不同硒水平对鲈鱼生长性能及抗氧化能力的影响
2018-05-16郭正富杨小琴
郭正富,李 军,杨小琴
(1.四川宜宾职业技术学院,四川宜宾 644003;2.四川省宜宾市高县水务局,四川宜宾 645150)
硒(Se)是动物维持生命代谢的必需微量元素,是生命体完成清除自身体内不同过氧化物的重要物质(Flatman,1990);同时它在生命体中起到防止细胞线粒体脂类过氧化,避免脂类代谢物对细胞膜的破坏。目前大量的研究结果表明,在饲料中添加硒盐在一定程度上对动物机体生长性能和抗氧化能力产生影响(刘康,2010;朱春峰,2009);如石斑鱼(Lin,2005)、大黄鱼(曹娟娟等,2015)、军曹鱼(Liu等,2010)、鲈鱼(梁萌青等,2006)等水产品种,也在鲤鱼(Ashouri等,2015)、大口黑鲈(Zhu等,2012)和革胡子鲶(Abdel等,2007)等水产品种进行硒对脂肪含量及血清脂肪代谢影响的试验。
鲈鱼(Lateolabrax japonicus)是一种广温性鱼类(适宜水温3~29 ℃),在我国南方和北方海域均有养殖,也是一种广盐性鱼类,近几年是海水养殖发展的重点品种。随着鲈鱼人工养殖技术的成熟和规模的不断扩大,其对于商品饲料的要求升高。目前除谈枫(2014)对中期规格(214.5±1.0)g鲈鱼硒元素需要量研究报道外,国内极少见硒对鲈鱼的影响研究。因此,本试验通过研究饲料中硒水平对鲈鱼营养生理及抗氧化能力的影响,以期为鲈鱼基础营养需求数据库的完善及高效配合饲料的研制提供参考。
1 材料与方法
1.1 试验饲料 在基础饲料中分别添加0、0.3、0.6、0.9、1.2、1.5 mg/kg 的 硒 盐(NaSeSO3),配置成硒水平依次为 0.06、0.37、0.65、0.97、1.28、1.60 mg/kg的6种等氮等脂饲料。试验饲料基础配方及营养组成见表1。试验所需原料均粉碎过80目筛,混合均匀后制成粒径为2 mm的颗粒饲料,60 ℃干燥后于-20℃保存备用。
1.2 试验动物及饲养管理 试验开始前,用0.06 mg/kg组饲料投喂试验鱼2周,分组前饥饿24 h后挑选体态均匀的(12.30±0.04)g鲈鱼720尾随机分组。按每组3个网箱(1.5 m×1.5 m×1 m)、40(尾/箱)的设置平行进行养殖实验60 d,每天饱食投喂2次(8:10和17:10)。试验期间水温稳定在24~29 ℃,溶氧6 mg/L以上。
1.3 样品采集和指标测定 养殖试验结束空腹24 h后将各试验组鲈鱼全部用MS-222(40 mg/L)麻醉后称重计数。然后按10尾/网箱的随机抽取各组试验鱼,尾静脉取血置于抗凝管中,4 ℃下5000 r/min离心8 min后,取上层血清48 h内完成检测。按5尾/网箱选取试验鱼,解剖取其肝脏保存于防冻管内,液氮急冻后保存在-80 ℃超低温冰箱中,一周内完成相关指标的检测。

表1 基础饲料原料组成及营养水平(干重基础) %
饲料及全鱼常规营养成分测定参照胡静等(2016)方法。
肝脏和血清谷胱甘肽还原酶(GR);谷胱甘肽过氧化物酶(GSH-Px);总超氧化物歧化酶(T-SOD);丙二醛(MDA)采用试剂盒(南京建成生物工程研究所)测定。
1.4 生产性能 生产性能计算公式为:
成活率/%=试验期末鱼尾数/试验期初鱼尾数×100;
增重率/%=(试验期末均重-试验期初均重)/试验期初均重×100;
特定生长率/(%/d)=(ln试验期末均重-ln试验期初均重)/试验天数×100;
饲料效率/%=(试验期末总重+死亡鱼总重-试验期初总重)/总摄入饲料量×100。
1.5 统计分析 数据全部采用“平均值±标准误”表示,用SPSS17.0进行单因子方差分析和多重比较。
2 结果
2.1 饲料不同硒含量对鲈鱼幼鱼生长性能的影响 不同硒含量饲料对鲈鱼幼鱼相关生长指标影响见表2,0.97 mg/kg组的鲈鱼幼鱼增重率(657.86±2.83)% 和特定生长率(3.38±0.11)%/d达到最高,显著高于除0.65 mg/kg组以外的其他各组(P<0.05),饲料效率达到(20.01±0.11)%,变化趋势也与增重率相似(P<0.05);各组鲈鱼的存活率均保持为100%(P>0.05)。通过二次回归分析增重率和特定生长率数据(杜强等,2011)得出,当饲料中硒含量约为0.81 mg/kg时,鲈鱼幼鱼能达到最大生长性能(图1和图2)。
2.2 不同硒含量饲料对鲈鱼幼鱼体组成的影响0.97 mg/kg组鲈鱼幼鱼粗蛋白质含量最高,显著高于0.06和0.65 mg/kg组(P<0.05)。0.06 mg/kg组鲈鱼幼鱼的全鱼粗脂肪含量显著低于0.37、1.28和 1.60 mg/kg组(P< 0.05),而鲈鱼幼鱼的全鱼水分在0.06 mg/kg组高于其他各组,且与0.37、0.97、1.28、1.60 mg/kg组差异达显著水平(P< 0.05)(表 3)。
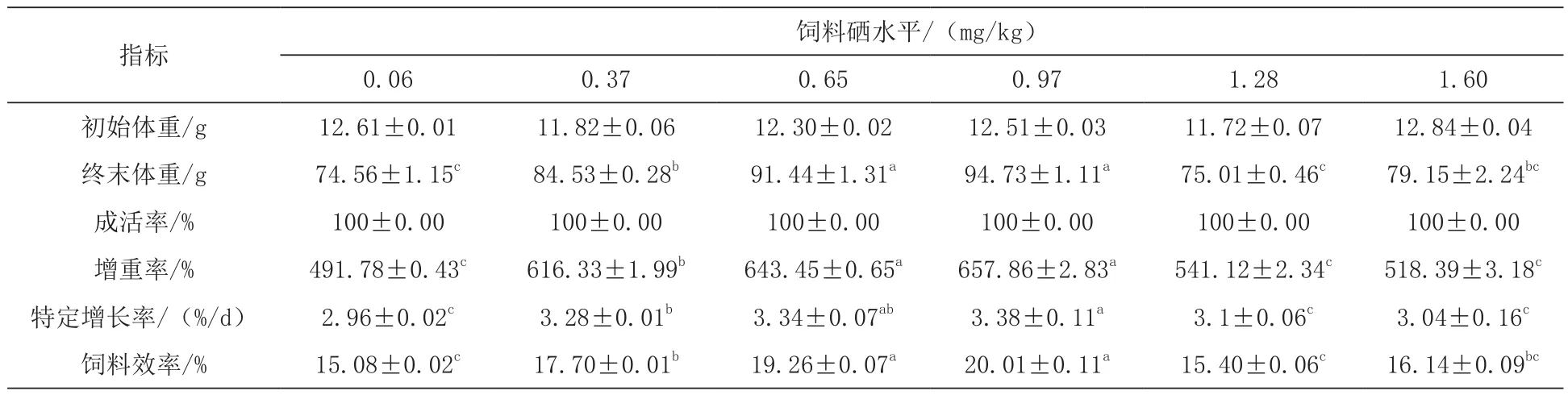
表2 饲料不同硒含量对鲈鱼幼鱼相关生长指标影响
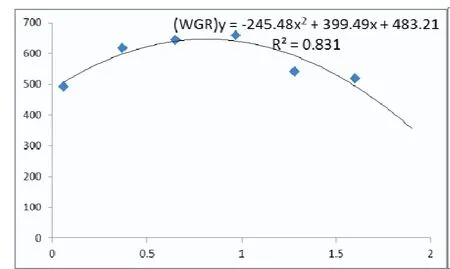
图1 不同硒含量对鲈鱼增重率的影响

图2 不同硒含量对鲈鱼特定生长率的影响
2.3 不同硒含量饲料对鲈鱼幼鱼肝脏抗氧化指标的影响 由表4可知,饲喂含硒饲料60 d后,随着饲料硒含量的增加,试验鲈鱼肝脏的谷胱甘肽过氧化物酶活性升高,1.60 mg/kg组显著高于其他各组(P<0.05);肝脏超氧化物歧化酶活性出现先升后降再升高的趋势,在0.96 mg/kg组达到最大活性,并显著高于其他各组(P<0.05)。各试验组鲈鱼幼体的肝脏丙二醛含量差异不显著(P > 0.05)。
3 讨论
动物对营养的需要量均存在一定的适宜区间,对于硒元素也不例外,适宜区间内,硒对动物具有积极有效的作用,而在区间外会导致硒缺乏症或中毒,这被前人定义为硒的剂量-效应曲线原理(徐辉碧,2014)。通过回归分析文中用不同硒水平饲料饲喂鲈鱼60 d的结果得出,当饲料中硒含量在0.81 mg/kg时,鲈鱼能达到最大生长性能。这与以往对军曹鱼(0.788 mg/kg)的研究结果接近(Liu等,2010),高于对鲈鱼(0.40~ 0.63 mg/kg)(谈 枫,2014;梁 萌 青 等,2006)、草 鱼(0.438~ 0.631 mg/kg)(苏 传 福,2008)、鲤 鱼(0.434 ~ 0.517 mg/kg)(金明昌,2007)、斑点叉尾鮰(0.25 mg/kg)(Gatlin,1984)等的研究结果,但低于军曹鱼(1.29和1.46 mg/kg)(杨原志等,2016)、鲍鱼(1.41 mg/kg)(Wang等,2012)的研究结果。原因可能与水产动物品种及其不同生长阶段、养殖环境和饲料加工工艺等因素有关。同时鲈鱼和军曹鱼同属鲈形目,具有一定的遗传学关系,推测这可能是导致二者硒需要量接近的主要原因,有待从分子生物学更深一步的研究论证。
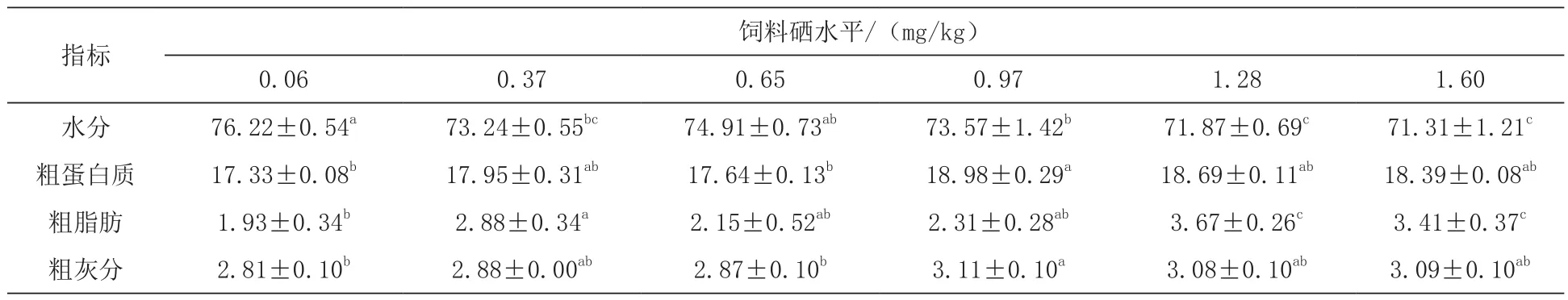
表3 饲喂不同硒含量饲料的鲈鱼幼鱼的营养组成(全鱼) %
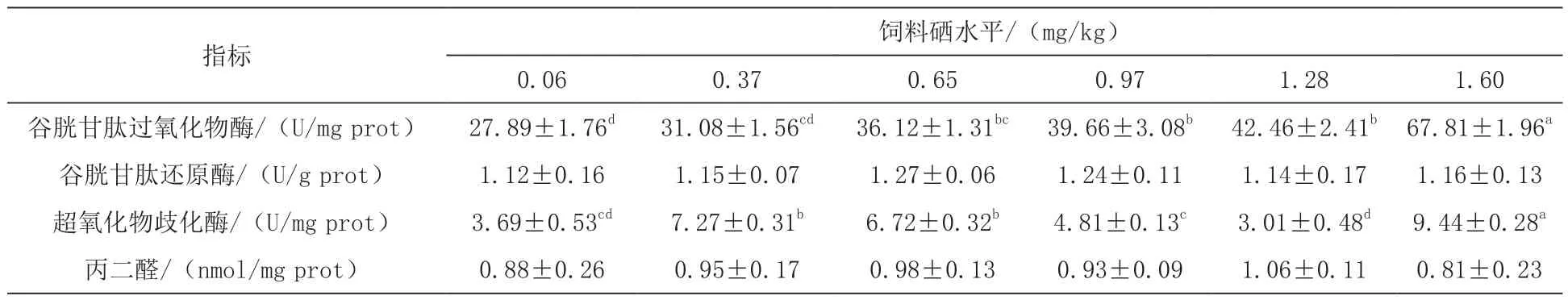
表4 不同硒含量饲料对鲈鱼幼鱼肝脏抗氧化指标的影响
肝脏是鱼类硒蓄积较高的组织(Kim,2014)。肝脏谷胱甘肽过氧化物酶、谷胱甘肽还原酶和超氧化物歧化酶活性及丙二醛含量常用于机体抗氧化能力判断的主要指标(Pacin等,2013)。硒作为谷胱甘肽过氧化物酶的活性中心,肝脏谷胱甘肽过氧化物酶的活性也能反面判断机体硒的效果(胡俊茹等,2016)。本试验中随着硒在动物体内的沉积,谷胱甘肽过氧化物酶活性显著升高,这与以往对石斑鱼(Lin,2005)、黄颡鱼(胡俊茹等,2016)及鲍鱼(Wang等,2012)的研究结果相似。谷胱甘肽过氧化物酶和超氧化物歧化酶的活性及两者间的平衡对于抵抗机体氧化损伤具有重要作用,它们与过氧化氢酶一起被定义为细胞防御功能的首道防线(Gerbhart,2002)。
超氧化物歧化酶是通过捕获超氧阴离子起到对机体氧化免疫防御的作用(Winston,1991)。而铜作为超氧化物歧化酶的结构中心,对于维持该酶的稳定性十分重要,如Lorentzen等(2015)对大西洋鲑研究发现,肝脏硒和铜的蓄积量呈正效应变化。本试验中,肝脏超氧化物歧化酶活性随饲料硒含量增加出现波动变化,推测原因可能是范围内硒能使超氧化物歧化酶活性表现出正效应关系,当硒水平超出一定范围则会抑制超氧化物歧化酶活性,然而当含量过高后,机体再次重启超氧化物歧化酶以降低高硒所引起的氧化应激反应,这一推断有待进一步地验证。
Orun等(2005)研究发现,虹鳟饲料中适量硒盐或有机硒能有效降低鱼体血清和肌肉丙二醛含量(Orun等,2005),这有异于本研究中硒对鲈鱼及西伯利亚鲟鱼(Acipenserbaeri)(Pacini等,2013)和虹鳟(Rider等,2009)中肝脏丙二醛含量的影响。据以往报告推测,出现这一现象的原因可能与试验检测方法有关,目前条件下高效液相色谱法灵敏度远高于试剂盒法(Kucukbay等,2009),这有待技术进一步的提升控制,但在同一实验模式和检测方法条件下,本试验结果仍是鲈鱼幼鱼营养研究较理想参考。
因此,综合本试验结果得出,本试验条件下饲料硒水平为0.81 mg/kg时,鲈鱼可以获得最大增重率和特定生长率。
参考文献
[1]曹娟娟,张文兵,徐玮,等.大黄鱼幼鱼对饲料硒的需求量[J].水生生物学报,2015,39(2):241~ 249.
[2]杜强,林黑着,牛津,等.卵形鲳鲹幼鱼的赖氨酸需求量[J].动物营养学报,2011,23(10):1725~ 1732.
[3]胡静,叶乐,赵旺,等.饲料脂肪水平对眼斑双锯鱼幼鱼生长性能和体成分的影响[J].中国饲料,2016,6:25~28.
[4]胡俊茹,王国霞,孙育平,等.饲料硒含量对黄颡鱼幼鱼生长性能、抗氧化能力和脂肪代谢基因表达的影响[J].动物营养学报,2016,28(12):3925~ 3934.
[5]金明昌.幼鲤硒缺乏症及其机制和硒需要量研究[D].成都:四川农业大学,2007.
[6]梁萌青,王家林,常青,等.饲料中硒的添加水平对鲈鱼生长性能及相关酶活性的影响[J].中国水产科学,2006,13(6):1017~1022.
[7]刘康.军曹鱼幼鱼(Rachycentron canadum)微量元素硒、锰的营养生理研究[D].青岛:中国海洋大学,2010.
[8]苏传福.草鱼幼鱼硒的营养需要研究[D].重庆:西南大学,2008.
[9]谈枫.养殖中期鲈鱼(Lateolabrax japonicus)矿物元素硒、镁的营养需求研究[D].青岛:中国海洋大学,2014.
[10]徐辉碧,黄开勋.硒的化学、生物化学及其在生命科学的应用[M].华中理工大学出版社,1994:25~35.
[11]杨原志,聂家全,谭北平,等.硒源与硒水平对军曹鱼幼鱼生长性能、肝脏和血清抗氧化指标及组织硒含量的影响[J].动物营养学报,2016,28(12):3894~ 3904.
[12]朱春峰.有机硒和无机硒对异育银鲫生长、生理的影响[D].苏州:苏州大学,2009.
[13]Abdel-Tawwab M,Mousa M A A,Abbass F E. Growth performance and physiological response of African catfish,Clarias gariepinus,(B.)fed organic selenium prior to the exposure to environmental copper toxicity [J]. Aquaculture,2007,272(1-4):335~345.
[14]Ashouri S,Keyvanshokooh S,Salati A P,et al. Effects of different levels of dietary selenium nanoparticles on growth performance,muscle composition,blood biochemical profiles and antioxidant status of common carp(Cyprinus carpio)[J]. Aquaculture,2015,446 :25~29.
[15]Flatman P W,Smith L M. Magnesium transport in ferret red cells[J].The Journal of physiology,1990,431(1):11~ 25.
[16]Gerbhart R. Oxidative stress,plant-derived antioxidants and liver fibrosis[J].Planta Medica,2002,68(4):289 ~ 296.
[17]Kim J H,Kang J C. The selenium accumulation and its effect on growth,and haematological parameters in red sea bream,Pagrus major,exposed to waterborne selenium[J].Ecotoxicology and Environmental Safety,2014,104:96~ 102.
[18]Kü·ükbay F Z,Yazlak H,Karaca I,et al. The effects of dietary organic or inorganic selenium in rainbow trout(Oncorhynchus mykiss)under crowding conditions [J].Aquaculture Nutrition,2009,15(6):569~576.
[19]Lin Y H,Shiau S Y. Dietary selenium requirements of juvenile grouper,Epinephelus malabaricus[J].Aquaculture,2005,250(1):356~363.
[20]Liu K,Wang X J,Ai Q H,et al. Dietary selenium requirement for juvenile cobia,Rachycentron canadumL.[J]. Aquaculture Research,2010,41(10):e594~ e601.
[21]Lorentzen M,Maage A,Julshamn K. Supplementing copper to a fish meal based diet fed to Atlantic salmon parr affects liver copper and selenium concentrations [J].Aquaculture Nutrition,2015,4(1):67~72.
[22]Orun I,Ates B,Selamoglu Z,et al. Effects of various sodium selenite concentrations on some biochemical and hematological parameters of rainbow trout(Oncorhynchus mykiss)[J].Fresenius Environmental Bulletin,2005,14(1):18~ 22.
[23]Pacini N,Elia A C,Abete M C,et al. Antioxidant response versus selenium accumulation in the liver and kidney of the Siberian sturgeon(Acipenser baeri)[J].Chemosphere,2013,93(10):2405~2412.
[24]Sebastiena R,Simonj D,Awadhesh J,et al. Supra-nutritional dietary intake of selenite and selenium yeast in normal and stressed rainbow trout(Oncorhynchus mykiss):implications on selenium status and healthresponses[J].Aquaculture,2009,295(3):282~291.
[25]Wang W F,Mai K S,Zhang W B,et al. Dietary selenium requirement and its toxicity in juvenile abalone Haliotis discus hannai Ino[J].Aquaculture,2012,330-333:42~ 46.
[26]Winston G W,Giulio R T D. Prooxidant and antioxidant mechanisms in aquatic organisms[J]. Aquatic Toxicology,1991,19(2):137~161.
[27]Zhu Y,Chen Y,Liu Y,et al. Effect of dietary selenium level on growth performance,body composition and hepatic glutathione peroxidase activities of largemouth bass Micropterus salmoide[J].Aquaculture Research,2012,43(11):1660~ 1668.
