大花紫薇大小孢子的发生及雌雄配子体的发育
2018-04-10朱栗琼韦立迪韦艳春招礼军
朱栗琼 袁 娟 韦立迪 韦艳春 招礼军
(广西大学林学院,南宁 530004)
大花紫薇(Lagerstroemiaspeciosa),又名大叶紫薇、大果紫薇,隶属千屈菜科(Lythraceae)紫薇属(Lagerstroemia),分布于斯里兰卡、印度、马来西亚、越南及菲律宾,我国常见广东、广西、福建、海南及云南有栽培[1]。大花紫薇树形整齐美观,枝繁叶茂,花冠硕大,颜色艳丽,花期长;又在冬春季落叶颜色变化明显,春季萌发的嫩叶为淡紫色[2]。大花紫薇不仅是一种有很高观赏价值的园林绿化树种,同时也是优良的建筑用材和医药用材树种[3]。关于大花紫薇的研究多集中在化学成分和药理作用两方面[4~8],同时在开花习性和花粉活力[9]、种子发芽[10~11]及植物抗逆性[12]方面也有报道。大花紫薇以硕大的花朵及较高的观赏价值成为本属杂交研究中的重要母本候选者,掌握大、小孢子的发生和雌、雄配子体的发育规律可以为其成功的生殖和杂交组合提供胚胎学证据。关于紫薇属植物此方面的研究只见到紫薇(包括结实和不结实)[13]及日本矮紫薇[14]的报道,其余的种类只见到部分关于花粉活力的零星报道外,在胚胎学的其它方面还都缺乏研究资料。为此,笔者采用石蜡切片法对大花紫薇大、小孢子的发生和雌、雄配子体的发育展开了胚胎学的研究,旨在为植物生殖生物学积累资料,并为大花紫薇的选育工作提供科学依据。
1 材料与方法
研究材料采于广西大学校园内的大花紫薇行道树,样地地处南宁市,位于北纬22°50′,东经108°17′,属于亚热带季风湿润气候类型,夏长冬短,阳光充裕,全年太阳总辐射量为468 413.4 J·cm-2,每年平均日照时数为1 827 h;年平均气温为21.6℃,冬季最冷的1月平均气温为12.8℃,夏季最热的7、8月平均气温达28.2℃;极端最高温度40.4℃,极端最低气温-2.1℃;年平均降雨量1 304.2 mm,主要集中在4~9月,占全年79%;全年相对空气湿度达79%。
在校园内环境相似的位置选取10株长势中等、大小基本一致的成年大花紫薇植株,于2016年4~5月开花期间采集不同发育阶段的花蕾各100个,用70%FAA固定液(70%酒精∶冰醋酸∶甲醛=18∶1∶1)将材料固定、保存。室内采用常规石蜡切片法制片,爱氏苏木精染色,中性树胶封片,使用Nikon E100显微镜对切片进行观察并拍照。
用游标卡尺测定花蕾直径,每一级别各测30个,取平均数为其观测值。
2 结果与分析
2.1 花药壁的发育
在四分体前大花紫薇花药具四个大小和形态基本一致的花粉囊,中间被药隔分离(图版Ⅰ:1)。大花紫薇花药壁的形成属于双子叶型,发育初期的花药横切面近梨形,由一层原表皮包裹着一群无分化的基本分成组织构成(图版Ⅰ:2)。之后,花药四个角落的分生组织细胞迅速分裂,在花药角隅处逐渐形成具有四个裂瓣的花药雏形,每个裂瓣原表皮的内侧分化出几个孢原细胞,它们与周围的细胞外形上存在显著的差异,表现在体积更大、细胞质更浓,且具明显细胞核,染色较深(图版Ⅰ:3)。分化形成的孢原细胞经过平周分裂,形成内外两层细胞,外侧的为初生壁细胞,内侧的为初生造孢细胞(图版Ⅰ:4)。初生壁细胞随之进行平周和垂周双向分裂,逐渐形成3层呈同心圆排列的异质细胞,与表皮一起共同组成花药壁,由外及里依次是:表皮、药室内壁、中层、绒毡层(图版Ⅰ:5)。
表皮由一层细胞构成。在花药发育的初期,表皮细胞体积较大,在横切面上呈矩形,常见到细胞核(图版Ⅰ:5);在小孢子母细胞发育初期开始出现液泡化,之后逐渐伸长呈扁长方形,相对体积变小(图版Ⅰ:6)。在花药即将成熟时,部分表皮细胞逐渐解体,表皮细胞外壁形成明显的角质层(图版Ⅰ:7)。
药室内壁也是一层细胞构成,细胞呈扁长方形,细胞质液泡化程度较高,但细胞核较明显(图版Ⅰ:5)。花药发育成熟时,药室内壁出现带状的纤维加厚,由点朝径向延长,自此药室内壁转化为纤维层,但是在同侧的两个花粉囊的连接处,药室内壁细胞未见纤维状加厚,且细胞间隙较明显,这种特性有利于花粉囊壁的开裂(图版Ⅰ:7)。
中层由一层细胞构成,细胞体积较小,后期扁平状,细胞质丰富,细胞核所占比例较大(图版Ⅰ:5)。随着花粉的不断发育,至小孢子减数分裂时期,中层细胞被逐渐分解吸收(图版Ⅰ:8),至花粉成熟时中层细胞完全消失,被吸收殆尽(图版Ⅰ:7)。
绒毡层是花药壁最靠内的一层细胞。在次生造孢细胞的时期,绒毡层已经开始发育,这时的绒毡层细胞较小,排列紧密,细胞质较浓,单核(图版Ⅰ:9),次生造孢细胞即将发育成小孢子母细胞时,绒毡层细胞开始变大,(图版Ⅰ:10),核分裂为双核(图版Ⅰ:11),小孢子减数分裂之前,绒毡层生长加快,细胞内出现许多液泡和内含物,至此,绒毡层发育达到最高峰(图版Ⅰ:12)。之后,绒毡层细胞开始原位解体(图版Ⅰ:7)。到花粉成熟的时候,绒毡层细胞被彻底吸收而消失,花药壁只剩表皮和纤维层(图版Ⅰ:13),因此大花紫薇的绒毡层发育类型属于腺质性绒毡层。
四分体时,花药同一边相隔较远的两个花粉囊的药隔解体消失,连成一体,形成具有两个花粉囊的花药。随后,在两个花粉囊的相连处的药室内壁渐渐变薄,纤维层细胞干燥后,由于带状纤维发生收缩作用,花药壁断裂,花粉得以散出(图版Ⅰ:14)。
2.2 小孢子的发生及雄配子体的发育
初生造孢细胞经过若干次有丝分裂,发育为次生造孢细胞,细胞呈多边形,体积较大,细胞核明显,细胞质浓,与花药壁细胞之间的界限清楚,之后由其发育成小孢子母细胞,并逐渐分离(图版Ⅰ:15)。随后进入减数分裂时期,先后经历了减数分裂Ⅰ期(图版Ⅰ:16~19)和减数分裂Ⅱ期(图版Ⅰ:20~21),减数分裂Ⅰ期后胞质不分离,经过短暂的分裂间期后进入减数分裂Ⅱ期,最终胞质同时分离形成四分体。大多数四分体为四面体型(图版Ⅰ:22),极少数为十字交叉型(图版Ⅰ:23)。通过小孢子母细胞发育同步性观察的结果表明:在同一朵花的不同花药之间、同一个花药的不同药室之间的小孢子母细胞减数分裂常常不同步,大约相差1~2个分裂时期;同一药室内,小孢子母细胞减数分裂基本属于同步。
最初的四分体小孢子包裹在胼胝质壁中,随着胼胝质壁的逐渐分解,小孢子从四分体中释放出来,此时的小孢子体积比较小,细胞质浓,细胞核处于细胞中央,花粉壁稍厚,被视为单核早期小孢子(图版Ⅰ:24)。随着小孢子的体积不断增大,细胞质液泡化,形成一个中央大液泡,细胞核被逐渐推向靠近花粉壁的一侧,即成为单核靠边期小孢子(图版Ⅰ:25)。液泡消失后,通过不均等的有丝分裂和胞质分裂,小孢子发育形成含有一个体积较大的营养细胞和一个较小的生殖细胞,其中营养细胞近圆形,保持了原来小孢子的大部分原生质体,而生殖细胞凸透镜状,只包含少部分原生质体,此时2-细胞型花粉粒发育完成(图版Ⅰ:26~27)。大花紫薇成熟的花粉粒,细胞质浓厚,花药外壁增厚明显(图版Ⅰ:28),此时的花粉粒又称为雄配子体。笔者同时观察到少部分小孢子出现空瘪、异形或者体积偏小等发育异常的现象(图版Ⅰ:29)。
2.3 大孢子的发生及雌配子体的发育
大花紫薇子房为多心皮构成的多室子房,心皮和子房室数通常为6~7,以6心皮6心室为主(图版Ⅱ:1~2),每室着生20~50枚胚珠,中轴胎座。子房发育初期,随着心皮的发育,子房内壁也开始进行分化,其表皮下的细胞局部平周分裂后形成突起的胚珠原基(图版Ⅱ:3)。在胚珠原基的顶端位置逐渐发育成为珠心,而基部位置则发育成为珠柄。接着紧靠珠心基部的位置先后发生两环状的突起,首先发育形成一层内珠被,随后形成的是一层外珠被,故大花紫薇具双层珠被(图版Ⅱ:4)。珠心原基由一层表皮细胞包裹着的基本分生组织构成,随着珠心原基的进一步生长,基本分生组织在珠心组织的中部分化出一个体积较大,细胞质浓,细胞核明显,与周围细胞差别显著的孢原细胞(图版Ⅱ:5)。孢原细胞经分裂后产生周缘细胞和一个造孢细胞,其中周缘细胞进行平周分裂,加入珠心组织的构成,逐渐形成厚珠心;而造孢细胞经过不断生长发育,体积逐渐增大,最终发育成大孢子母细胞(图版Ⅱ:6)。由于胚珠生长发育过程中珠柄一侧的细胞分裂速度明显快于另一侧,从而使得整个珠孔端逐渐朝珠柄方向弯曲,最后整个胚珠倒转而形成倒生胚珠(图版Ⅵ:4)。大孢子母细胞进入减数分裂期,首先形成二分体(图版Ⅱ:7),进而再分裂发育成四分体(图版Ⅱ:8),四分体呈直线型排列。之后,远离合点端的3个大孢子先后退化消失,只剩近合点端的大孢子发育为功能性大孢子(图版Ⅱ:9),此种大孢子发育方式属于蓼形胚囊发育。
功能性大孢子持续发育,体积不断增大,液泡化增强,形成单核胚囊(图版Ⅱ:10),之后经过连续的三次有丝分裂,依次从单核变成为二核胚囊(图版Ⅱ:11)、四核胚囊(图版Ⅱ:12)、八核胚囊,接着珠孔端和合点端各有一核向中央移去构成两个极核(图版Ⅱ:13),两个极核融合后形成中央细胞,胚囊最终发育成为8核7细胞的成熟胚囊,即雌配子体,包括近珠孔端的1个卵细胞(图版Ⅱ:14)和2个助细胞(图版Ⅱ:15)、含2个极核的中央细胞及近合点端的3个反足细胞组成(图版Ⅱ:16)。
2.4 雌、雄蕊发育的对应关系
大花紫薇为两性花,同一朵花中雄蕊的分化稍早于雌蕊,但后期雌蕊发育速度较快,到花朵盛开的时候,雌、雄蕊发育基本可以达到同步成熟,同时花蕾的大小与内部的发育进程有一定的相关性(表1)。
表1花蕾大小与雌、雄蕊发育的对应关系
Table1Relationshipbetweenflowerbudsizeanddevelopmentstageofstamenandpistil

花蕾直径Diameterofflowerbud(mm)雄蕊发育Stamendevelopment雌蕊发育Pistildevelopment3~4.5孢原细胞Archosporialcell未分化Undifferentiatedcell4.5~6.5初生造孢细胞Primarysporogenouscell心皮原基Carpelprimordium6.5~7次生造孢细胞Secondarysporogenouscell胚珠原基Ovuleprimordium7~8小孢子母细胞Microsporemothercell孢原细胞Archosporialcell8~8.5减数分裂时期Meiosisstage大孢子母细胞Megasporemothercell8.5~9.5小孢子四分体Microsporetetrad功能性大孢子Functionalmegaspore9.5~13单核花粉Mononuclearpollen二核胚囊、四核胚囊2-nucleated&4-nucleatedembryosac12~14.52-细胞成熟花粉Two-celledpollens八核,七细胞八核胚囊8-nucleatedembryosacwith7cells13~16成熟花粉粒Maturepollengrains成熟胚囊Matureembryosac
3 讨论
大花紫薇的雄蕊为4个花粉囊,花药壁从外到内依次包括表皮、药室内壁、中层和绒毡层,各由一层细胞组成,其发育类型属双子叶型;绒毡层属腺质绒毡层;小孢子母细胞减数分裂时为同时型,四分体排列方式多为四面型,偶见十字交叉型;成熟花粉粒为2-细胞型,具3孔沟。在已有对紫薇花粉的研究中,结实的“红叶”紫薇与不结实的“湘韵”紫薇[13],以及日本矮紫薇[14]的成熟花粉粒均属3-细胞型,而本次笔者的研究发现大花紫薇的成熟花粉粒属于2-细胞型,这可作为紫薇属分类的一个重要而可靠的依据;除此之外,“红叶”和“湘韵”紫薇的四分体均为正四面体,日本矮紫薇除花药偶见6室外,4种紫薇在其余的各方面特征上均相似。
一般植物的小孢子母细胞减数分裂是高度同步的[15],大花紫薇在其同一药室中基本保持一致,但在不同药室发生的小孢子母细胞减数分裂,有时会相差1~2时期,表现出不同步性。董立格对“晚银桂”(Osmanthusfragrans)[16]、许小连对羊角槭(Aceryanjuechi)[17]的小孢子母细胞减数分裂的研究也得到相似的结果。究其原因可能是这类植物长期适应其传粉机制及本地环境的结果,这有利于延长传粉时间,为繁殖的成功率提供了保障,在进化上有一定的积极意义[18]。
花粉发育异常现象通常是由外界环境或者遗传机制等因素引起,如极端气候、小孢子母细胞减数分裂异常、绒毡层的发育失常以及雄配子体发育不良等。本研究中这些因素均可排除,同时从丰硕的结实率来看,大花紫薇出现的极少数异常花粉的现象受遗传因素影响的可能性较小,可能主要是因量多与营养供应有关。
大花紫薇雌蕊为子房上位,心皮数6~7,每个心皮单独形成一个子房室,中轴胎座,倒生胚珠,双珠被,厚珠心;大孢子减数分裂过程所形成的4个大孢子呈直线排列,合点端大孢子发育为功能性大孢子,其余3个大孢子先后退化消失;胚囊为单胞蓼型的发育方式,成熟胚囊为7细胞8核。大花紫薇大孢子发生和雌配子体发育特征与紫薇和日本矮紫薇[14]基本一致,只是后者的心皮及心室数为4-6。这一研究结果为紫薇属植物胚囊发育的类型确定提供了新的佐证资料。
同一朵花中,大花紫薇的雄蕊、雌蕊发育的时间及发育的速度有一定差异,雄蕊分化启动稍早于雌蕊,但到花朵盛开的时候,雌、雄蕊发育基本可以达到同时成熟,与许欢[13]的研究结果一致。另外,大花紫薇的雌、雄蕊发育的进程与花朵的外形和大小呈一定相关性,可通过花部的外形特征初步判断内部雌、雄配子的发育程度,了解这一发育的相关规律,可以为进一步探讨大花紫薇的有性遗传过程、遗传育种、组织培养等的研究工作提供有益参考。
1.方文培,张泽荣.中国植物志:第52卷[M].北京:科学出版社,1983:97.
Fang W P,Zhang Z R.Flora of China:Volume 52[M].Beijing:Science Press,1983:97.
2.广西科学院广西植物研究所.广西植物志:第1卷[M].南宁:广西科学技术出版社,1991.
Guangxi Institute of Botany.Flora of Guangxi:Volume 1[M].Nanning:Guangxi Science and Technology Press,1991.
3.Murakami C,Myoga R,Kasai K,et al.Screening of plant constituents for effect on glucose transport activity inEhrlichascitestumour cells[J].Chemical & Pharmaceutical Bulletin,1993,41(12):2129-2131.
4.王燕,孙连娜,楼永明.大花紫薇化学成分与药理作用研究进展[J].福建分析测试,2014,23(5):20-24.
Wang Y,Sun L N,Lou Y M.Advances in research of the chemical constituents and pharmacological effects ofLagerstroemiaspeciosa(Linn.) Pers[J].Fujian Analysis & Testing,2014,23(5):20-24.
5.Hattori K,Sukenobu N,Sasaki T,et al.Activation of insulin receptors by Lagerstroemin[J].Journal of Pharmacological Sciences,2003,93(1):69-73.
6.Suzuki Y,Hayashi K,Sakane I,et al.Effect and mode of action of banaba(LagerstroemiaspeciosaL.) leaf extracts on postprandial blood glucose in Rats[J].Nippon Eiyo Shokuryo Gakkaishi,2001,54(3):131-137.
7.Liu F,Kim J K,Li Y S,et al.An extract ofLagerstroemiaspeciosaL. has insulin-like glucose uptake-stimulatory and adipocyte differentiation-inhibitory activities in 3T3-L1 cells[J].Journal of Nutrition,2001,131(9):2242-2247.
8.Sinhababu A,Das S,Laskar S,et al.Characterization and identification of antibacterial components in extracts of seeds fromLagerstroemiaspeciosa[J].Advances in Food Sciences A,1999,21(1-2):19-22.
9.任翔翔,张启翔,潘会堂,等.大花紫薇开花及花粉特性研究[J].安徽农业科学,2009,37(28):13507-13509.
Ren X X,Zhang Q X,Pan H T,et al.Study on the characteristics of florescence and pollen inLagerstroemiaspeciosa[J].Journal of Anhui Agricultural Sciences,2009,37(28):13507-13509.
10.钟文勇,陈建丽,许鸿源.大花紫薇种子发芽特性研究[J].广西农业生物科学,2005,24(2):164-166.
Zhong W Y,Chen J L,Xu H Y.Studies on the characteristics of seed germination ofLagerstroemiaspeciosa[J].Journal of Guangxi Agricultural and Biological Science,2005,24(2):164-166.
11.许鸿源,钟文勇,陈建丽,等.几种因素对大花紫薇种子萌发的影响[J].种子,2005,24(9):57-58,61.
Xu H Y,Zhong W Y,Chen J L,et al.Influence of several factors on germination ofLagerstroemiaspeciosaseeds[J].Seed,2005,24(9):57-58,61.
12.滕维超,郑绍鑫,覃梅,等.大花紫薇幼苗对盐胁迫的生理响应[J].东北林业大学学报,2015,43(7):31-34.
Teng W C,Zheng S X,Qin M,et al.Physiological mechanism ofLagerstroemiaspeciosaL.seedlings under salt stress[J].Journal of Northeast Forestry University,2015,43(7):31-34.
13.许欢.结实与不结实紫薇生物学特性比较研究[D].长沙:中南林业科技大学,2015.
Xu H.Biological characteristic comparison research between fructiferous and acarpousLagerstroemiaindica[D].Changsha:Central South University of Forestry and Technology,2015.
14.郭玉敏.日本矮紫薇花芽形态分化及在武汉的引种适应性研究[D].武汉:华中农业大学,2006.
Guo Y M.Studies on flower bud morphodifferentiation and adaptability of introducedLagerstroemiaindicacv.in Wuhan[D].Wuhan:Huazhong Agricultural University,2006.
15.胡适宜.被子植物胚胎学[M].北京:人民教育出版社,1982.
Hu S Y.Embryology of Angiospermis[M].Beijing:People’s Education Press,1982.
16.董立格,丁雨龙,王贤荣,等.‘晚银桂’的小孢子发生和雄配子体发育[J].南京林业大学学报:自然科学版,2015,39(4):44-50.
Dong L G,Ding Y L,Wang X R,et al.The microsporogenesis and the development of male gametophyte inOsmanthusfragrans‘Wanyingui’[J].Journal of Nanjing Forestry University:Natural Science Edition,2015,39(4):44-50.
17.许小连,金荷仙,陈香波,等.濒危植物羊角槭小孢子发生与雄配子体发育研究[J].植物分类与资源学报,2012,34(4):339-346.
Xu X L,Jin H X,Chen X B,et al.Studies on the Formation of Microspores and Development of Male Gametophyte inAceryanjuechi(Aceraceae)[J].Plant Diversity and Resources,2012,34(4):339-346.
18.赵桦,孟华菊,徐娟.七叶树小孢子发生及雄配子体发育研究[J].西北植物学报,2011,31(2):217-222.
Zhao H,Meng H J,XU J.Microsporogenesis and male gametogenesis inAesculuschinensisBunge[J].Acta Botanica Boreali-Occidentalia Sinica,2011,31(2):217-222.
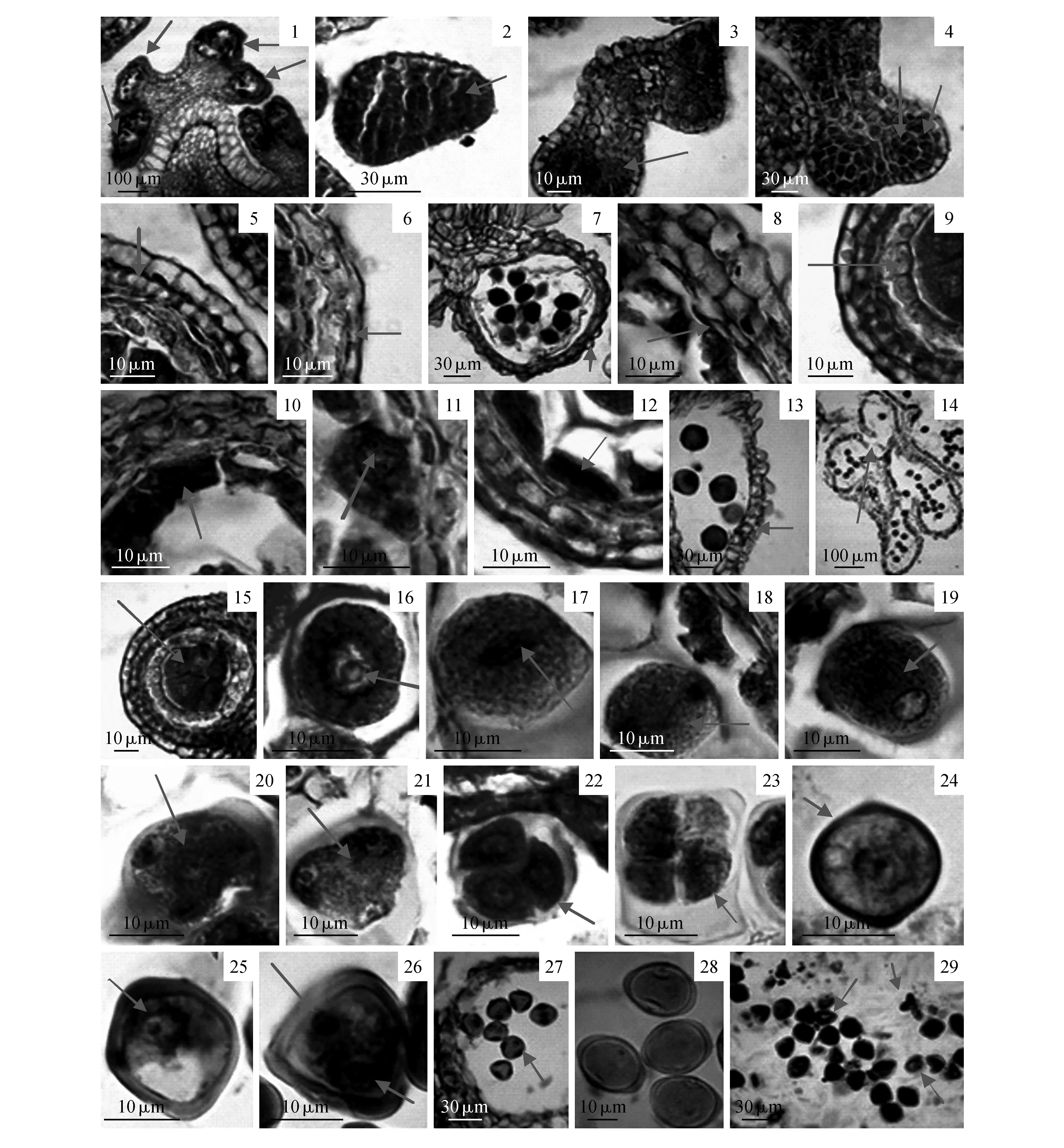
图版Ⅰ 大花紫薇小孢子发生和雄配子体发育 1.花药具4个花粉囊;2.未分化花药;3.孢原细胞;4.孢原细胞平周分裂产生周缘细胞和初生造孢细胞;5.花药壁;6.呈扁长方形的表皮细胞;7.表皮细胞角质化;8.中层细胞被挤压为扁平状;9.发育初期的绒毡层细胞;10.绒毡层细胞质浓厚;11.绒毡层细胞二核;12.绒毡层细胞径向生长;13.药室内壁纤维状加厚;14.花药开裂;15.小孢子母细胞;16.小孢子母细胞减数分裂前期Ⅰ;17.小孢子母细胞减数分裂中期Ⅰ;18.小孢子母细胞减数分裂后期Ⅰ;19.小孢子母细胞减数分裂末期Ⅰ;20.小孢子母细胞减数分裂后期Ⅱ;21.小孢子母细胞减数分裂末期Ⅱ;22.四面体型小孢子;23.十字交叉型四分体;24.单核早期小孢子;25.单核靠边期小孢子;26~27. 2-细胞花粉;28.成熟花粉粒;29.部分空瘪、变形、体积小等发育不正常的小孢子PlateⅠ Microsporogenesis and development of male Gametophyte of L.speciosa 1.The anther with four pollensacs; 2.Undifferentiated anther; 3.Archesporium; 4.The archesporium is divided into a parietal cell and a sporogenous cell,by the periclinal division; 5.Anther wall; 6.Flattened epidermal cell; 7.Keratinized epidermis cell; 8.Middle layer cell was flattened by extruding; 9.Tapetum cells in beginning stage; 10.Tapetum cells with dense cytoplasm; 11.Tapetum cell with two nucleus; 12.Tapetum cell radial growth; 13.Fibrous and thickened endothecium; 14.Anther dehiscence; 15.Microspore mother cell(MMC); 16.Meiosis prophase Ⅰ of MMC; 17.Meiosis metaphase Ⅰ of MMC; 18.Meiosis anaphase Ⅰ of MMC; 19.Meiosis telophase Ⅰ of MMC; 20.Meiosis anaphaseⅡof MMC; 21.Meiosis telophaseⅡof MMC; 22.Tetrahedral microspore; 23.Decussate tetrads; 24.Mononuclear microspore in early stage; 25.Mononuclear microspore in nucleus located aside stage; 26-27.Two-celled pollens; 28.Mature pollen grains; 29.Abnormal microspores
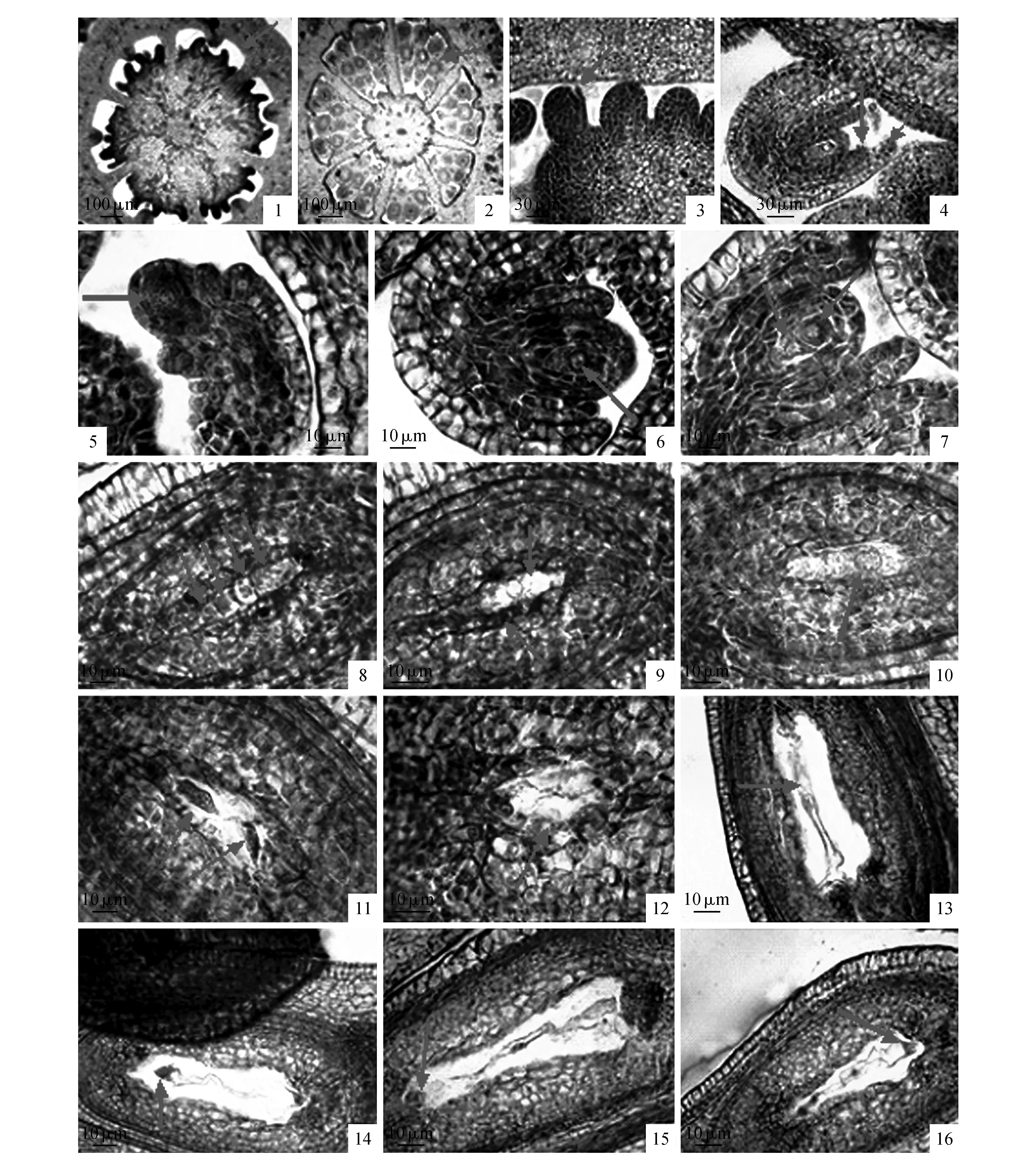
图版Ⅱ 大花紫薇大孢子发生及雌配子体发育 1.子房室数量为6;2.子房室数量为7;3.胚珠原基4.双层珠被;5.大孢子孢原细胞;6.大孢子母细胞;7.大孢子二分体;8.直线排列的四分体大孢子;9.靠珠孔端大孢子相继退化,合点端大孢子发育为功能大孢子;10.单核胚囊;11.二核胚囊;12.四核胚囊;13.成熟胚囊的两个极核;14.成熟胚囊的卵细胞;15.成熟胚囊的助细胞;16.成熟胚囊的反足细胞Plate Ⅱ Megasporogenesis and development of female gametophytes of L.speciosa 1.Each ovary has 6 locules; 2.Each ovary has 7 locules; 3.Anlage of ovule; 4.Double layers integument; 5.The megaspore archesporial cell; 6.The megaspore mother cell; 7.The megaspore dyad; 8.Four megaspores forming one liner tetrad; 9.The chalazal megaspore was functional, while the other three megaspores in micropyle degenerated; 10.Mononuclear embryo sac; 11. 2-nucleated embryo sac; 12. 4-nucleated embryo sac; 13.Two polar nuclei in the mature embryo sac; 14.The egg in mature embryo sac; 15.Two synergids in the mature embryo sac; 16.The antipodal cells in the mature embryo sac
