植被活动对气候变化的响应过程研究进展
2018-04-19焦珂伟高江波吴绍洪侯文娟
焦珂伟,高江波,吴绍洪,侯文娟
1 中国科学院地理科学与资源研究所,中国科学院陆地表层格局与模拟重点实验室, 北京 100101 2 中国科学院大学,北京 100049
植被是全球气候变化的敏感指示器[1],气候环境中的水热条件是决定植被物候、生产力、分布格局等特征及其动态变化的主要非生物因素[2]。气候-植被关系及其对气候变化响应的研究,可为应对气候变化、提升和优化生态功能提供理论依据,因而成为全球变化研究的前沿和难点[3]。植被活动是表征植被生长、覆盖度、生产力等结构和功能属性的综合性概念,揭示植被活动对气候变化的响应过程,是气候-植被关系研究领域的重要内容[4]。
当前,植被活动响应气候变化过程的研究仍存在诸多难点,包括:由于植被对环境胁迫的敏感性和适应能力,气候变化对植被活动的影响呈现出显著的非线性作用规律[5- 6];另一方面,气候变化的影响涉及到植被光合、呼吸、蒸腾等多个紧密联系、相互作用的理化和生物过程,植被活动对气候因子的响应过程体现了多要素-多过程的综合效应特征[7- 8],形成具有复杂作用过程和空间分异规律的气候-植被关系[9-10]。鉴于此,本文针对植被活动响应气候变化的利弊特征、综合效应及其区域规律等方面的研究进展,多层次逐步推进以系统揭示植被活动响应气候变化复杂过程的研究态势与发展方向,以期推动该领域研究的持续深化。
1 水热要素对植被活动影响的阈值效应与利弊特征
植被总是在不断地去适应环境条件(包括气候特征)使其自身活动更为有利,是动态、非线性的过程,尤其在极端气候条件下,植被活动随气候因子的变化过程存在明显的阈值[11]。然而,当温度和降水等气候条件的改变超过植被的适应能力时,植被将会在结构功能等多方面受到抑制作用。气候变化对植被活动的影响机制与临界条件如表1所示。

表1 气候变化对植被活动利弊的影响机制与临界条件
1.1 温度升高对植被活动的影响
适度的增温能够对植被活动的增强产生积极作用。IPCC报告指出,20世纪全球温度升高约0.85℃(0.65—1.06℃)[12],而全球潜在植被NPP总量在这一增温环境下增长了13% (r2=0.40)[13]。Lucht等[14]基于构建的生物地球化学模型,发现20世纪后20年北半球高纬度升温0.8℃,植被呈现出变绿的趋势,说明了植被覆盖度对气温升高的正相关响应。植被活动对温度的响应机理主要体现在增暖对光合、呼吸等过程的影响程度,并且在时间尺度上有着不同的表现。徐振锋等[15]通过模拟增温实验表明,日平均气温增加2.9℃,土壤温度升高0.4℃会使得净光合速率、蒸腾速率和气孔导度分别增加17.4%,21.4%和33.9%,而对水分利用率没有显著的影响。Peuelas等[16]利用被动式夜间气温变暖处理,证明当温度大约增高1℃时,灌木林生态系统的地上生物量将会增加15%。春季增温对光合作用速率的增加要强于呼吸作用,而秋季则恰好相反,过去20年来北半球春季和秋季增温分别为1.1℃和0.8℃,因此目前北半球生态系统生产力仍处于增加的趋势[17]。此外,昼夜温度对植被的影响也不尽相同,植被的光合作用程度对日最高温的变化较为敏感,因此日间增温能够促进湿冷地区的植被活动,而呼吸作用则受到最高温和最低温的共同影响[18]。
然而,过高的升温会对植被活动过程产生不利影响。增温过高可能会加速土壤水分蒸发,形成干旱趋势,以南半球及北半球中低纬度的部分地区尤为明显,进而植被通过减小叶面积和降低光饱和点等方式防止自身失水,导致植被覆盖度相应减少,同时也会限制光合作用速率[19],影响有机物的合成;另一方面,增温使得植被的自养呼吸和蒸腾速率有所上升,加速了有机物的消耗,造成了植被净生产力的减少,因此对植被活动产生抑制作用。研究表明,未来增温对我国生态系统脆弱性存在着较为严重的影响,并处于加重的趋势,尤其是当增温超过3℃时,NPP较基准期(1961—1990年)减少13.43%[20]。王根绪等[21]建立了青藏高原高寒草地生物量对气候变化响应的多元回归模型,结果表明在降水量不变的条件下,若未来10年升温0.44℃,高寒草甸和高寒草原地上生物量分别减少2.7%和2.4%;若未来10年升温到达2.2℃时,高寒草甸和高寒草原地上生物量分别减少6.8%和4.6%。在2010—2050年这一时间段内,长江流域大部分地区在SRES B2气候情景下升温1.2—1.6℃,森林、郁闭灌丛和农作物的NPP有所减少[22]。此外,过去的气象观测数据表明,高纬度地区的增温幅度远高于低纬度地区,因此将会对中国东北地区的森林类型产生负面效应,其中当气温升高3℃时,针叶落叶林将被阔叶林取代[23]。
1.2 降水变动对植被活动的作用
气候变化将造成降水的分布、频率及强度发生显著变化,对陆地生态系统的水分平衡及植被生产力和覆盖度带来巨大影响。水分参与了植被的光合和蒸腾作用等生理生化过程,土壤中许多养分和矿物质要在溶于水的条件下才能被植物吸收,因此植被在水分减少的条件下会导致光合速率下降,有机物产量降低,生长、覆盖等植被活动受到抑制[24]。然而,当降水的增加超过植被所需时,还可能通过减少辐射量和增大相对湿度等形式间接对生长发育等植被活动产生不利影响[25]。随着温带干旱程度的加剧,过去30年植被活动与温度的相关性逐渐减弱,由0.72下降至0.32,与降水的相关性则有所增强[26],因而水分的保持成为植被进行生长发育和再生产的一种适应策略[27]。在年平均降水量大于600mm的地区,植被对热量因子的变化更敏感,而降水量略有减少可能会间接促进植被活动;而在水分不足的半干旱地区(年平均降水量小于600mm),降水则成为了植被活动的限制性因子[28]。
降水的改变能否引起植被活动的变化取决于是否对土壤水含量和植被可利用水分产生影响[29-30]。田间实验结果表明,集中的降水会使得半干旱区的草原生态系统地上NPP增加70%,而湿润区则下降18%[31]。春夏两季(暖干条件)降水变动对植被活动的影响程度要强于秋冬两季(湿冷条件),草地生态系统在降水同样减少80%的情况下,春季生物量的减少相比于冬季则更为明显[32];地中海森林在秋季降水减少97%的情况下仍未受到明显的影响,而在春季降水减少87%的情况下,叶片水势的负值显著增加,植被失水较为严重[33]。许多研究针对降水增加与减少对植被活动的不同影响进行比较,例如美国怀俄明州混合草草原的夏季降水增加50%时,生物量增加44%,而降水减少50%时生物量则下降18%[34];英格兰地区夏季草地覆被减少和增加的降水变化阈值分别为-90%和20%[35]。此外,热带雨林降水减少34%—40%时,地上NPP会下降22%,而当降水下降60%以上时,热带雨林的死亡率将上升34%[36]。
2 气候变化影响植被活动的多因素综合分析
植被活动对气候变化的响应过程在一定程度上依赖于特定区域的水热背景条件。在达到光合作用最适宜温度前,温度的升高会促进光合作用,土壤中的营养物质释放速度加快,养分的有效性提高[38];当超过这一温度时,营养物质消耗加速,干旱程度增强,负面效应显现[39]。此时降水的变化可以在一定程度上调节植被活动。吴杨周等[40]通过田间控制试验证明,增温2℃时植被的地上生物量增加46.0%,而再加上水分减少30%的条件后,增幅则变为19.8%。因此,水热因子的组合关系决定了植被结构、生长发育,以及植被地带性分布[41]。
植被活动过程随水热组合变化存在一个适宜的温度区间。在降水量较低时,适宜植被生长的温度区间相对较窄;随着降水量的增加,适宜植被生长的温度变幅有所拓宽。胡光成等通过对植被在水热组合共同驱动下的动态演化规律和趋势做出合理的预估,结果表明,在降水量大于250mm以上的低温区间内,植被分布及生产力主要由温度驱动,而且低温区的升温幅度明显高于最适宜植被活动的温度范围,因此高寒植被对气温响应更为敏感[42]。Wang等[43]的模拟结果显示,温度从2.7℃升高到3.9℃,降雨增加10%时,温带草原NPP将增加7%—21%;若温度升高7.5—7.8℃,降雨只增加10%,其NPP则降低24%。黄土高原水热组合直接影响植被物候的差异,气温高于9℃,降水在475—540mm之间时,植被活动对气候变化的响应过程最为明显[44]。
温度和降水的空间差异越大,表明植被的生境类型越复杂,而不同生境能够满足不同植被水热条件的需求[45]。对中国过去几十年的研究结果表明,在温度升高且降水增多的地区,植被覆盖度均表现一定程度的增加趋势,并且在未来气候以“暖湿化”为主导的地区,自然植被生产力将有所提高[46-47]。在温度普遍升高、西南部降水量增加显著而东北部减少的气候变化背景下,内蒙古温带草原的覆盖度和生产力有所提高,总面积将在本世纪末增加12.4%—18.5%[48]。在降水量满足植被活动的区域,如果气温处于增长趋势,则可能导致生长季开始期的提前与结束期的推迟,从而导致植被生长季延长。在季节尺度上,植被活动在不同季节对水热条件变化的敏感性不同,春季对温度的敏感性较降水高,夏季和秋季则对降水的敏感性较高[17]。
此外,光照也是植被所必需的气候因子,与温度、降水等气候条件共同对植被活动产生影响。光照强度直接影响植被光合作用的强弱,当光照强度超过光合作用饱和点时会引起叶绿素的分解,或使得细胞失水导致气孔关闭,造成光合作用减弱甚至停止[49]。一般情况下,光照在春秋两季与温度的联系较为密切,光照强的地方温度会有短时升高;而夏季降水量相对较大,大气中水汽充分,云量多且厚,降水对光照的影响超过温度且呈现出负相关[25]。可见在水分充沛的地区,光照成为了植被活动的一个主要限制因子。不同植被类型中,热带雨林对光照的响应要强于温度和降水[50];在中国的华东、华中和华南等湿润半湿润地区,植被活动受到光照的影响最为明显,由于光照强度的增加,NPP在20世纪末以每年0.99%的速率增长[51]。
3 植被活动响应气候变化的区域分异特征
过去几十年来气候变化致使中国植被/陆地生态系统的群落结构和生长过程发生改变,未来气候变化将继续对中国自然植被活动过程产生影响。而气候变化存在空间异质性,如升温幅度的空间差异性及升温与降温的空间分布[52]。在此背景下,植被活动对气候变化的响应过程具有显著的区域分异特征[53- 56]。气候-植被关系的相互作用机理及其空间变异,是地理学多要素相互作用及其区域异质性在全球变化研究领域中的实践和拓展。
3.1 气候变化影响下植被活动过程的空间异质性
区域气候模式模拟结果表明在气候变化过程中,高纬度地区的增温幅度远比低纬度地区的增温幅度大,因此目前普遍认为气候变化对高纬度地区植被活动过程的影响要比对低纬度地区的影响大得多。在我国的热带和亚热带地区,20世纪80年代至90年代的NPP以每年0.22%的速度持续增长,而在温带却以每年0.05%的速度不断下降[57]。针对我国东部植被活动的研究表明,1982—2006年华北平原、呼伦贝尔草原和洞庭湖平原的春季植被活动呈显著增强趋势,而长三角和珠三角地区则为显著下降趋势[58];21世纪前10年,东北平原、华北平原、黄土高原和新疆农业区的植被覆被显著增加,而内蒙古东北部和部分西部地区却有所退化[59],青藏高原的植被活动在气候变化的影响下,从东南到西北表现出“退化-增强-变化不大”的规律[60]。限制植被活动过程的气候要素同样体现出明显的区域差异,西北地区的植被活动主要受到水分限制,而东北、华北和青藏高原的植被活动则对温度的响应最为敏感[51]。Duo等[61]通过相关分析指出,从年际植被覆盖率来看,草地对气候要素的响应以降水为主导,而林地和农业用地则以温度为主导。
无论是过去还是未来,气候要素对植被活动过程的影响均体现出明显的空间变异。在我国干旱和半干旱地区,植被活动受到水分的限制,降水的增加能够促进植被生长,而温度升高则可能引起水分蒸散加强,加剧干旱趋势[62],因此与降水的关系较为密切;而在湿润半湿润地区,土壤水分含量高,植被活动不受水分限制,温度的升高有利于植被生长季的延长和干物质量的积累,因此与温度呈正相关[63]。然而,同样是在植被活动对温度响应正相关的区域,青藏高原未来30年的降水趋势有所增加,而高海拔的背景温度相对较低,温度的升高更有利于植被的光合作用,因此植被活动对温度的响应要强于降水[64];而华北地区的背景温度较高,未来30年降水为减少趋势,气候逐渐趋于暖干,降水对植被活动的影响高于温度[65]。可见,区域气候、水分和热量的空间异质性是形成植被活动空间分异规律的重要原因。
3.2 气候变化影响下植被带分布变化
植被的地理分布及其对气候变化的响应过程是生态系统与环境胁迫作用关系的具体表现,气候与植被的这种对应关系包含了气候系统与植被之间的相互作用和相互适应[66]。一旦气候变化导致水热组合格局的改变,势必导致植被带的范围、面积和界线发生变化。在以增暖为主要特征气候变化影响下,我国的植被带的分布存在着向高纬度和高海拔地区推移的趋势[67]。气候变化对中国主要森林类型的影响研究结果表明增暖可能导致北方森林带将有继续北移的趋势[68],对于亚热带常绿阔叶林,气温升高2℃,纬向上则向北扩大3个纬度,气温升高4℃,纬向上扩大6个纬度[69];垂直方向上,2℃增温可使得东北森林的垂直带谱普遍上移300m左右,而在CO2浓度倍增的条件下,温带落叶阔叶林的林线将升高100—160m,而亚热带山地针叶林和热带阔叶林分别升高150—350m和280—560m[70]。
不同区域植被带对未来气候变化响应预估方面开展了诸多研究[71- 75](图1)。在SRES排放情景下,热带森林、暖温带森林、热带草原和灌丛面积有所增加,而北方针叶林、温带森林、冻土苔原的面积减少;我国东部地区多数植被带发生北移,尤其是热带森林到本世纪中期将会向东北方向移动30—174km[72]。东北地区的中温带和暖温带的面积有所增加,植被带分布的界线发生北移,一些针叶林将逐渐被落叶阔叶林所取代[73];华北地区在未来气温降水单一变化的暖干气候作用下草原面积也将迅速扩大[26],但内蒙古草原植被在RCP排放情景下,面积却有所减少,南部界限大幅度北移[74];青藏高原的高寒植被在气候暖干化过程的控制下,高寒草原群落南向扩展,大部分转为温带草原,高寒草甸植被退化速率随着升温和降水减少的趋势有所加快,大部分转为山地寒温带针叶林[75]。
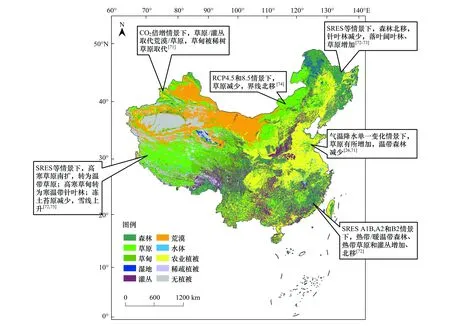
图1 不同气候变化情景下中国植被类型变化示意图Fig.1 Variations of Chinese vegetation types under different climate change scenarios
4 植被活动响应气候变化过程研究展望
气候变化对植被活动过程的影响有着明显的空间分异规律,这与气候变化趋势、植被类型、地形等特征有关[76],体现了植被活动对气候变化响应过程的复杂性。单一气候要素变化的影响,存在着由正面作用转为负面效应所发生的阈值[77],例如,当增温超过光合作用最适宜温度时,植被活动开始下降,进而在进一步增温的情况下,植被可能从形态和功能等方面形成适应[78-79];气候、土壤、地形等多要素交互作用与协同变化,共同对植被活动产生影响[80],例如升温加速了土壤水分的蒸散,并逐渐形成干旱环境,抑制植被生长,此时降水能够通过改变水热条件来调节温度对植被活动的作用[81]。针对植被活动对气候变化的响应过程,今后应加强如下两方面研究:
4.1 多方法集成,增强植被活动响应气候变化机理研究
当前,自然生态系统响应指标与评估方法缺乏统一性,不同研究成果之间难以比较。实验观测一直是研究气候变化对植被活动影响的重要手段。但如何将样点过程、典型物种的研究成果应用到区域尺度的生态系统,需要进行深入研究[82]。生态模拟和空间统计方法是实现观测实验到区域格局规律转换的必要手段,近年来,已从单一气候指标影响的分析发展到综合反映植被结构和功能的动态植被模型研发。然而,当前植被动态模型的发展还存在许多问题[83],如何考虑人类干扰、实现多尺度转换等,需要进一步解决(表2)。针对生态系统的复杂性、尺度性和对气候变化响应的时滞性[84],应综合集成不同方法,深入研究自然生态系统响应气候变化的过程,包括辨识波动、阈值、幅度、速率等非线性特征,揭示气候变化影响的适应性和脆弱性机理及其多尺度特征,为适应技术研发提供科学依据。
4.2 多要素融合,推进植被活动响应气候变化过程的耦合研究
国内外重大研究计划和组织都非常重视生态系统对气候变化的响应以及在应对气候变化中的作用,但基于生态系统响应过程的综合分析和区域性研究仍很缺乏[85]。首先,针对单一气候要素的变化趋势和影响过程研究较为成熟,然而将多要素进行综合的研究尚显不足[86],从系统视角对植被形成过程及其空间异质性的研究整体思路有待理清,特别是在不同气候要素的相互耦合作用下植被活动响应的内在机理[87]。其次,全球变化驱动下的植被格局呈现出动态变化特征,而传统的研究多关注静态关系以及局部的区域规律,反映出长时间的平均状态,缺乏对以要素变率为主的动态过程进行深入刻划[88]。因此在今后的研究中,需从动态的视角出发,系统研究气候变化-植被活动关系的变化规律及其在多要素、多过程交互作用下的机理,揭示植被格局变化的动态规律及区域差异,对格局变动的研究从长时段的平均态发展为阶段性的格局动态,揭示其变化趋势、幅度以及对气候变化的动态响应机制。
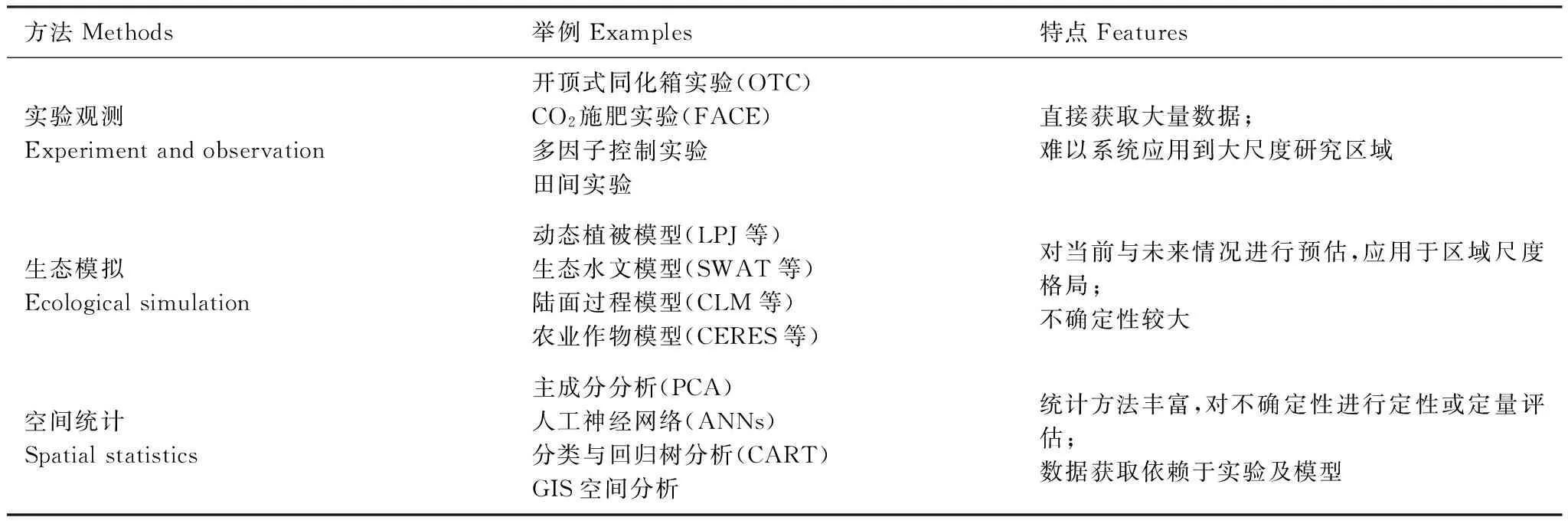
表2 植被活动响应气候变化的部分研究方法
OTC, Open-top Chamber; FACE, Free-air CO2Enrichment; LPJ, the Lund-Potsdam-Jena Dynamic Global Vegetation Model; SWAT, Soil and Water Assessment Tool; CLM, Community Land Model; CERES, Crop Environment Resource Synthesis; PCA, Principal Component Analysis; ANNs, Artificial Neural Networks; CART, Classification And Regression Tree
参考文献(References):
[1]Wang Q, Zhang Q P, Zhou W. Grassland coverage changes and analysis of the driving forces in Maqu County. Physics Procedia, 2012, 33: 1292- 1297.
[2]Zhu Z C, Piao S L, Myneni R B, Huang M T, Zeng Z Z, Canadell J G, Ciais P, Sitch S, Friedlingstein P, Arneth A, Cao C X, Cheng L, Kato E, Koven C, Li Y, Lian X, Liu Y W, Liu R G, Mao J F, Pan Y Z, Peng S S, Peuelas J, Poulter B, Pugh T A M, Stocker B D, Viovy N, Wang X H, Wang Y P, Xiao Z Q, Yang H, Zaehle S, Zeng N. Greening of the earth and its drivers. Nature Climate Change, 2016, 6(8): 791- 796.
[3]Levine J M. Ecology: a trail map for trait-based studies. Nature, 2015, 529(7585): 163- 164.
[4]Fang J Y, Tang Y H, Son Y. Why are East Asian ecosystems important for carbon cycle research? Science China Life Sciences, 2010, 53(7): 753- 756.
[5]Franzke C L E. Warming trends: nonlinear climate change. Nature Climate Change, 2014, 4(6): 423- 424.
[6]Ji F, Wu Z H, Huang J P, Chassignet E P. Evolution of land surface air temperature trend. Nature Climate Change, 2014, 4(6): 462- 466.
[7]Mahlstein I, Daniel J S, Solomon S. Pace of shifts in climate regions increases with global temperature. Nature Climate Change, 2013, 3(8): 739- 743.
[8]Moles A T, Perkins S E, Laffan S W, Flores-Moreno H, Awasthy M, Tindall M L, Sack L, Pitman A, Kattge J, Aarssen L W, Anand M, Bahn M, Blonder B, Cavender-Bares J, Cornelissen J H C, Cornwell W K, Díaz S, Dickie J B, Freschet G T, Griffiths J G, Gutierrez A G, Hemmings F A, Hickler T, Hitchcock T D, Keighery M, Kleyer M, Kurokawa H, Leishman M R, Liu K, Niinemets Ü, Onipchenko V, Onoda Y, Penuelas J, Pillar V D, Reich P B, Shiodera S, Siefert A, Sosinski E E Jr, Soudzilovskaia N A, Swaine E K, Swenson N G, Van Bodegom P M, Warman L, Weiher E, Wright I J, Zhang H X, Zobel M, Bonser S P. Which is a better predictor of plant traits: temperature or precipitation? Journal of Vegetation Science, 2014, 25(5): 1167- 1180.
[9]Chang N B, Vasquez M V, Chen C F, Imen S, Mullon L. Global nonlinear and nonstationary climate change effects on regional precipitation and forest phenology in Panama, Central America. Hydrological Processes, 2015, 29(3): 339- 355.
[10]Dong B W, Sutton R. Dominant role of greenhouse-gas forcing in the recovery of Sahel rainfall. Nature Climate Change, 2015, 5(8): 757- 760.
[11]Krishnaswamy J, John R, Joseph S. Consistent response of vegetation dynamics to recent climate change in tropical mountain regions. Global Change Biology, 2014, 20(1): 203- 215.
[12]IPCC. Climate Change 2013: the Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK: Cambridge University Press, 2013.
[13]Del Grosso S, Parton W, Stohlgren T, Zheng D L, Bachelet D, Prince S, Hibbard K, Olson R. Global potential net primary production predicted from vegetation class, precipitation, and temperature. Ecology, 2008, 89(8): 2117- 2126.
[14]Lucht W, Prentice I C, Myneni R B, Sitch S, Friedlingstein P, Cramer W, Bousquet P, Buermann W, Smith B. Climatic control of the high-latitude vegetation greening trend and Pinatubo effect. Science, 2002, 296(5573): 1687- 1689.
[15]徐振锋, 胡庭兴, 张力, 张远彬, 鲜骏仁, 王开运. 青藏高原东缘林线交错带糙皮桦幼苗光合特性对模拟增温的短期响应. 植物生态学报, 2010, 34(3): 263- 270.
[17]Piao S L, Ciais P, Friedlingstein P, Peylin P, Reichstein M, Luyssaert S, Margolis H, Fang J Y, Barr A, Chen A P, Grelle A, Hollinger D Y, Laurila T, Lindroth A, Richardson A D, Vesala T. Net carbon dioxide losses of northern ecosystems in response to autumn warming. Nature, 2008, 451(7174): 49- 52.
[18]Peng S S, Piao S L, Ciais P, Myneni R B, Chen A P, Chevallier F, Dolman A J, Janssens I A, Peuelas J, Zhang G X, Vicca S, Wan S Q, Wang S P, Zeng H. Asymmetric effects of daytime and night-time warming on northern Hemisphere vegetation. Nature, 2013, 501(7465): 88- 92.
[19]Zhang K, Kimball J S, Nemani R R, Running S W, Hong Y, Gourley J J, Yu Z B. Vegetation greening and climate change promote multidecadal rises of global land evapotranspiration. Scientific Reports, 2015, 5: 15956.
[20]Wu S H, Yin Y H, Zhao D S, Huang M, Shao X M, Dai E F. Impact of future climate change on terrestrial ecosystems in China. International Journal of Climatology, 2010, 30(6): 866- 873.
[21]王根绪, 胡宏昌, 王一博, 陈琳. 青藏高原多年冻土区典型高寒草地生物量对气候变化的响应. 冰川冻土, 2007, 29(5): 671- 679.
[22]苗茜, 黄玫, 李仁强. 长江流域植被净初级生产力对未来气候变化的响应. 自然资源学报, 2010, 25(8): 1296- 1305.
[23]程肖侠, 延晓冬. 气候变化对中国东北主要森林类型的影响. 生态学报, 2008, 28(2): 534- 543.
[24]Gourdji S M, Sibley A M, Lobell D B. Global crop exposure to critical high temperatures in the reproductive period: historical trends and future projections. Environmental Research Letters, 2013, 8(2): 024041.
[25]Ukkola A M, Prentice I C, Keenan T F, Van Dijk A I J M, Viney N R, Myneni R B, Bi J. Reduced streamflow in water-stressed climates consistent with CO2effects on vegetation. Nature Climate Change, 2015, 6(1): 75- 78.
[26]Piao S L, Nan H J, Huntingford C, Ciais P, Friedlingstein P, Sitch S, Peng S S, Ahlström A, Canadell J G, Cong N, Levis S, Levy P E, Liu L L, Lomas M R, Mao J F, Myneni R B, Peylin P, Poulter B, Shi X Y, Yin G D, Viovy N, Wang T, Wang X H, Zaehle S, Zeng N, Zeng Z Z, Chen A P. Evidence for a weakening relationship between interannual temperature variability and northern vegetation activity. Nature Communications, 2014, 5: 5018.
[27]Williams C A, Reichstein M, Buchmann N, Baldocchi D, Beer C, Schwalm C, Wohlfahrt G, Hasler N, Bernhofer C, Foken T, Papale D, Schymanski S, Schaefer K. Climate and vegetation controls on the surface water balance: synthesis of evapotranspiration measured across a global network of flux towers. Water Resources Research, 2012, 48(6): W06523.
[28]Clifford M J, Royer P D, Cobb N S, Breshears D D, Ford P L. Precipitation thresholds and drought-induced tree die-off: insights from patterns ofPinusedulismortality along an environmental stress gradient. New Phytologist, 2013, 200(2): 413- 421.
[29]Zeppel M J B, Wilks J V, Lewis J D. Impacts of extreme precipitation and seasonal changes in precipitation on plants. Biogeosciences, 2014, 11(11): 3083- 3093.
[30]Hoover D L, Knapp A K, Smith M D. Resistance and resilience of a grassland ecosystem to climate extremes. Ecology, 2014, 95(9): 2646- 2656.
[31]Heisler-White J L, Blair J M, Kelly E F, Harmoney K, Knapp A K. Contingent productivity responses to more extreme rainfall regimes across a grassland biome. Global Change Biology, 2009, 15(12): 2894- 2904.
[32]Bates J D, Svejcar T, Miller R F, Angell R A. The effects of precipitation timing on sagebrush steppe vegetation. Journal of Arid Environments, 2006, 64(4): 670- 697.
[33]Misson L, Degueldre D, Collin C, Rodriguez R, Rocheteau A, Ourcival J M, Rambal S. Phenological responses to extreme droughts in a Mediterranean forest. Global Change Biology, 2011, 17(2): 1036- 1048.
[34]Chimner R A, Welker J M, Morgan J, LeCain D, Reeder J. Experimental manipulations of winter snow and summer rain influence ecosystem carbon cycling in a mixed-grass prairie, Wyoming, USA. Ecohydrology, 2010, 3(3): 284- 293.
[35]Morecroft M D, Masters G J, Brown V K, Clarke I P, Taylor M E, Whitehouse A T. Changing precipitation patterns alter plant community dynamics and succession in an ex-arable grassland. Functional Ecology, 2004, 18(5): 648- 655.
[36]Brando P M, Balch J K, Nepstad D C, Morton D C, Putz F E, Coe M T, Silvério D, Macedo M N, Davidson E A, Nóbrega C C, Alencar A, Soares-Filho B S. Abrupt increases in Amazonian tree mortality due to drought-fire interactions. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(17): 6347- 6352.
[37]Fay P A, Blair J M, Smith M D, Nippert J B, Carlisle J D, Knapp A K. Relative effects of precipitation variability and warming on tallgrass prairie ecosystem function. Biogeosciences, 2011, 8(10): 3053- 3068.
[38]Michaletz S T, Cheng D L, Kerkhoff A J, Enquist B J. Convergence of terrestrial plant production across global climate gradients. Nature, 2014, 512(7512): 39- 43.
[39]Brohan P, Kennedy J J, Harris I, Tett S F B, Jones P D. Uncertainty estimates in regional and global observed temperature changes: a new data set from 1850. Journal of Geophysical Research-Atmospheres, 2006, 111(D12): D12106.
[40]吴杨周, 陈健, 胡正华, 谢燕, 陈书涛, 张雪松, 申双和, 陈曦. 水分减少与增温处理对冬小麦生物量和土壤呼吸的影响. 环境科学, 2016, 37(1): 280- 287.
[41]Fridley J D, Lynn J S, Grime J P, Askew A P. Longer growing seasons shift grassland vegetation towards more-productive species. Nature Climate Change, 2016, 6(9): 865- 868.
[42]胡光成, 金晓媚, 万力, 蔡晓雨. 祁连山区植被生长与水热组合关系研究. 干旱区资源与环境, 2009, 23(2): 17- 20.
[43]Wang Y H, Zhou G S, Wang Y H. Modeling responses of the meadow steppe dominated byLeymuschinensisto climate change. Climatic Change, 2007, 82(3/4): 437- 452.
[44]李强, 张翀, 任志远. 近15年黄土高原植被物候时空变化特征分析. 中国农业科学, 2016, 49(22): 4352- 4365.
[45]陈圣宾, 欧阳志云, 方瑜, 李振基. 中国种子植物特有属的地理分布格局. 生物多样性, 2011, 19(4): 414- 423.
[46]Bai W M, Wan S Q, Niu S L, Liu W X, Chen Q S, Wang Q B, Zhang W H, Han X G, Li L H. Increased temperature and precipitation interact to affect root production, mortality, and turnover in a temperate steppe: implications for ecosystem C cycling. Global Change Biology, 2010, 16(4): 1306- 1316.
[47]陈卓奇, 邵全琴, 刘纪远, 王军邦. 基于MODIS的青藏高原植被净初级生产力研究. 中国科学: 地球科学, 2012, 42(3): 402- 410.
[48]盛文萍, 李玉娥, 高清竹, 万运帆, 于贵瑞. 内蒙古未来气候变化及其对温性草原分布的影响. 资源科学, 2010, 32(6): 1111- 1119.
[49]Eiserhardt W L, Svenning J C, Kissling W D, Balslev H. Geographical ecology of the palms (Arecaceae): determinants of diversity and distributions across spatial scales. Annals of Botany, 2011, 108(8): 1391- 1416.
[50]Seddon A W R, Macias-Fauria M, Long P R, Benz D, Willis K J. Sensitivity of global terrestrial ecosystems to climate variability. Nature, 2016, 531(7593): 229- 232.
[51]朱文泉, 潘耀忠, 阳小琼, 宋国宝. 气候变化对中国陆地植被净初级生产力的影响分析. 科学通报, 2007, 52(21): 2535- 2541.
[52]IPCC. Climate Change 2014: Impacts, Adaptation, and Vulnerability. Part A: Global and Sectoral Aspects. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK, New York, NY: Cambridge University Press, 2014.
[53]丁一汇, 任国玉. 中国气候变化科学概论. 北京: 气象出版社, 2008.
[54]Burrows M T, Schoeman D S, Richardson A J, Molinos J G, Hoffmann A, Buckley L B, Moore P J, Brown C J, Bruno J F, Duarte C M, Halpern B S, Hoegh-Guldberg O, Kappel C V, Kiessling W, O′Connor M I, Pandolfi J M, Parmesan C, Sydeman W J, Ferrier S, Williams K J, Poloczanska E S. Geographical limits to species-range shifts are suggested by climate velocity. Nature, 2014, 507(7493): 492- 495.
[55]Friend A D, Lucht W, Rademacher T T, Keribin R, Betts R, Cadule P, Ciais P, Clark D B, Dankers R, Falloon P D, Ito A, Kahana R, Kleidon A, Lomas M R, Nishina K, Ostberg S, Pavlick R, Peylin P, Schaphoff S, Vuichard N, Warszawski L, Wiltshire A, Woodwardj F I. Carbon residence time dominates uncertainty in terrestrial vegetation responses to future climate and atmospheric CO2. Proceedings of the National Academy of Sciences of the United States of America, 2014, 111(9): 3280- 3285.
[56]De Jong R, Schaepman M E, Furrer R, De Bruin S, Verburg P H. Spatial relationship between climatologies and changes in global vegetation activity. Global Change Biology, 2013, 19(6): 1953- 1964.
[57]Wang Q, Zhao P, Ren H, Kakubari Y. Spatiotemporal dynamics of forest net primary production in China over the past two decades. Global and Planetary Change, 2008, 61(3/4): 267- 274.
[58]张学珍, 戴君虎, 葛全胜. 1982- 2006年中国东部春季植被变化的区域差异. 地理学报, 2012, 67(1): 53- 61.
[59]王筝, 赵天保, 侯美亭, 延晓冬. 21世纪初中国北方地区植被变化特征分析. 气候与环境研究, 2013, 18(2): 156- 164.
[60]王敏, 周才平, 吴良, 徐兴良, 欧阳华. 近10a青藏高原干湿状况及其与植被变化的关系研究. 干旱区地理, 2013, 36(1): 49- 56.
[61]Duo A, Zhao W J, Qu X Y, Jing R, Xiong K. Spatio-temporal variation of vegetation coverage and its response to climate change in North China plain in the last 33 years. International Journal of Applied Earth Observation and Geoinformation, 2016, 53: 103- 117.
[62]Feng X M, Fu B J, Piao S L, Wang S, Ciais P, Zeng Z Z, Lü Y H, Zeng Y, Li Y, Jiang X H, Wu B F. Revegetation in China′s Loess Plateau is approaching sustainable water resource limits. Nature Climate Change, 2016, 6(11): 1019- 1022.
[63]方精云, 朴世龙, 贺金生, 马文红. 近20年来中国植被活动在增强. 中国科学(C辑), 2003, 33(6): 554- 565.
[64]Wang C Z, Guo H D, Zhang L, Liu S Y, Qiu Y B, Sun Z C. Assessing phenological change and climatic control of alpine grasslands in the Tibetan Plateau with MODIS time series. International Journal of Biometeorology, 2015, 59(1): 11- 23.
[65]Xu H J, Wang X P. Effects of altered precipitation regimes on plant productivity in the arid region of northern China. Ecological Informatics, 2016, 31: 137- 146.
[66]周广胜, 何奇瑾. 生态系统响应全球变化的陆地样带研究. 地球科学进展, 2012, 27(5): 563- 572.
[67]Wilson R J, Gutiérrez D, Gutiérrez J, Martínez D, Agudo R, Monserrat V J. Changes to the elevational limits and extent of species ranges associated with climate change. Ecology Letters, 2005, 8(11): 1138- 1146.
[68]Wang H, Ni J, Prentice I C. Sensitivity of potential natural vegetation in China to projected changes in temperature, precipitation and atmospheric CO2. Regional Environmental Change, 2011, 11(3): 715- 727.
[69]Ni J. Impacts of climate change on Chinese ecosystems: key vulnerable regions and potential thresholds. Regional Environmental Change, 2011, 11(S1): 49- 64.
[70]贾庆宇, 王笑影, 吕国红, 谢艳兵. 气候变化对植被带影响研究进展. 安徽农业科学, 2010, 38(21): 11305- 11307.
[71]吕佳佳, 吴建国. 气候变化对植物及植被分布的影响研究进展. 环境科学与技术, 2009, 32(6): 85- 95.
[72]Zhao D S, Wu S H. Responses of vegetation distribution to climate change in China. Theoretical and Applied Climatology, 2014, 117(1/2): 15- 28.
[73]郭笑怡, 张洪岩. 生态地理分区框架下的大兴安岭植被动态研究. 地理科学, 2013, 33(2): 181- 188.
[74]苏力德, 杨劼, 万志强, 谷蕊, 闫玉龙, 高青竹. 内蒙古地区草地类型分布格局变化及气候原因分析. 中国农业气象, 2015, 36(2): 139- 148.
[75]Ni J, Herzschuh U. Simulating biome distribution on the Tibetan Plateau using a modified global vegetation model. Arctic, Antarctic, and Alpine Research, 2011, 43(3): 429- 441.
[76]Zhu Q A, Jiang H, Peng C H, Liu J X, Fang X Q, Wei X H, Liu S R, Zhou G M. Effects of future climate change, CO2enrichment, and vegetation structure variation on hydrological processes in China. Global and Planetary Change, 2012, 80- 81: 123- 135.
[77]Mackey K R M, Paytan A, Caldeira K, Grossman A R, Moran D, Mcilvin M, Saito M A. Effect of temperature on photosynthesis and growth in marineSynechococcusspp. Plant Physiology, 2013, 163(2): 815- 829.
[78]Pierangelini M, Stojkovic S, Orr P T, Beardall J. Photosynthetic characteristics of twoCylindrospermopsisraciborskiistrains differing in their toxicity. Journal of Phycology, 2014, 50(2): 292- 302.
[79]赵娜, 李富荣. 温度升高对不同生活型植物光合生理特性的影响. 生态环境学报, 2016, 25(1): 60- 66.
[80]刘晓, 丛静, 卢慧, 蒋军, 李广良, 宿秀江, 王秀磊, 李迪强, 张于光. 典型阔叶林的物种多样性分布和环境解释. 生态科学, 2016, 35(4): 125- 133.
[82]高江波, 焦珂伟, 吴绍洪, 郭灵辉. 气候变化影响与风险研究的理论范式和方法体系. 生态学报, 2017, 37(7): 2169- 2178.
[83]Zhao D S, Wu S H, Yin Y H. Responses of terrestrial ecosystems′ net primary productivity to future regional climate change in China. PLoS One, 2013, 8(4): e60849.
[84]Keenan T F, Gray J, Friedl M A, Toomey M, Bohrer G, Hollinger D Y, Munger J W, O′Keefe J, Schmid H P, Wing I S, Yang B, Richardson A D. Net carbon uptake has increased through warming-induced changes in temperate forest phenology. Nature Climate Change, 2014, 4(7): 598- 604.
[85]周广胜, 何奇瑾, 殷晓洁. 中国植被/陆地生态系统对气候变化的适应性与脆弱性. 北京: 气象出版社, 2015.
[86]Wan S Q, Norby R J, Ledford J, Weltzin J F. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland. Global Change Biology, 2007, 13(11): 2411- 2424.
[87]国际地圈生物圈计划秘书处, 国际地圈生物圈计划——科学计划与实施战略. 曲建升, 林海, 译. 北京: 气象出版社, 2006.
[88]吴绍洪, 赵艳, 汤秋鸿, 郑景云, 高江波, 梁涛, 葛全胜. 面向“未来地球”计划的陆地表层格局研究. 地理科学进展, 2015, 34(1): 10- 17.
