of sound impulses and sound pauses[J]. Acustica, 1970, 22(4):214-218.
[19] CRAIG J C. A constant error in the perception of brief temporal intervals[J]. Attention, perception and psychophysics, 1973, 13(1):99-104.
[20] GIBBON J, CHURCH R M. Sources of variance in an information processing theory of timing. In ROITBLAT H L, et al. (Eds.), Animal cognition[M]. Hillsdale, NJ: Erlbaum, 1984:465-488.
[21] HASUO E, NAKAJIMA Y, TOMIMATSU E, GRONDIN S, UEDA K. The occurrence of the filled duration illusion: a comparison of the method of adjustment with the method of magnitude estimation[J]. Acta psychologica, 2014, 147(2):111-121.
[22] TSE C Y, PENNEY T B. Preattentive timing of empty intervals is from marker offset to onset.[J]. Psychophysiology, 2006, 43(2):172-179.
[23] DELORME A, MAKEIG S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis[J]. Journal of neuroscience methods, 2004, 134(1): 9-21.
[24] HSIEH L T, EKSTROM A D, RANGANATH C. Neural oscillations associated with item and temporal order maintenance in working memory[J]. Journal of neuroscience, 2011, 31(30): 10803-10810.
[25] ROBERTS B M, HSIEH L T, RANGANATH C. Oscillatory activity during maintenance of spatial and temporal information in working memory[J]. Neuropsychologia, 2013, 51(2): 349-357.
[26] JUNG T P, MAKEIG S, HUMPHRIES C, LEE T W, MCKEOWN M J, IRAGUI V, SEJNOWSKI T J. Removing electroencephalographic artifacts by blind source separation[J]. Psychophysiology, 2000, 37(2):163-178.
[27] JUNG T P, MAKEIG S, WESTERFIELD M, TOWNSEND J, COURCHESNE E, SEJNOWSKI T J. Removal of eye activity artifacts from visual event-related potentials in normal and clinical subjects[J]. Clinical neurophysiology,2000, 111(10):1745-1758.
[28] MAKERIG S, DELORME A, WESTERFIELD M, JUNG T P, TOWNSEND J, COURCHESNE E, SEJNOWSKI T J. Electroencephalographic brain dynamics following manually responded visual targets[J]. Plos biology, 2004, 2(6):e176.
[29] MAKERIG S. Auditory event-related dynamics of the EEG spectrum and effects of exposure to tones[J]. Electroencephalography and clinical neurophysiology, 1993,86(4):283-293.
[30] GREENHOUSE S W, GEISSER S. On methods in the analysis of profile data[J]. Psychometrika, 1959, 24(2):95-112.
[31] LEVINE T R, HULLETT C R. Eta squared, partial eta squared, and misreporting of effect size in communication research[J]. Human communication research, 2002, 28(4):612-625.
