水分胁迫对不同倍性小麦穗部干物质积累和分配及转运的影响
2018-05-04李媛媛李玉萍李红兵张岁岐
李媛媛,李玉萍,王 楠,李红兵,张岁岐,2
(1.黄土高原土壤侵蚀与旱地农业国家重点实验室/西北农林科技大学,陕西杨凌 712100;2.黄土高原土壤侵蚀与旱地农业国家重点实验室/中国科学院水利部水土保持研究所,陕西杨凌 712100)
小麦穗部在干物质积累、籽粒产量形成过程中发挥着重要的作用[1]。研究表明小麦穗部对产量的贡献率可能因环境和基因型的不同而存在差异[2]。Araus等[3]在研究遮穗对硬质小麦的影响时发现穗部对硬质小麦籽粒的贡献率为59%。在干旱等逆境胁迫下,小麦穗部表现出更高的贡献率,成为灌浆过程中的主要贡献器官[4]。Maydup等[5]研究发现,正常水分条件下,穗部对籽粒的贡献率为13%~30%,干旱条件下,贡献率上升至22%~45%。这些研究表明,小麦穗部可能在逆境环境下产量维持中起到缓冲作用,对籽粒充实有着重要贡献,尤其是在水分亏缺下具有明显的生理优势,因此,充分挖掘干旱条件下穗同化物生产潜力,对旱作小麦稳产、高产具有重要的理论及实际意义。
小麦籽粒产量一部分来自营养器官花前贮存物质的转运,另一部分来自花后光合产物的积累、转运及分配。姜 东等[6]认为小麦花前的物质积累主要向非籽粒营养器官(根、茎、叶)分配,干物质运转率低于20%,对籽粒的贡献率低于30%。郭天财等[7]认为花后干物质积累对籽粒贡献率达60%以上。张 磊等[8]认为水分亏缺下,水分敏感品种郑引1号颖壳、内外稃及籽粒的干物质分配率变幅显著,而旱地品种西农928分配率没有显著变化,即小麦不同器官干物质的转运和分配特征与品种关系密切。除了氮肥、栽培模式和品种基因型[7,9]外,水分也是影响小麦干物质积累和分配的重要因素[10-11]。一般认为干旱比正常水分条件下花前贮藏物质运转量大,但也有研究结果与此相反[12]。如干旱降低了小麦营养器官花前贮藏物质的转运量和转运率,增加了花前干物质转运量对籽粒产量的贡献率[13]。而过多的灌水会增加干物质向营养器官的积累分配,不利于籽粒产量的积累[14]。因此,深入研究水分胁迫对不同倍性小麦穗部干物质积累、转运特征及对产量形成的影响,挖掘小麦穗部物质生产潜力,对指导缺水地区小麦高产栽培具有重要应用价值。
黄明丽和赵紫平等[15-16]研究表明,随二倍体到四倍体再到六倍体的演化,小麦产量和水分利用效率显著增加,这是因为六倍体小麦花后较强的光合能力和较高的水分利用效率是产量提高的重要生理基础。前人研究大多集中于干旱对不同倍性小麦光合、产量和水分利用效率的影响[15-18],而有关干旱条件下不同倍性小麦穗部干物质分配规律及对籽粒产量的贡献率之间的差异、穗部干物质积累与转运特性及对水分胁迫响应的差异的研究少有报道。为此,本研究选取3个不同染色体倍性的小麦进化材料,通过对比干旱和灌水两种处理下不同倍性小麦品种穗部干物质分配规律及对籽粒产量的贡献率之间的差异,分析品种进化过程中水分状况对小麦穗部干物质积累、分配、转运及籽粒产量的影响,以期为进一步挖掘小麦穗部物质生产潜力、提高单产打下理论基础,也为明确不同倍性小麦穗部对产量的贡献率提供理论依据。
1 材料与方法
1.1 试验地概况
试验于2014年10月至2016年6月连续两个生长季在陕西杨凌中国科学院水土保持研究所(34°22′N,108°00′E)活动防雨棚内进行。该地气候属于暖温带季风区半湿润气候,年均温 10.7~13.7℃。降水量552.6~663.9 mm,全年降雨集中在夏秋季节,春冬季节多干燥。试验地土壤为耕作性塿土,质地重壤,田间最大持水量 24%,土壤容重 1.2 g·cm-3左右。通过CNC100型中子仪分别于播种前、开花期、收获期对每个小区10~200 cm 土层土壤含水量变化进行测定。小麦整个生育期的降雨量利用置于田间的降雨量筒测得。
2015-2016年小麦生长时期水分状况见表1。

表1 3种小麦各时期0~2 m土壤含水量Table 1 Water content of the experiment field at 0-2 msoil during the growth periods of three wheat species %
BM:碧蚂1号;YE:野生二粒小麦;YY:野生一粒小麦;Ir:灌水;Dr:干旱。同一列数字后的不同小写字母表示不同处理之间差异显著(P<0.05)。下同。
BM:T.aestivumBima 1;YE:T.dicoccoides;YY:T.boeoticum;Ir:Irrigation;Dr:Drought.Different small letters indicate significant differences among different water treatments of the different wheat species(P<0.05). The same below.
1.2 试验设计
1.2.1 试验材料
试验选用3个不同染色体倍性的小麦进化材料,包括:染色体组型为AA的野生一粒小麦(Triticumboeoticum,YY),2n=14;染色体组型为AABB的野生二粒小麦(T.dicoccoides,YE),为具AA染色体组的野生一粒小麦与具BB染色体组的拟斯卑尔脱山羊草(Aegilopsspeltoides)天然杂交并加倍所产生,4n=28;染色体组型为AABBDD的普通小麦品种碧蚂1号(T.aestivumBima1,BM),6n=42。
1.2.2 田间试验
播种日期分别为2014年10月10日和2015年10月14日,人工点播播种,次年5月底到6月初收获。共设干旱(返青期前正常降雨,返青期后遮雨且全生育期无灌水)和灌水(全生育期正常降雨,冬前、拔节期分别灌水600和800 kg·hm-2)2个水分处理。试验采用随机区组设计,每个处理3个重复,共18个小区。每个小区面积为7.26 m2(3.3×2.2 m),小区间用3 m深的水泥墙隔开,以防止水分渗出和相互渗入。小麦株行距分别为2和20 cm(人工点播)。试验期内土壤每年播前肥力较为稳定,两年平均土壤养分含量为:有机质18.81 mg·kg-1,全 N 0.99 g·kg-1,碱解N 67.00 mg·kg-1,全P 0.87 g·kg-1,速效P 19.40 mg·kg-1,速效K 150.40 mg·kg-1。试验田播前一次性施足基肥,施尿素150 kg·ha-1(以N计),过磷酸钙 120 kg·ha-1(以P2O5计),生育期内不再追肥,其他管理措施同一般大田试验。
1.3 测定项目和方法
1.3.1 干物质及穗部分配率测定
在小麦开花当天,每小区选择花期一致,长势、株高、穗子大小基本相同植株进行标记,开花当天开始取样,以后每5 d取样1次,直至完熟,每次每个品种的3个小区各取10个主茎带回实验室,贴地面去掉根部,将小麦植株的旗叶、其他叶、叶鞘、茎秆及穗部的穗轴、颖壳、籽粒分离,70 ℃ 烘48 h至恒重。不同器官分别称取干重。计算穗部各器官干物质分配率及对产量的贡献率、各器官干物质积累量与转运量。计算公式[11,19-21]如下:
穗部各器官干物质分配率[1]=穗部器官干重/穗部总干重×100%
花前干物质转运量=开花期干重-成熟期干重(除去籽粒产量)
花前干物质转运率=花前干物质转运量/开花期干重×100%
花前干物质转运对籽粒的贡献率=花前干物质转运量/成熟期籽粒干重×100%
花后干物质积累量=籽粒产量-花前干物质转运量
花后干物质积累对籽粒的贡献率=花后干物质积累量/成熟期籽粒干重×100%
1.3.2 产量性状测定
各小区在成熟期收割1 m2用于测产,风干后记录地上生物量、产量、千粒重、总穗数、小穗数及穗粒数。计算收获指数:
收获指数=籽粒产量/地上部生物产量
1.4 数据处理和分析
通过Excel 2010软件计算数据和作图,用SPSS 17.0统计分析软件进行数据差异显著性检验。数据处理和分析后发现2014年和2015年的试验结果基本一致,因此,本研究主要依据2015年生长季的实验数据来分析和说明。
2 结果与分析
2.1 水分胁迫对不同倍性小麦品种穗部各器官干物质分配的影响
小麦干物质积累与转运不仅与基因型有密切关系,而且受环境条件的影响。水分作为主要的环境因子,显著影响小麦光合产物的积累和分配,在正常供水和干旱胁迫两种水分条件下,穗轴干物质分配率均为六倍体>二倍体>四倍体(图1A);颖壳干物质分配率均为二倍体>六倍体>四倍体(图1B);籽粒干物质分配率均为六倍体>四倍体>二倍体(图1C)。
花后15 d(图1A、1B),与正常供水相比,3种小麦穗轴、颖壳干物质分配率在水分胁迫下均提高,水分胁迫可能通过改变小麦穗轴、颖壳中的光合同化物滞留,影响了同化物向籽粒的转运分配;花后35 d,水分胁迫下3种小麦的籽粒干物质分配率与正常供水相比均降低(图1C),穗轴、颖壳干物质分配率均提高(图1A、1B)。
2.2 水分胁迫对不同倍性冬小麦品种干物质转运特性的影响
小麦籽粒产量的高低与营养器官暂贮物质的积累与再转运有密切关系,小麦营养器官在开花前和开花后可临时贮存光合产物,到了籽粒灌浆后期再转运到籽粒中,形成产量。如表2所示,叶+茎+叶鞘、旗叶、穗部各器官(除穗轴外)的花前干物质转运量在两种水分条件下均为四倍体最高。其中叶+茎+叶鞘花前干物质转运量高于其他器官。叶+茎+叶鞘、旗叶、颖壳的花前干物质转运效率的变化趋势与花前干物质转运量相似,也表现为四倍体显著高于六倍体和二倍体小麦。结果表明,四倍体的花前干物质转运能力及转运效率高于六倍体和二倍体,有利于产量的增加。
由表3可知,四倍体的叶+茎+叶鞘、旗叶、穗部各器官花前干物质转运量对籽粒的贡献率高于六倍体和二倍体,3种小麦的叶+茎+叶鞘花前干物质转运量对籽粒的贡献率均高于其他器官。3种小麦穗部各器官花前干物质转运量对籽粒的贡献率为颖壳>穗轴。由表4可知,从二倍体→四倍体→六倍体的进化过程中,两种水分条件下,营养器官的花后干物质积累量呈显著升高趋势,而花后干物质积累量对籽粒的贡献率呈先降低后升高的趋势;六倍体的花后干物质积累量及其对籽粒的贡献率均大于其他两种小麦。
与正常供水相比,水分胁迫下,3种小麦各器官花前干物质转运量及转运效率显著降低,花后干物质积累量降低,而花后干物质积累量对籽粒的贡献率升高。水分胁迫下,3种小麦穗部各器官的花前干物质转运量及转运效率、花前干物质转运量对籽粒的贡献率显著降低,下降幅度为二倍体 > 六倍体 >四倍体,即四倍体小麦穗部器官的花前干物质转运量降低幅度最小。

图1 水分胁迫对小麦穗轴(A)、颖壳(B)、籽粒(C)干物质分配率的影响
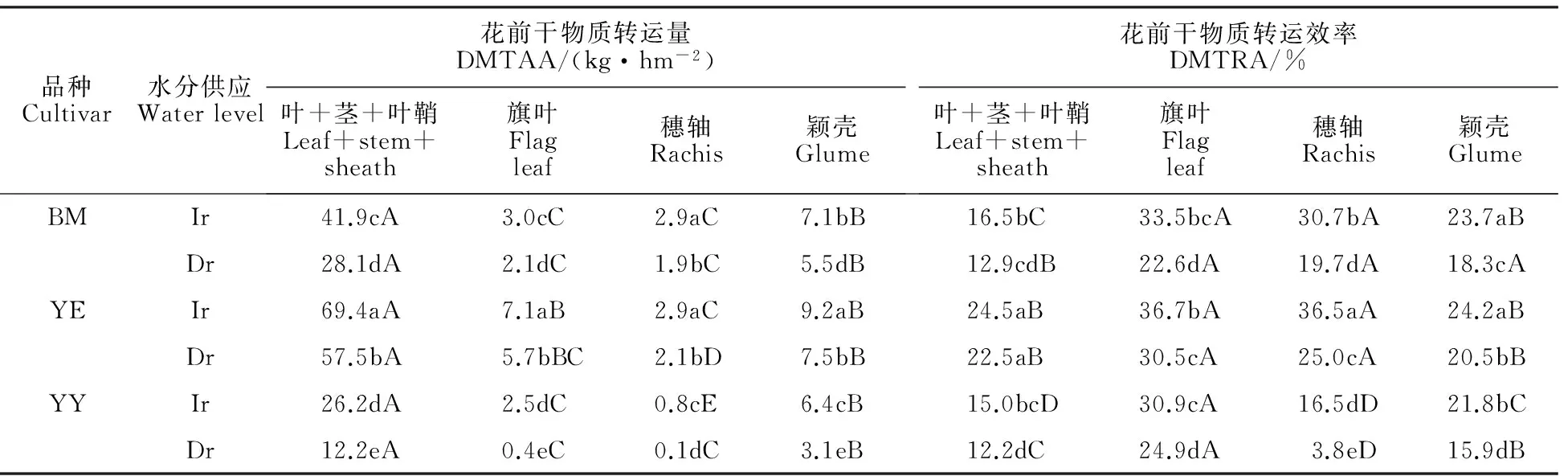
品种Cultivar水分供应Waterlevel花前干物质转运量DMTAA/(kg·hm-2)叶+茎+叶鞘Leaf+stem+sheath旗叶Flagleaf穗轴Rachis颖壳Glume花前干物质转运效率DMTRA/%叶+茎+叶鞘Leaf+stem+sheath旗叶Flagleaf穗轴Rachis颖壳GlumeBMIr41.9cA3.0cC2.9aC7.1bB16.5bC33.5bcA30.7bA23.7aBDr28.1dA2.1dC1.9bC5.5dB12.9cdB22.6dA19.7dA18.3cAYEIr69.4aA7.1aB2.9aC9.2aB24.5aB36.7bA36.5aA24.2aBDr57.5bA5.7bBC2.1bD7.5bB22.5aB30.5cA25.0cA20.5bBYYIr26.2dA2.5dC0.8cE6.4cB15.0bcD30.9cA16.5dD21.8bCDr12.2eA0.4eC0.1dC3.1eB12.2dC24.9dA3.8eD15.9dB
不同大写字母表示同一水分处理同一品种不同器官之间差异显著(P<0.05),下同。
Different capital letters indicate significant differences among different organs for the same wheat species under the same water treatment(P<0.05);DMTAA:Dry matter translocation amount before anthesis;DMTRA:Dry matter translocation ratio before anthesis. The same in other tables.

表3 两种水分处理下不同倍性小麦各营养器官对籽粒的贡献率Table 3 Contribution rate of different organs to grain in different ploidy wheat species under two water treatments
CDMTAATG:Contribution rate of dry matter translocation amount before anthesis to grains.

表4 水分胁迫对不同倍性小麦花后干物质积累量的影响Table 4 Effect of water stress on dry matter accumulation amount after anthesis in different ploidy wheat species
DMAAA:Dry matter accumulation amount after anthesis;CDMAAATG:Contribution rate of dry matter assimilation amount after anthesis to grains.
2.3 水分胁迫对不同倍性冬小麦品种农艺性状和产量构成因素及产量的影响
小麦籽粒产量形成由基因型和环境共同作用,水分是影响籽粒产量最重要的环境因素之一,显著影响小麦生育进程,从而影响籽粒产量。在拔节至灌浆期,干旱对小麦产量及产量构成因素有不同程度的影响,如表5所示,随染色体倍性由2n→4n→6n的进化过程,小麦的穗粒数、千粒重、籽粒产量和收获指数总体呈增加趋势,而生物量呈现先增加后降低的趋势,且不同倍性小麦品种间差异显著。
不同倍性小麦品种产量构成因素和产量均对不同水分处理敏感程度不一。与正常供水相比,水分胁迫下,3种小麦品种的产量构成因素均降低,产量亦显著降低;六倍体小麦籽粒产量、千粒重和每穗穗粒数分别降低了30.63%、14.43%、4.55%,四倍体小麦籽粒产量、每穗穗粒数分别降低了24.91%、12.18%,二倍体小麦籽粒产量、千粒重和每穗穗粒数分别降低了35.75%、4.96%、20.22%;即干旱对四倍体-野生二粒小麦的产量影响最小,产量最稳定,对二倍体野生一粒小麦的产量影响最大,产量下降幅度最高。

表5 水分胁迫对不同倍性小麦品种产量及其构成的影响Table 5 Effect of water stress on yield components in different ploidy wheat species
SNS:Spikelet number per spike;GNP:Grain number per spike;SN:Spike number;TGW:1 000-grain weight;GY:Grain yield;GM:Above-ground biomass,HI:harvest index.
3 讨 论
Maydup等[5]报道,穗部既是小麦光合器官,又具有临时库的作用,因此在小麦籽粒形成过程中具有不可忽视的作用,尤其是在水分亏缺的条件下。Gallagher等[22]研究表明,水分亏缺下,大麦籽粒中大多数同化物来自于灌浆前的累积,干旱加速了营养器官中贮藏物质的分解和输出,促进茎部等器官同化物向籽粒快速转运。而本研究中,水分胁迫下3种小麦的籽粒干物质分配率、各器官花前干物质转运量及转运效率与正常供水相比均降低,穗轴、颖壳干物质分配率均提高,表明干旱胁迫降低了营养器官开花前贮藏同化物向籽粒的再分配,从而降低了产量。
Fang等[23]认为小麦品种“长武134”的产量与抽穗后干物质积累量显著正相关,而与抽穗前干物质积累无显著差异。有研究表明,花后干物质积累量增加是品种产量提高的主要原因,是品种生产力提高的具体表现[24]。由表4可知,从二倍体→四倍体→六倍体的进化过程中,营养器官的花后干物质积累量呈显著升高趋势,而花后干物质积累量对籽粒的贡献率呈先降低后升高的趋势,表明六倍体的花后干物质积累能力高于四倍体和二倍体,这些参量与高产密切相关。
不同倍性小麦品种产量构成因素对不同水分处理敏感程度不一,水分胁迫下,四倍体产量下降幅度最小。一方面, 3种不同倍性小麦相比,四倍体小麦的穗轴、颖壳干物质分配率最低,而花前干物质转运量、转运效率及其对籽粒产量的贡献率显著高于其他两种小麦,水分胁迫下四倍体小麦的下降幅度最小,即干旱胁迫对四倍体小麦穗部干物质向籽粒的转运影响最小,说明四倍体小麦穗轴、颖壳中光合同化物滞留最少,干物质向籽粒转运分配较多,使得千粒重增加,为维持产量奠定了基础。另一方面,与六倍体和二倍体小麦相比,四倍体小麦从开花期到成熟期土壤水分的消耗最少,这可能与四倍体小麦的穗部在水分胁迫下能够维持较低的蒸腾速率和较高的水分利用效率有关[25],四倍体小麦具有更好的水分胁迫适应性,因而能够缓解水分胁迫导致的产量降低。因此水分胁迫下,四倍体小麦表现出更强的耐旱性。
细胞学实验已证实野生一粒小麦、野生二粒小麦和碧蚂1号这3种小麦材料存在一定的进化关系[26-27]。研究表明,随二倍体到四倍体再到六倍体的演化,小麦产量显著增加[17-18]。本研究结果与前人研究结果一致。在小麦进化过程中,产量的显著增加并不是完全依靠地上生物量的增加而获得的,而是与收获指数的显著增加相关[16]。同时,小麦生物量和产量的提高与千粒重的增加有密切关系。
4 结 论
本研究通过对比干旱和灌水两种处理下不同倍性小麦品种穗部干物质分配规律及对籽粒产量的贡献率之间的差异,分析品种进化过程中水分状况对小麦穗部干物质积累、分配、转运的变化特征及其产量的影响。发现与六倍体和二倍体小麦穗部干物质分配率相比,四倍体小麦的穗轴、颖壳干物质分配率最低,其花前干物质转运量、转运效率及对籽粒的贡献率最高。六倍体的花后干物质积累量及其对籽粒的贡献率均大于其他两种小麦。与旗叶相比,3种小麦颖壳花前干物质转运量对籽粒的贡献率较高,表明穗部对籽粒的贡献率高于旗叶。从二倍体→四倍体→六倍体的进化过程中,产量显著增加。四倍体小麦的转运量、转运效率和花前贡献率受水分胁迫影响最小,表现出更强的耐旱性。
参考文献:
[1] ABBAD H,EI JAAFARI S A,BORT J,etal.Comparative relationship of the flag leaf and the ear photosynthesis with the biomass and grain yield of durum wheat under a range of water conditions and different genotypes[J].Agronomie,2004,24:19.
[2] ARAUS J L,VILLEGAS D,APARICIO N,etal.Environmental factors determining carbon isotope discrimination and yield in durum wheat under Mediterranean conditions[J].CropScience,2003,43:170.
[3] ARAUS J L,BROWN H R,FEBRERO A,etal.Ear photosynthesis,carbon isotope discrimination and the contribution of respiratory CO2to differences in grain mass in durum wheat[J].Plant,Cell&Environment,1993,16:383.
[4] TAMBUSSI E A,NOGUÉS S,ARAUS J L.Ear of durum wheat under water stress:water relations and photosynthetic metabolism [J].Planta,2005,221:446.
[5] MAYDUP M L,ANTONIETTA M,GUIAMET J J,etal.The contribution of ear photosynthesis to grain filling in bread wheat(TriticumaestivumL.) [J].FieldCropsResearch,2010,119(1):48.
[6] 姜 东,于振文,李永庚,等.高产小麦营养器官临时贮存物质积运及其对粒重的贡献[J].作物学报,2003,29(1):31.
JIANG D,YU Z W,LI Y G,etal.Accumulation and redistribution of temporal reserves in vegetative organs and its contribution to grain weight in high yield winter wheat[J].ActaAgronomicaSinica,2003,29(1):31.
[7] 郭天财,宋 晓,马冬云,等.氮素营养水平对冬小麦碳氮运转的影响[J].西北植物学报,2007,27(8):1605.
GUO T C,SONG X,MA D Y,etal.Effects of nitrogen application rate on carbon and nitrogen transportation in winter wheat [J].ActaBotanicaBoreali-OccidentaliaSinica,2007,27(8):1605.
[8] 张 磊,吕金印,贾少磊.水分亏缺对小麦穗部光合特性及花前14C-同化物分配的影响[J].作物学报,2013,39(8):1514.
ZHANG L,LÜ J Y,JIA S L.Photosynthetic characteristics of spike and distribution of14C-assimilates accumulated before anthesis in wheat under water deficit condition [J].ActaAgronomicaSinica,2013,39(8):1514.
[9] 王桂良,叶优良,李欢欢,等.施氮量对不同基因型小麦产量和干物质累积的影响[J].麦类作物学报,2010,30(1):116.
WANG G L,YE Y L,LI H H,etal.Effect of nitrogen fertilizer application on grain yield and dry matter accumulation for different genotypes of winter wheat [J].JournalofTriticeaeCrops,2010,30(1):116.
[10] 闫永銮,郝卫平,梅旭荣,等.拔节期水分胁迫-复水对冬小麦干物质积累和水分利用效率的影响[J].中国农业气象,2011,32(2):190.
YAN Y L,HAO W P,MEI X R,etal.Effects of water stress-rewatering at jointing stage on dry matter accumulation and WUE of winter wheat[J].ChineseJournalofAgrometeorology,2011,32(2):190.
[11] 骆兰平,于振文,王 东,等.土壤水分和种植密度对小麦旗叶光合性能和干物质积累与分配的影响[J].作物学报,2011,37(6):1049.
LUO L P,YU Z W,WANG D,etal.Effects of planting density and soil moisture on flag leaf photosynthetic characteristics and dry matter accumulation and distribution in wheat[J].ActaAgronomicaSinica,2011,37(6):1049.
[12] 王志敏,王树安,苏宝林.小麦茎秆贮藏物质积累与运转[J].北京农业大学学报,1994,(4):369.
WANG Z M,WANG S A,SU B L.Accumulation and remobilization of stem reserves in wheat:Review [J].ActaAgriculturaeUniversitatisPekinensis,1994,(4):369.
[13] 姜 东,谢祝捷,曹卫星,等.花后干旱和渍水对冬小麦光合特性和物质运转的影响[J].作物学报,2004,30(2):175.
JIANG D,XIE Z J,CAO W X,etal.Effects of post-anthesis drought and water logging on photosynthetic characteristics and assimilates transportation in winter wheat[J].ActaAgronomicaSinica,2004,30(2):175.
[14] 董 剑,赵万春,高 翔,等.水氮调控对小麦植株干物质积累、分配与转运的影响[J].华北农学报,2012,27(3):196.
DONG J,ZHAO W C,GAO X,etal.Effects of irrigation and nitrogen control on accumulation,distribution and transport of dry matter of wheat plant [J].ActaAgriculturaeBoreali-Sinica,2012,27(3):196.
[15] 黄明丽,邓西平,周生路,等.二倍体、四倍体和六倍体小麦产量及水分利用效率[J].生态学报,2005,27(3):1113.
HUANG M L,DENG X P,ZHOU S L,etal.Grain yield and water use efficiency of diploid,tetraploid and hexaploid wheats [J].ActaEcologicaSinica,2005,27(3):1113.
[16] 赵紫平,邓西平,刘立生,等.灌浆期干旱对不同倍性小麦光合和产量的影响[J].麦类作物学报,2009,29(3):470.
ZHAO Z P,DENG X P,LIU L S,etal.Effect of water deficit on photosynthetic capacity and yield of different ploidy wheat during grain filling stage[J].JournalofTriticeaeCrops,2009,29(3):470.
[17] AUSTIN R B,MORGAN C L.Flag leaf photosynthesis ofTriticumaestivumand related diploid and tetraploid species [J].AnnalsofBotany,1982,49:177.
[18] BAMAKTRAMAH H S,HALLORAN G M,WILSON J H.Components of yield in diploid and hexaploid wheats(Triticumspp.) [J].AnnalsofBotany,1984,54:51.
[19] 黄 玲,高 阳,李新强,等.水分胁迫下不同年代冬小麦品种干物质积累与转运特性[J].中国生态农业学报,2013,21(8):943.
HUANG L,GAO Y,LI X Q,etal.Effects of water stress on dry matter accumulation and translocation in winter wheat cultivars planted at different ages [J].ChineseJournalofEco-Agriculture,2013,21(8):943.
[20] MA S Y,YU Z W,SHI Y.Effect of field border width for irrigation on dry matter accumulation and distribution,yield,and water use efficiency of wheat [J].ActaEcologicaSinica,2015,35:169.
[21] MA S C,DUAN A W,WANG R,etal.Root-sourced signal and photosynthetic traits,dry matter accumulation and remobilization,and yield stability in winter wheat as affected by regulated deficit irrigation [J].AgriculturalWaterManagement,2015,148:123.
[22] GALLAGHER J N,BISCOE P V,HUNTER B.Effects of drought on grain growth [J].Nature,1976,264:541.
[23] FANG Y,XU B C,TURNER N C,etal.Grain yield,dry matter accumulation and remobilization,and root respiration in winter wheat as affected by seeding rate and root pruning [J].EuropeanJournalofAgronomy,2010,33:257.
[24] 田中伟,王方瑞,戴廷波,等.小麦品种改良过程中物质积累转运特性与产量的关系[J].中国农业科学,2012,45(4):801.
TIAN Z W,WANG F R,DAI T B,etal.Characteristics of dry matter accumulation and translocation during the wheat genetic improvement and their relationship to grain yield [J].ScientiaAgriculturaSinica,2012,45(4):801.
[25] LI Y P,LI Y Y,LI D Y,ZHANG S Q.Photosynthetic response of tetraploid and hexaploid wheat to water stress [J].Photosynthetica,2017,55(3):454.
[26] 张正斌.小麦遗传学[M].北京:中国农业出版社,2001:1.
ZHANG Z B.Wheat Genetics[M].Beijing:China Agriculture Press,2001:1.
[27] ZHAO C X,DENG X P,SHAN L,etal.Changes in root hydraulic conductivity during wheat evolution [J].JournalofIntegrativePlantBiology,2005,47:302.
