以色列野生二棱大麦籽粒氮素含量的基因型及其生态差异
2018-05-11程晓彬阮景军程剑平NEVOEviatar
程晓彬,李 念,阮景军,赵 钢,程剑平,NEVO Eviatar,严 俊
(1.四川民族学院环境与生命科学系,四川康定 626001; 2.贵州大学麦作研究中心,贵州贵阳 550025;3.成都大学药学与生物工程学院,四川成都 610106; 4.海法大学进化研究所,以色列海法 31905)
大麦是世界上最古老的作物之一,具有早熟、生育期短、适应性广、丰产以及营养丰富等特性,分为栽培大麦和野生大麦2种。大麦因兼具食用、饲用、酿造用以及医药等多种用途而受到人们越来越多的重视[1]。大麦育种多以高产为主要目的,在人工选择过程中容易失去其长期进化产生的遗传多样性,而遗传多样性的缺乏和丧失会导致大麦对环境及病虫害的抗性减弱[2-3]。野生二棱大麦[Hordeumspontancum(Aberg) Shao]受生态环境的影响,在长期自然选择过程中积累了大量抗病、抗虫、耐寒、耐旱、耐贫瘠、耐盐等多种抗逆基因[4],是大麦遗传育种、品种改良的重要材料[5]。中东新月沃地(包括以色列、约旦等国)是野生二棱大麦起源地和遗传多样性中心。野生二棱大麦是栽培大麦的祖先,可直接用于杂交育种,也可作为转基因技术育种的目的基因供体[6]。
籽粒蛋白质含量是评价大麦品质优劣的标准之一,提高籽粒蛋白质含量是大麦育种工作的重要目标[7]。对于饲用大麦,通常育种目标是在注重提高品种产量的同时提高其蛋白质含量;而对于酿造大麦,高蛋白会导致啤酒酿造过程中糖化困难,易混浊,但如果麦芽的蛋白质含量过低,其酿造的啤酒也可能因酵母氨基酸含量过低而影响品质,所以培育籽粒中氮素含量高或适中的大麦品种是满足各种市场需求的首要任务。野生大麦丰富的遗传多样性可为该项育种工作提供丰富的遗传资源[7-8]。气候条件对大麦籽粒蛋白质含量也有一定的影响,年份不同,其籽粒蛋白质含量变化较大[9-10]。本研究对采自以色列不同生长环境的9个群体90个野生二棱大麦基因型的有机氮及无机氮营养水平进行分析测定,以期筛选出不同蛋白质含量和氮素含量的大麦基因型,为栽培大麦种质创新研究提供一定的实验基础和理论依据。
1 材料与方法
1.1 材 料
供试种子采自以色列从南到北不同区域,共9个群体90个基因型(表1)。于2012和2013年种于贵州大学麦作农场、2013年种于安顺农科所试验农场和贵州大学园艺农场,共4个处理,分别为:2012贵州大学麦作农场田间(Y2012 Farm)、2013安顺农科所试验农场田间(Y2013 AS)、2013贵州大学麦作农场盆栽(2013 9F)和2013贵州大学园艺农场田间(Y2013 Garden)。贵州大学麦作农场位于106.4°~106.9°E,26.2°~26.6°N,海拔1 100 m;安顺农科所试验农场(Y2013 AS)位于105.9°E,26.3°N,海拔1 395 m。田间试验采用随机区组排列,播种方式为条播,行长1.5 m,行间距0.4 m,一行为同一基因型,每个基因型3次重复,三叶期定苗,每行留长势基本一致的5株苗;盆栽实验每盆装土8.0 kg,一盆一个基因型,每个基因型3次重复,三叶期定苗,每盆留长势一致的3株苗。麦穗成熟后收获,室内自然风干后脱粒,去杂和挑去不完整籽粒,用去离子水将种子洗净烘干后妥善保存,备用。
1.2 分析方法
样品分析在贵州大学麦作研究中心实验室完成,随机取10粒籽粒用万分之一电子天平称量,计算单粒重;按照国标法(GB2905-82)采用KJ 8400凯氏定氮仪测定籽粒总蛋白质含量,蛋白质N素换算系数为6.25;通过UV1000分光光度计采用考马斯蓝染色法测定可溶性蛋白含量。所有测定设3次重复,取其平均值进行数据分析。
1.3 数据分析
采用Microsoft Excel 2007处理数据,变异系数≥25 %的基因型重测,直到<25%为止。采用JMP6.0和SigmaPlot12.0分析数据,通过Tukey HSD检验进行多重比较,P<0.05为差异显著,P<0.01为差异极显著。
表1试验材料
Table1Materialsusedintheexperiment

群体Population样品数量Samplequantity基因型GenotypeGolanHigh20GN1GN2GN3GN4GN6GN7GN9GN10GN11GN12GN18GN19GN20GN21GN22GN25GN26GN28GN29GN30Shlomi20SH1SH3SH4SH6SH7SH9SH10SH12SH13SH15SH16SH17SH18SH19SH20SH21SH22SH23SH24SH25Hermon13HM1HM2HM3HM4HM5HM7HM9HM14HM16HM17HM18HM19HM20Mehola3ME13ME14ME15Atlit1A5Tabigha8T4T5T6T7T8T9T12T14RoshPina14R1R2R3R6R7R10R11R12R13R15R16R17R19R23Gilboa9G3G4(M)G4(R)G8G10G11G19G20G21ECNorth⁃facingslope2NF1NF2
2 结果与分析
2.1 不同环境下籽粒总氮含量及单粒重的差异
经Tukey HSD检验方法分析,Y2012 Farm和Y2013 AS环境下90个野生大麦基因型籽粒总氮及单粒重差异显著(图1),在Y2012 Farm环境下野生大麦籽粒总氮含量显著高于Y2013 AS,Y2013 AS环境下单粒重显著高于Y2012 Farm。由此可见,环境对野生大麦籽粒总氮含量和单粒重影响显著,单粒重变化趋势与籽粒中总氮含量变化相反,说明每粒种子能富集的氮素有一定上限并具有一定的保守性。
2.2 不同环境下籽粒可溶性蛋白质含量的差异
4个环境下野生二棱大麦籽粒可溶性蛋白质存在差异(图2)。在2013AS环境下野生大麦籽粒可溶性蛋白质含量显著高于其他3个环境,Y2013 Garden、2012 Farm和2013 9F三个环境间差异不显著,说明野生二棱大麦籽粒氮素营养积累受种植环境的影响。在Y2013 AS环境下可溶性蛋白质含量最高,但总氮含量相对偏低,而在Y2012 Farm环境下有相反的结果,这对调控大麦库尔巴哈值有一定意义。

不同小写字母表示差异显著(P<0.05)。下同。
Letters above the bars indicate significant differences (P<0.05).The same below.
图1不同环境下籽粒总氮含量及单粒重的差异
Fig.1Differenceofgraintotalnitrogenandsinglegrainweightintwoenvironments
2.3 不同群体氮素相关指标的比较
2.3.1 不同环境下群体总氮含量的比较
对种植于Y2013 AS和Y2012 Farm环境下6个基因型较多的野生二棱大麦群体籽粒含氮量进行分析,结果(图3)显示,2个环境中Tabigha群体平均总氮含量均最高,在Y2012 Farm环境中显著高于Golan High 和 Shlomi 2个群体,在Y2013 AS环境中显著高于Hermon群体。在Y2012 Farm环境下,Golan High群体平均总氮含量最低,但与群体Shlomi、Gilboa和 Hermon无显著差异,仅与Tabigha和RoshPina群体差异显著,此外,Shlomi、Gilboa、 Hermon和RoshPina 4个群体相互间也无显著差异。在Y2013 AS环境下,Hermon群体平均总氮含量最低,但与除Tabigha外的其他5个群体间并无显著差异。由此可以看出,不同群体的野生大麦在氮素营养性状上存在一定的进化差异和保守性。

图2 不同环境下籽粒可溶性蛋白质含量的差异

图3 不同野生大麦群体平均籽粒含氮量的差异
2.3.2 不同环境下群体可溶性蛋白质含量的比较
对6个野生二棱大麦群体籽粒可溶性蛋白质含量进行Tukey HSD检验发现(图4),在Y2013 9F和Y2013 Garden 2个环境下,Rosh Pina群体的可溶性蛋白质含量显著高于Golan High和Shlomi群体。在Y2013 Garden环境中,Gilboa群体的可溶性蛋白质含量显著高于Golan High和Shlomi群体;在Y2012 Farm环境中,Gilboa群体的可溶性蛋白质含量显著高于Golan High和Rosh Pina群体;在Y2013 AS环境中,Gilboa群体显著高于Hermen、Golan High、Tabigha和Shlomi群体。总体来看,Y2012 Farm、Y2013 AS及Y2013 Garden 3个环境中的Gilboa群体的可溶性蛋白质含量最高,均分别显著高于各自环境下的2个以上群体。此外,每个环境下野生二棱大麦籽粒可溶性蛋白质含量在4个群体间均无显著差异。以上结果说明,野生二棱大麦籽粒可溶性蛋白质含量仍具有一定的保守性。
2.4 极端氮素营养基因型的筛选结果
由表2可知,籽粒总氮含量较高的基因型有 T8、 R3、 GR13和 R23,其中 T8最高,为3.42%;可溶性蛋白质含量较高的基因型有 R23、 T8、 G20、 R3和 GR13,其中 T8最高,为10.22%。综合分析可知,籽粒总氮和可溶性蛋白质含量同时都较高的基因型为 T8,均较低的是 HM17。不同基因型野生大麦的平均可溶性蛋白含量和平均总氮含量均不同,变异幅度相差较大,说明野生二棱大麦基因型籽粒可溶性蛋白和总氮含量存在丰富的遗传多样性,基因型 T8和 HM17均可作为改良现代栽培大麦的优异基因材料。
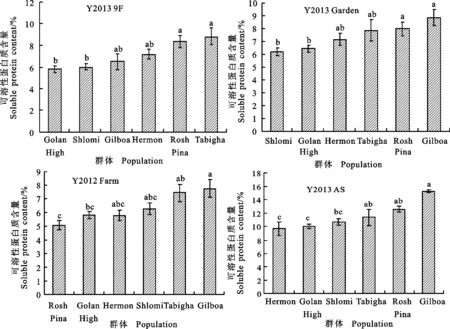
图4 不同野生大麦群体籽粒可溶性蛋白含量的差异
Table2Comparisonofgenotypeswithsignificantgrainnitrogen-relatedindicators

基因型Genetype平均籽粒可溶性蛋白含量Averagegraincontentofsolubleprotein/%平均籽粒总氮含量Averagegraintotalnitrogencontent/%GN267.35d2.01dGN286.10e2.35cR2310.06ab3.04bR39.49c3.12bR199.28c2.96bT810.22a3.42aG209.92b2.93bG219.26c3.06bHM173.23f2.00d
3 讨 论
3.1 不同环境下籽粒总氮含量及可溶性蛋白质含量的差异
从野生大麦群体中筛选极端氮素水平的种质资源用于大麦育种,是提高和改良大麦蛋白质含量的有效方法和途径[11]。本研究中,不同环境籽粒总氮含量和单粒重差异显著,单粒重变化趋势与籽粒中总氮含量变化相反,说明每粒种子富集的氮素具有一定上限并具有一定的保守性。Y2013 AS环境野生大麦籽粒可溶性蛋白质含量显著高于其他3个环境,Y2013 Garden、Y2012 Farm和2013 9F三个环境间的可溶性蛋白质含量差异不显著,说明野生二棱大麦籽粒氮素营养积累既受种植环境的影响,又有一定保守性。本研究选用的种子材料均由中以合作发掘与利用野生麦类资源优异基因课题组(成都大学与海法大学联合成立)采自以色列自然环境,与之前选自以色列海法大学种子库的群体和基因型不同,但仍得到一些相似的结果,尤其在同样种植环境下,个别群体氮素营养显著高于或低于其他群体,说明起源地生态地理环境对籽粒氮素积累有影响,并且籽粒氮素营养在长期的进化和对环境的适应过程中逐步稳定并形成了丰富的遗传多样性[12]。
3.2 不同基因型籽粒总氮、可溶性蛋白质含量和单粒重的比较
大麦高产、高蛋白质含量主要受遗传因素和环境因素的影响[13-14],大麦蛋白质含量属于数量性状遗传且受多个微效基因控制,单个基因效应较小,易受环境条件影响,且在分离世代中表现出连续变异,难以明确分组[10]。因此,在传统的蛋白质遗传研究中,通常将控制同一性状的多基因看作一个整体,运用生物统计的方法计算其均值、方差、显性效应、上位效应、加性效应等参数,用这些遗传参数来描述群体的遗传特征[15]。本研究发现,离散程度较明显的基因型有 GN26、 HM17、 T8、 GR13、 R3和 R23。 GN26和 HM17分别为籽粒总氮和可溶性蛋白质含量较低的基因型; T8为籽粒总氮和可溶性蛋白质含量均较高的个体, R3、 R23和 GR13分别为籽粒总氮和可溶性蛋白质含量较高的基因型。由于研究条件限制,此次筛选出来的基因型个体在某些性状上可能存在不足,测定的相关数据仅供参考,在今后的试验中将进一步扩大种植环境与年份,选用更多起源地具有未知性状的野生大麦种质材料,分析大麦籽粒蛋白质含量的变化特征和氮素积累机制。通过高、低蛋白大麦种质间的杂交构建遗传群体,对籽粒蛋白质含量的遗传特性及控制基因进行分析,尽可能探明影响籽粒蛋白质含量的遗传效应和环境效应的关系,可为选育受环境条件影响小的优质大麦品种提供理论指导。
参考文献:
[1] 卢良恕.中国大麦学[M].北京:中国农业出版杜,1996:1,58,93.
LU L S.Chinese barley science [M].Beijing:China Agriculture Press,1996:1,58,93.
[2] 方桂英,杜向红,张保军,等.约旦野生大麦种质资源形态与产量性状分析 [J].西北农业学报,2010,19(6):82.
FANG G Y,DU X H,ZHANG B J,etal.Analysis of agronomic traits ofHordeumspontaneumC.Koch germplasm from Jordan [J].JournalofNorthwestAgriculturalSciences,2010,19(6):82.
[3] 禹代林,洛桑更堆,欧 珠,等.西藏野生大麦种质资源研究概述 [J].西藏农业科技,1998,20(2):18.
YU D L,LUO S G D,OU Z,etal.A Survey of wild barley germplasm resources in Tibetan [J].TibetanJournalofAgriculturalSciences,1998,20(2):18.
[4] 程晓彬,赵 钢,薛文韬,等.约旦野生二棱大麦在川西高原的物候期和农艺性状分析 [J].西南农业学报,2016,29(5):1027.
CHENG X B,ZHAO G,XUE W T,etal.Analysis of phenophase and agronomic characters of wild barley(Hordeumspontaneum) originated from Jordan in western Sichuan plateau [J].SouthwestChinaJournalofAgriculturalSciences,2016,29(5):1027.
[5] 任喜峰,孙东发.大麦杂交组合蛋白质及部分农艺性状的遗传分析 [J].湖北农业科学,2008,47(4):400.
REN X F,SUN D F.Genetic analysis of protein content and some agronomic traits in annual wild barley from Qing-Tibetan plateau [J].HubeiAgriculturalSciences,2008,47(4):400.
[6] 朱彩梅,张 京.中国糯大麦品种资源及地理分布研究 [J].中国农业科学,2008,41(12):4248.
ZHU C M,ZHANG J.Geographical distribution of waxy barley germplasm resources in China [J].ScientiaAgriculturaSinica,2008,41(12):4248.
[7] 黄志仁,周美学,黄友圣,等.大麦籽粒蛋白质含量的配合力研究 [J].遗传学报,1991,18(3):264.
HUANG Z R,ZHOU M X,HUANG Y S,etal.Combining ability analysis for protein content in barley grain [J].JournalofGenetics,1991,18(3):264.
[8] 张想平,雷耀湖,何庆祥,等.黑糯大麦的品种特性及营养品质分析 [J].种子,2011,30(9):85.
ZHANG X P,LEI Y H,HE Q X,etal.Analysis on characteristic and nutrition quality of black and waxy barley [J].Seed,2011,30(9):85.
[9] 朱彩梅,张 京.应用SSR标记分析中国糯大麦种质的遗传多样性 [J].植物遗传资源报,2010,11(1):63.
ZHU C M,ZHANG J.Genetic diversity analysis of waxy barley in China based on SSR markers [J].JournalofPlantGeneticResources,2010,11(1):63.
[10] 胡立勇,丁艳峰.作物栽培学(第1版)[M].北京:高等教育出版社,2008:574.
HU L Y,DING Y F.Crop cultivation(first edition) [M].Beijing:Higher Education Press,2008:574.
[11] 李 念,阮景军,严 俊,等.以色列野生二棱大麦籽粒多组分营养性状的相关性分析 [J].安徽农业大学学报,2016,43(6):1021.
LI N,RUAN J J,YAN J,etal.Correlation analysis on multiple nutrition traits ofHordeumspontaneumoriginating from different locations in Israel [J].JournalofAnhuiAgriculturalUniversity,2016,43(6):1021.
[12] 文玲玲,薛文韬,严 俊,等.约旦野生二棱大麦群体籽粒休眠特性的网络分析 [J].植物生理学报,2014,50(8):1235.
WEN L L,XUE W T,YAN J,etal.The network analysis on characters of caryopsis dormancy in wild barley (Hordeumspontaneum) populations from Jordan [J].PlantPhysiologyJournal,2014,50 (8):1235.
[13] HAJMANSOOR S,BIHAMTA M R,ALISOLTANI A.Genetic diversity among and within Iranian and non-Iranian barely (HordeumvulgareL.) genotypes using SSR and storage proteins markers [J].BiochemicalSystematicsandEcology,2013,46:15.
[14] 辛培尧,普晓英,杜娟,等.大麦籽粒和苗粉蛋白质含量的检测 [J].麦类作物学报,2016,36(1):60.
XIN P Y,PU X Y,DU J,etal.Protein content determination of Barley grain and seedling powder [J].JournalofTriticeaeCrops,2016,36(1):60.
[15] 汪军妹,张新忠,林 峰,等.73 份优异啤酒大麦种质资源主要麦芽品质性状的评价 [J].浙江农业学报,2012,24(1):4.
WANG J M,ZHANG X Z,LIN F,etal.The assessment of major malt qualities among 73 elite varieties in the germplasm for malting barley [J].ActaAgriculturaeZhejiangensis,2012,24(1):4.
