六道沟流域2种典型灌木不同季节水分来源及利用效率
2018-04-08杨国敏
杨国敏,王 爱,王 力,3*
(1 西北农林科技大学 资源环境学院, 陕西杨陵 712100;2 国家林业局调查规划设计院, 北京 100714;3 中国科学院 水利部 水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 陕西杨陵 712100)
风蚀水蚀交错区是黄土高原比较独特的自然地带,由于该区处于干旱半干旱的过渡地带,其自然条件恶劣,成为中国水土流失最为严重的地区,也是黄河粗泥沙的集中来源区。六道沟小流域位于该交错区的强烈侵蚀中心,由于水土流失导致的土地干旱、贫瘠,空气湿度和土壤含水量较低,植被类型多以耐旱的乔灌木和草本为主[1]。由于土地的不合理开发利用和天然植被的严重破坏,该流域水土流失剧烈,植被退化严重,生态环境脆弱。为了改善当地的生态环境,该区实施了大规模的植被建设,但由于地处干旱半干旱过渡地带,降水较少,加上土壤储水能力差,水资源亏缺一直是植被生长的主要限制因子。水分亏缺影响着植物的分布、光合特征和代谢水平,植物只有适应干旱环境才能维持正常的生长发育。柠条(Caraganakorshinskii)和沙柳(Salixpsammophila)作为该区分布范围较广的典型灌木树种,经过长期的自然选择和协同进化已表现出较强的生态适应性,然而其应对干旱胁迫所采取的水分利用策略和机制尚不清楚。因此,研究流域植物对有限水资源的利用情况,即植物的水分来源和水分利用效率对该区的植被恢复与重建具有重要的现实意义。
稳定氢氧同位素技术作为研究植物水分来源的一种新技术,近年来广泛应用于国内外的研究。研究发现,植物在不同季节遭受不同程度的水分胁迫,会采取不同的水分利用方式[2]。Dai等[3]比较了古尔班通古特沙漠中梭梭(Haloxylonammodendron)和白梭梭(Haloxylonpersicum)的根系吸水来源,发现在土壤表层水分充足的早春,梭梭主要利用浅层土壤水,白梭梭主要利用中层土壤水;在夏季白梭梭主要吸收深层土壤水,梭梭主要利用地下水。朱建佳等[4]通过研究沙拐枣(Calligonummongolicum)、合头草(Sympegmaregelii)、驼绒藜(Ceratoideslatens)和麻黄(Ephedraprzewalskii)的水分来源发现,生长季初期植物的主要水分来源是河水和地下水,生长季中后期,合头草主要利用浅层土壤水,其他三种植物较多的利用较深层土壤水和地下水。丁亚丽等[5]研究表明枫香(Liquidambarformosana)旱季和雨季始终以浅层土壤水为主要水分来源;尾巨桉(Eucalyptusgrandis)旱季上坡主要利用深层土壤水,下坡利用浅层土壤水,而雨季上坡依赖浅层土壤水,下坡主要利用深层土壤水。
本试验以中国科学院水利部水土保持研究所神木侵蚀与环境试验站所处的六道沟小流域为研究样地,选择该区典型灌木柠条和沙柳为研究对象,测定了旱季和雨季2种灌木木质部水及其潜在水源的稳定氢同位素值、叶片水势、气孔导度、光合速率和蒸腾速率等生理因子以及整个生长季植物叶片的δ13C值,对比不同季节柠条和沙柳的水分来源和水分利用效率的变化,以期阐明这2种灌木的水分利用策略及其对干旱环境的适应机制,为风蚀水蚀区植被恢复提供科学参考。
1 材料和方法
1.1 研究区概况
本试验设在陕西省榆林市神木县六道沟小流域(E110°21′~110°23′,N38°46′~38°51′),地处黄土高原与鄂尔多斯高原的结合部,是黄土高原向毛乌素沙地过渡以及森林草原向典型草原过渡的中间地带,又是水蚀严重的陕北黄土丘陵区向风蚀剧烈的毛乌素沙地的过渡地带,自然条件的过渡特征十分明显。流域面积约6.89 km2,海拔约1 094~1 274 m。属于中温带半干旱气候,冬天寒冷干燥,夏天温暖湿润,年平均气温8.4 ℃,月平均最低温-9.7 ℃(1月),最高温23.7 ℃(7月),极端最高温38.9 ℃,极端最低温-28.1 ℃,最高最低温相差67 ℃。年内和年际气候变化剧烈,暴雨、洪涝、干旱、冰雹、沙尘暴等自然灾害频繁发生[6]。全年日照时数2 836 h,年总太阳辐射量为5 922 MJ/m2,多年平均降水量437.4 mm,70%以上的降雨集中在7~9月份。主要土壤类型为黄绵土、沙黄土以及在沙地上发育起来的风沙土、坝地於土。黄绵土、风沙土和沙黄土的饱和含水量分别为0.44、0.32和0.40 cm3·cm-3;田间持水量分别为0.183、0.11和0.180 cm3·cm-3;饱和导水率分别为3.66、17.26和6.96 cm·h-1;萎蔫系数分别为0.064、0.002 3和0.031 cm3·cm-3。植被类型为干旱草原,天然植被大部分已遭破坏,人工植被比例较高。人工林以小叶杨(Populussimonii)、沙柳为主的退化林和柠条、沙蒿为主的灌木林组成,在该地区生态环境中占据着比较重要的地位。本研究选取柠条和沙柳为研究对象,2种灌木的生长状况及形态特征如表1所示。
1.2 样品采集
1.2.1植物茎秆样品分别于2016年5月16日和8月22日进行样品的采集。采样点均选择在开阔、距离村落较远的地区,尽可能避免人为活动对同位素组分的影响。各选取3株形态特征相近的柠条和沙柳,每株每次取样1个。剪取阳面非绿色的栓化小枝(长4~5 cm,直径约0.3~0.5 cm),迅速去皮后装入采样瓶并用parafilm封口膜密封,放入冰盒,带回实验室后冷冻保存,用于测植物茎水的δD值。
1.2.2土壤样品在植物样地用土钻取0~100 cm的土壤样品,每10 cm取样1次,将土壤样品迅速装入取样瓶,用封口膜密封,并放入随身携带的冰盒,带回实验室冷冻保存,用于测土壤水中的δD值。
1.2.3大气降水样品降水量用自计式翻斗雨量筒(美国RainWise Wired)测定,精度0.25 mm,试验区气象站观测值作对比校正。用雨量筒承接单次雨量超过5 mm的降水作为样品,3次重复,采集时间是降水后10 min或夜间降水的次日清晨,以减少蒸发作用的影响[7]。水样用1.5 mL的特制玻璃瓶用封口膜密封后带回实验室冷藏保存(1~4 ℃),用于测定δ18O和δD。降水样品采集日期为2016年5月至9月,共采集到降水28次。
1.2.4植物叶片样品于2016年5~9月进行取样。分别选取长势良好、生长状况相似的柠条和沙柳3~5株,取树冠向阳面、成熟、无病虫害枝条中部的叶片进行混合取样。将采集的叶片在105 ℃杀青后,置于70 ℃恒温干燥箱中烘干48 h,再用粉碎机将其粉碎过80目筛制成供试样品,用于测定δ13C值,每个样品重复测定3~5次。
1.3 测定指标及方法
1.3.1样品同位素值(1)氢氧同位素值:所有水样的稳定性同位素的测定由西安理工大学水文循环实验室完成,先用低温真空抽提法抽提植物茎秆和土壤样品中的水分,抽提出的水分放在玻璃样品瓶中密封,并低温保存,然后采用美国LGR液态水同位素分析仪进行测定,测定精度:δ2H (δD)=0.5‰,δ18O =0.15‰。
δX(‰)=(Rsample/Rstandard-1)×100
式中Rsample是样品中元素的重轻同位素丰度之比,如(D/H)sample和(18O/16O)sample,Rstandard是国际通用标准物(H、O稳定同位素采用v-SMOW)稳定同位素丰度之比,如(D/H)standard和(18O/16O)standard。
(2)叶片碳同位素值:在西北农林科技大学水保所质谱仪分析室进行稳定碳同位素分析,称取处理好的样品3~5 mg,封入真空的燃烧管,并加入催化剂和氧化剂,燃烧产生的CO2经结晶纯化后,用DELTAplusXP质谱仪和FLASH EA固体分析仪(Thermo Fisher, Bremen, Germany)测定,精确度为±0.2‰。测定碳同位素的组成,以PDB(Pee Dee Belemnite)为标准,根据以下公式计算叶片的δ13C值[8]:


表1 2种灌木生长状况
其中,δ13C表示样品13C/12C与标准样品偏离的千分率,(13C/12C)PDB表示南卡罗来纳州白碚石(Pee Dee Belemnite)中的13C/12C。
1.3.2净光合速率、蒸腾速率和气孔导度在生长季,每月选择典型晴天,利用Li-6400便携式光合仪测定叶片净光合速率(Pn)、蒸腾速率(Tr)和气孔导度(Gs),分别选取3株生长良好的沙柳和柠条,在东西南北4个方向,每个方向取3片植物叶片,每张叶片连续读取5个稳定数据,频率是每隔2 h测定1次(7:00~19:00)。
1.3.3叶片水势叶水势用压力室(美国PMS 公司,1000型)进行测定。分别选取3株生长良好的沙柳和柠条,每样株选取生长良好无损伤的植物上部完全展开的成熟叶,用锋利的刀片沿叶柄垂直切割,之后去除叶柄基部小叶,迅速将叶轴插入压力室进行测定,当切口端有气泡冒出,即刻记录压力表显示值,频率是每隔2 h测定1次(5:00~19:00)。
1.4 数据分析
不同潜在水源如不同土层的土壤水对植物的贡献比例用IsoSource模型分析。计算时来源增量(source increment)设为1%,表示以1%的增量赋值植物对各水源的利用比例,用以检测可能的比例组合;质量平衡公差(mass balance tolerance)设为0.1%,表示各水源同位素值的差异不超过0.1%时,比例组合被认为是可能的组合。质量平衡公差一般不小于来源增量与各可能水源同位素值之间最大差值的乘积的1/2。采用Excel和SPSS统计软件对数据进行相关性分析。
2 结果与分析
2.1 植物生长季降水分布与温度的变化
由图1可知,研究区在柠条和沙柳生长季5~9月份的降水总量为569.1 mm。其中,<5 mm的降水事件有23次,5~10 mm的降水事件有13次,>10 mm的降水事件有15次;降水主要集中在7~8月,共425.3 mm,占整个生长季的74.7%。生长季内最高气温出现在6月29日,为26.29 ℃,生长季内平均气温为19.4 ℃。
2.2 土壤水和植物木质部水的δD特征
图2显示,2种植物立地土壤水的同位素值(δD)在旱季和雨季均以表层(0~10 cm)最大,且δD值随着土层深度的增加而逐渐降低。其中,沙柳立地旱季各土层土壤水δD值处于-42.98‰~-72.76‰之间,且80~100 cm与0~10 cm、10~40 cm土层间差异显著(P<0.05);而沙柳立地雨季相应土层土壤水δD值则处于-65.83‰~-78.19‰之间,除0~10 cm和10~40 cm土层间差异显著(P<0.05)外,其余各土层间差异均不显著(P>0.05)。同时,柠条立地各土层土壤水δD值在旱季处于-34.14‰~-73.72‰之间,且土层0~10 cm与40~60 cm、60~80 cm、80~100 cm间,以及10~40 cm与80~100 cm间差异均显著(P<0.05);而柠条立地雨季各土层土壤水δD值处于-66.53‰~-84.97‰之间,但各土层间差异不显著。

图2 2种植物立地土壤水δD值Fig.2 Change of δD of soil water with soil depth

图1 2016年植物生长季降水分布及同期气温的变化Fig.1 The variation of precipitation distribution and temperature during growth stage of two shrubs in 2016
从图3可知,沙柳植物茎秆水δD值在旱季和雨季分别为-56.62‰和-77.13‰,而柠条旱季和雨季植物茎秆水δD值分别为-57.79‰和-75.50‰。进一步通过比较发现,沙柳茎秆水δD值在旱季与60~80 cm土层土壤水δD值接近,在雨季与10~40 cm土层土壤水δD值接近;同时,柠条茎秆水δD值在旱季与40~60 cm土层土壤水δD值接近,在雨季与10~40 cm土层土壤水δD值接近。说明沙柳和柠条在旱季分别更倾向于利用60~80和40~60 cm土层的水分,而在雨季都更倾向于利用10~40 cm土层的水分。
进一步通过降水氢氧稳定同位素值用回归分析得出神木六道沟流域大气降水线方程:δD=7.69δ18O + 9.56(R2=0.97)(图4),比较其与全球大气降水线方程:δD=8δ18O + 10,发现区域大气降水线方程斜率和截距均偏小,降水的δ18O-δD坐标点基本落在区域大气降水线左边,但是土壤水及植物水δ18O-δD值都位于区域大气降水线的右侧,说明植物利用的水源氢氧同位素组成受到了二次蒸发的影响,从而发生了富集作用;而表层土壤水坐标点更远离区域大气降水线,又说明表层土壤水受蒸发影响较大。
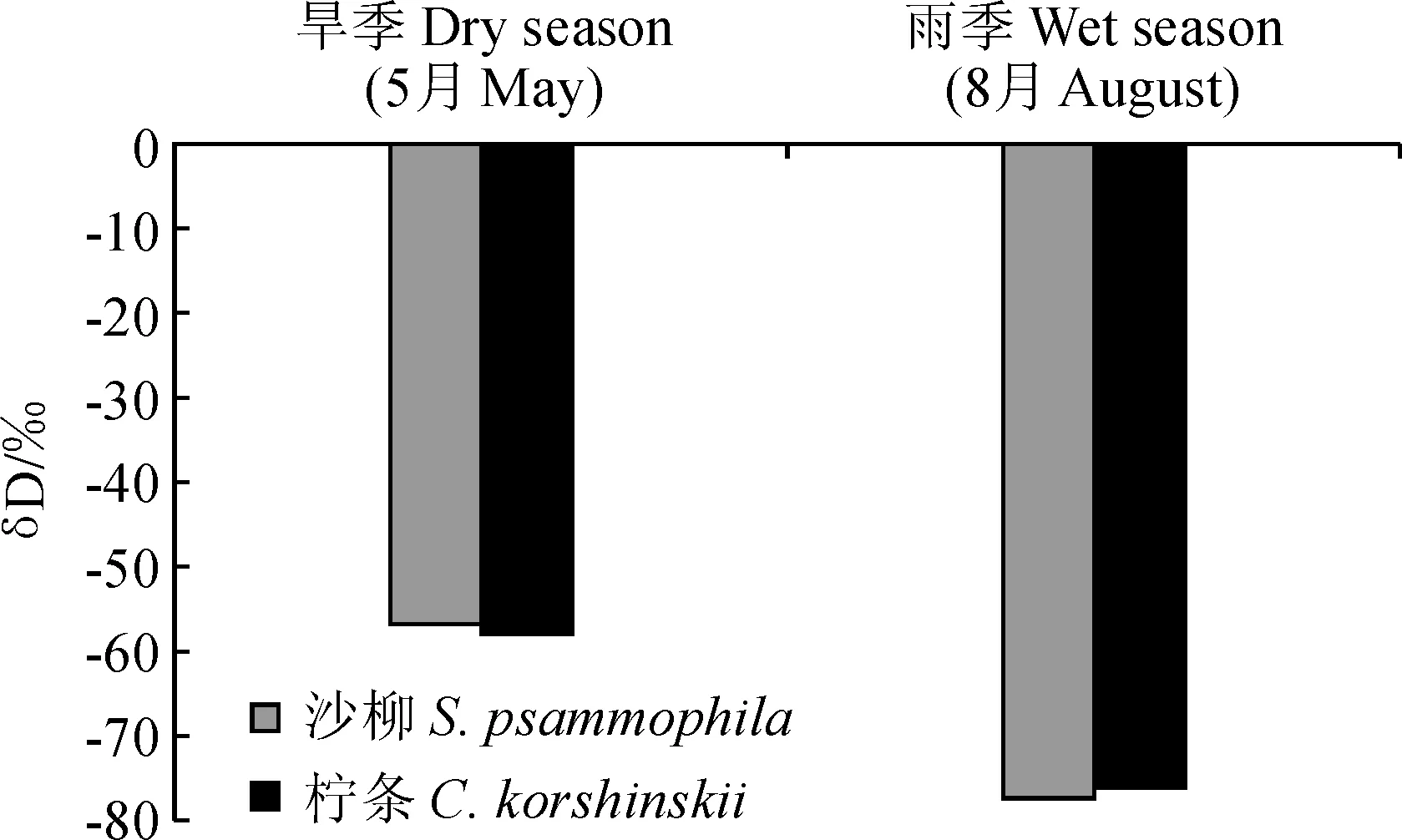
图3 旱季和雨季2种植物立地茎秆水δD值Fig.3 Change of δD of plant stem xylem water in dry season and wet season
2.3 2种灌木对不同土层水分利用的比例
运用IsoSource模型计算柠条和沙柳对各土层土壤水的可能利用比例。将0~100 cm土层分为5层即0~10、10~40、40~60、60~80和80~100 cm,其中0~10 cm为表层,10~60 cm为浅层,60~100 cm为深层。将植物木质部水和5层土壤水的δD值代入该模型,运行模型所得结果如表2所示。旱季柠条和沙柳对各层次土壤水分的利用比例接近,差异不明显。其中,柠条对40~60 cm土层土壤水分的利用率最高,比例达24.2%;其次为60~80 cm土层水分,利用比例为23.9%;而对0~10 cm、10~40 cm和80~100 cm土层土壤水的利用比例分别为13.6%、18.8%和19.4%。沙柳对60~80 cm土层土壤水分的利用率最高,比例达25.2%;其次为40~60 cm土层水分,利用比例为25.2%;而对0~10 cm、10~40 cm和80~100 cm土层土壤水的利用比例分别为12.8%、18.6%和19.5%。雨季2种植物对各层次土壤水分的利用表现出明显的不同。柠条利用0~10 cm土壤水分的比例最高(比例达36.8%),其次是10~40 cm土层的土壤水(利用率为20.7%),对其余各层次水分的利用比例接近,利用率为12.1%~16.2%;沙柳主要利用10~40 cm土层的土壤水,利用率高达76%,而对其他层次水分的利用很少(表2)。

图4 土壤水、植物木质部水的δ18O和δD及其与大气降水线关系Fig.4 δ18O and δD of soil water and plant stem xylem water and their relationship with meteoric water lines
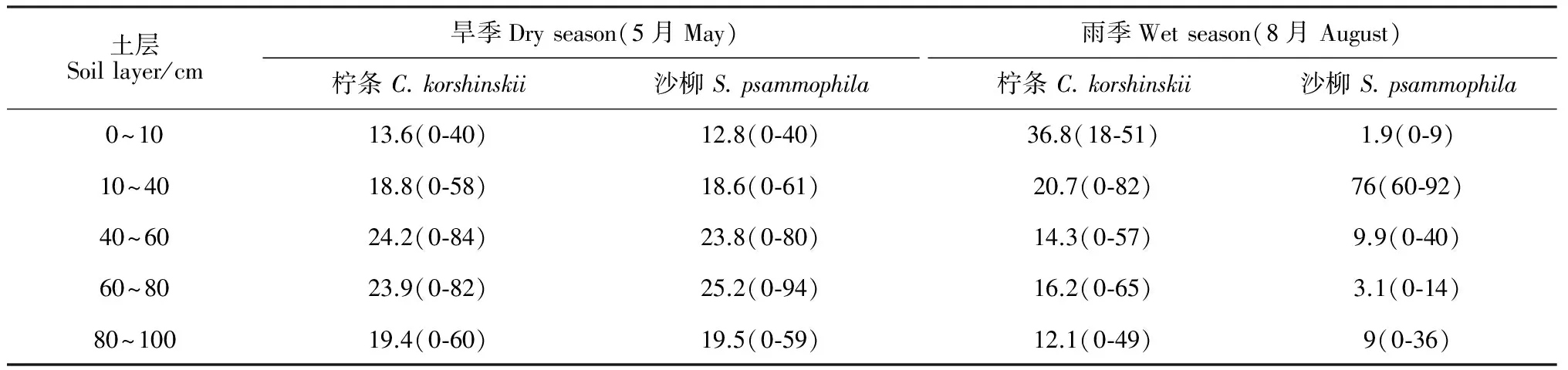
表2 不同季节2种灌木对各层次土壤水分的利用比例(最小值-最大值)
2.4 2种灌木叶片光合生理生态特征
首先,无论是在旱季还是雨季,柠条和沙柳叶片水势均呈单峰型变化,即从早晨到中午再到晚上,叶片水势呈先降低后升高的趋势,说明2种灌木在一天中其水分亏缺程度存在差异;同时,在总体上,旱季的叶片水势小于雨季,又说明旱季2种灌木的水分亏缺更为严重(图5,A)。
其次,柠条和沙柳气孔导度(Gs)在旱季和雨季均呈双峰型曲线变化(图5,B)。其中,从7:00~11:00呈上升趋势,并于11:00达到峰值,到13:00出现极小值,而后在15:00出现第二次峰值,随后呈下降趋势。柠条和沙柳在雨季的平均Gs分别比旱季高58.33%和41.67%。柠条雨季和旱季首次Gs峰值的平均值分别为0.25 和0.17 mol·m-2·s-1,而二次Gs峰值的平均值分别为0.22 和0.13 mol·m-2·s-1,雨季分别比旱季高47.06%和69.23%;沙柳雨季和旱季首次Gs峰值的平均值分别为0.23 和0.17 mol·m-2·s-1,二次峰值的平均值分别为0.19 和0.15 mol·m-2·s-1,雨季分别比旱季高出35.29%和26.67%。
再次,柠条和沙柳的光合速率(Pn)在旱季和雨季均呈双峰型曲线变化(图5,C)。其中,受气孔张开的影响,2种植物的Pn在旱季从7:00~11:00呈上升趋势,于11:00达到峰值,而在雨季从7:00~9:00光合速率呈上升趋势,于9:00达到峰值,旱季和雨季均在中午13:00出现极小值,而后在15:00出现第二次峰值,随后15:00~19:00呈下降趋势。柠条Pn在旱季和雨季的第一峰值分别为10.81 和14.30 μmol·m-2·s-1,第2峰值分别为10.00 和12.42 μmol·m-2·s-1,雨季分别比旱季高出32.28%和24.20%;沙柳Pn在旱季和雨季的第一峰值分别为9.60 和12.49 μmol·m-2·s-1,第二峰值分别为8.29 和11.18 umol·m-2·s-1,雨季分别比旱季高出30.10%和34.86%。
另外,柠条和沙柳的蒸腾速率(Tr)在旱季和雨季呈不规则变化(图5,D)。在旱季,柠条Tr变化呈双峰型曲线,而沙柳Tr变化呈不规则曲线,且变化平缓;在雨季,柠条和沙柳Tr变化均呈单峰型曲线。
以上结果说明在土壤水分条件相对较差的旱季,柠条和沙柳叶片水势较小,其通过降低气孔导度来减小水分的损失来维持自身的生长。柠条和沙柳的光合速率与蒸腾速率在旱季和雨季分别呈现不同的变化规律。
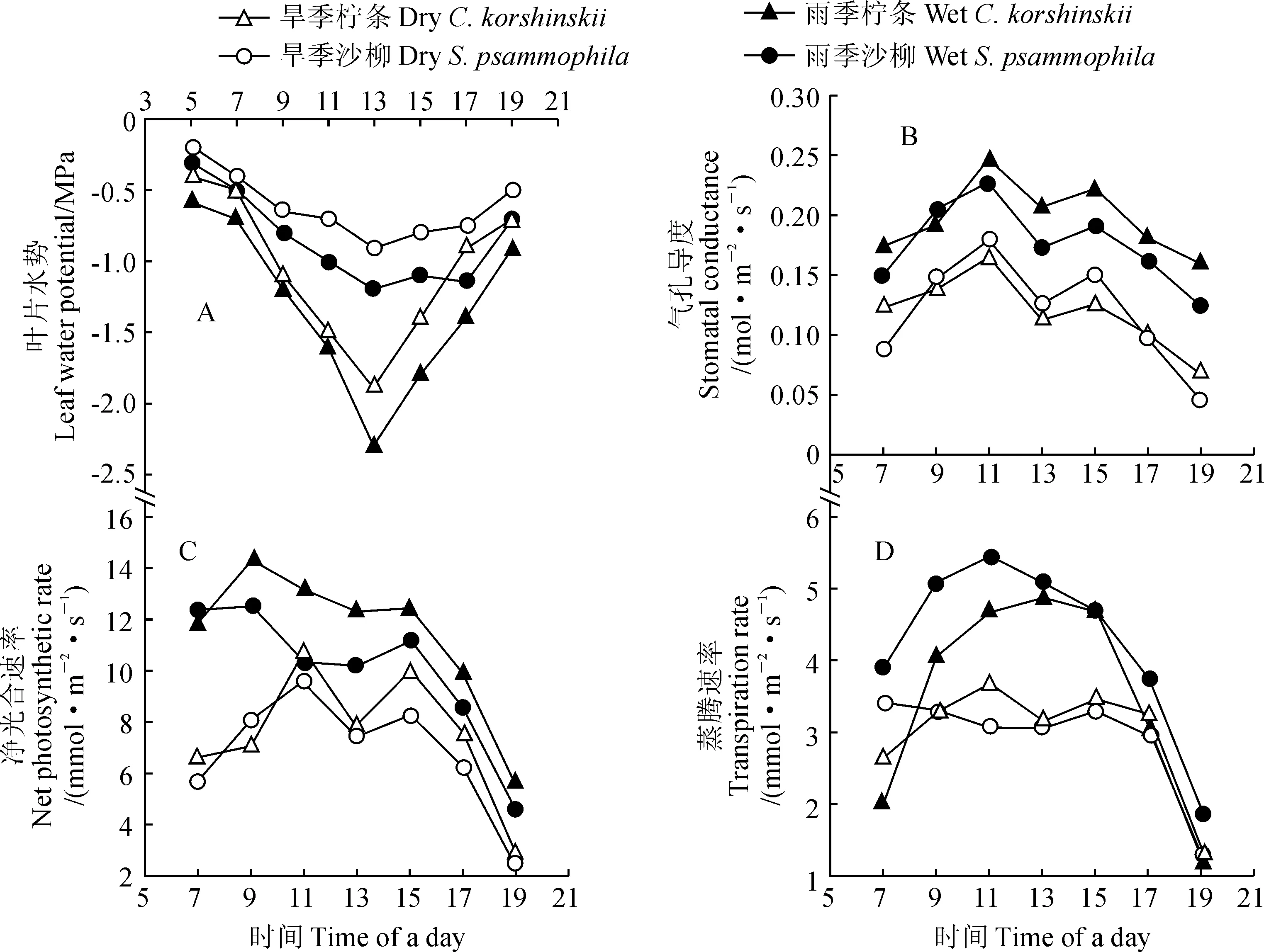
图5 柠条和沙柳叶片水势、气孔导度、光合速率和蒸腾速率的日变化Fig.5 Daily variations of leaf water potential, stomatal conductance, photosynthetic rate and transpiration rate of C. korshinskii and S. psammophila
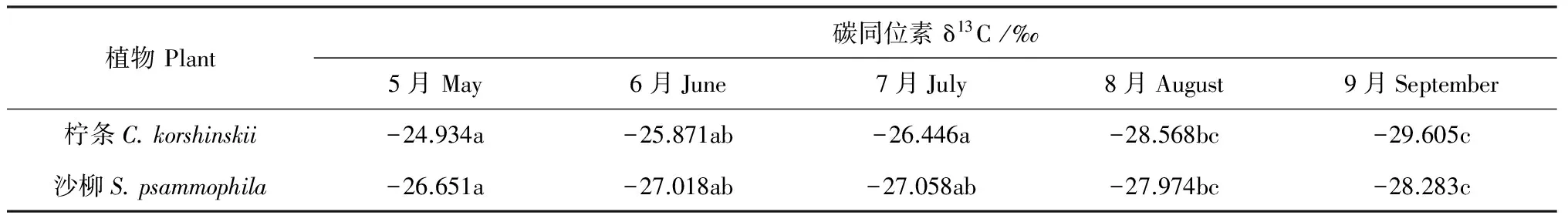
表3 2种灌木不同月份碳同位素值比较
注:数据后不同字母表示δ13C值在不同月份的Tukey多重比较结果在0.05水平上差异显著
Note:By the multiple comparison, δ13C at different height of different seasons with the different letters differ significantly at 0.05 level according to Tukey’s test
2.5 2种灌木叶片碳同位素组成比较
Deines[9]曾对全球陆生C3植物的δ13C值进行过统计,结果发现全球C3植物的δ13C值分布在-22.0‰~-34.0‰之间,其中90%左右植物的δ13C分布在-24.0‰~-29.0‰之间。由表3可知,本研究中的柠条和沙柳δ13C值的分布范围位于全球C3植物的δ13C范围之内。柠条和沙柳的δ13C值在5~9月份呈逐渐变轻的趋势,5月较重,分别为-24.934‰和-26.651‰;9月较轻,分别为-29.605‰和-28.283‰。柠条和沙柳不同月份之间的δ13C值存在差异,均具体表现为5、6、7月份之间差异不显著(P>0.05),5、6、7月份分别与8、9月份之间差异均极显著(P<0.01),而8月份与9月份之间差异不显著(P>0.05)。以上结果说明两灌木叶片δ13C值的季节变化模式呈现总体下降的趋势且各个月份之间δ13C值差异不同。
3 讨 论
3.1 2种灌木水分来源及生理生态适应性
植物生长于土壤之中,土壤水是植物水分的直接来源。植物根系吸收土壤水分的过程控制着水分状态以及光合作用等生命过程[10]。根系分布对植物功能的维持起着关键作用,尤其是处于风蚀水蚀区的六道沟流域在没有灌溉条件下植物仍然能够度过干旱时期,说明该区域植物可能通过根系获得稳定的水分来源。植物水分来源包括地下水、地表水、土壤水等,但是大气降水却是这些水分的原始来源[11]。旱季降水偏少,表层土壤蒸发强烈,水分散失,浅层土壤水分也随之减少,植被为了度过干旱季节,在表层和浅层土壤水分不足的条件下必然会利用深层土壤水。研究表明黄土高原地区柠条的根系主要密集于10~100 cm土层中[12]。沙柳为浅根系植物,其根系垂直分布一般不超过1.5 m,但水平根系分布可达冠幅的3倍左右,沙柳主要通过水平根扩展来获取浅层的土壤水[13]。本研究中,柠条和沙柳对0~100 cm土壤剖面各土层的土壤水均有利用,但对浅层(40~60 cm)和深层(60~80 cm)土壤水的利用较多。这是因为旱季柠条和沙柳仅靠利用表层(0~10 cm)土壤水不能保证其正常生长,为了度过旱季,必须依赖浅层(10~60 cm)和深层(60~100 cm)根系从土壤中吸收水分,因此较多地利用了40~60 cm和60~80 cm的土壤水,利用比例分别达24.2%、23.9%和23.8%、25.2%。
雨季来临后,2种植物的水分来源均有变化,柠条由旱季主要利用浅层(40~60 cm)和深层(60~80 cm)的土壤水变为主要利用表层(0~10 cm)和浅层(10~40 cm)的土壤水。沙柳由旱季主要利用浅层(40~60cm)和深层(60~80 cm)的土壤水变为主要利用浅层(10~40 cm)的土壤水。2种植物对表层和浅层土壤水分的利用比例明显增加,可能有两方面的原因:一方面是雨季中雨水补充了表层土壤水,使得表层土壤水分有效性增加;另一方面是土壤表层相对疏松,表层土壤中积累了一定量的由枯枝落叶形成的有机质,水分供给充足时,表层活性根的数量会大量增加[14],且表层根系吸收水分时消耗能量相对较少,植物优先利用表层水分[15]。徐庆等[16]对四川省炉霍县亚高山地区的野生植物种变叶海棠(Malustoringoides)及其伴生植物峨眉小檗(Berberisaemulans)的水分利用策略的研究表明,在旱季变叶海棠主要利用20~60 cm层次的土壤水,峨眉小檗主要利用40~60 cm层次的土壤水;在雨季变叶海棠转而利用0~40 cm的浅层土壤水,峨眉小檗的水分主要来源变为0~20 cm土层土壤水。Yang等[17]的研究也表明在降水少、表层土壤含水量低的旱季植物利用深层土壤水或地下水,在降水多、表层土壤含水量高的湿润季节转向利用浅层土壤水。在美国犹他州和亚利桑那州,Quercusgambelii在降水量比较少的地区利用的都是深层地下水,而在降水量最大的地区利用的则主要是浅层地下水[18]。
由旱季到雨季,2种植物的主要水分来源由深变浅,即水分利用策略随季节波动,当土壤水分出现亏缺时,植物显示出一定的忍耐能力,表现出不同的生理反应[19]。结合柠条和沙柳的各项生理生态指标来看,在旱季2种植物的叶片水势较低,说明水分亏缺较为严重,其气孔开张程度不如雨季大,光合速率和蒸腾速率也比雨季低,这可能是柠条和沙柳适应干旱环境的一种策略;另外,典型晴天正午,太阳辐射较强,蒸腾作用也较剧烈,2种植物采取关闭气孔的方式减少水分散失,从而有效地使其适应生境。孙守家等[20]在研究太行山南麓山区栓皮栎(Quercusvariabilis)-扁担杆(Grewiabiloba)生态系统水分利用策略时也发现,植物会通过深水源及部分气孔关闭和浅水源及低蒸腾耗散的水分利用策略来利用旱季中有限的水分。张佩等[21]在研究黑河中游荒漠绿洲过渡带多枝柽柳(Tamarixramosissima)时同样发现,当水分条件更差时,植物通过调节气孔维持相对稳定的碳同化能力及较高水分利用效率,表现出较好的适应能力。
3.2 2种灌木的水分利用效率
不同光合途径(C3、C4和CAM)因光合羧化酶(RuBP和PEP羧化酶)和发生羧化的时间和空间上的差异对13C有不同的识别和排斥,导致不同光合途径的植物具有显著不同的δ13C值。据此,δ13C值可以用于区分植物的C3和C4光合途径。在陆生植物中,C3植物δ13C值变化在-20‰~-35‰(平均为-26‰),C4植物变化在-7‰~-15‰(平均为-12‰),而CAM植物则大约介于上述2类植物之间,一般变化在-10‰~-22‰之间(平均为-16‰)[22]。根据柠条δ13C值变化范围为(-29.60±0.90)‰~(-24.93±0.92)‰,沙柳δ13C值变化范围为(-28.28±0.45)‰~(-26.65±0.17)‰,说明柠条和沙柳属于C3植物。
对于C3植物而言,叶片的δ13C值能指示植物的长期水分利用效率:植物的δ13C值越小,其水分利用效率越低[8,23-24],一些学者通过试验也证明了用植物叶片的δ13C值来指示植物长期水分利用效率的可靠性[25],而水分利用效率又是植物抗旱性能的重要指标之一,通常水分利用效率较高的植物,其抗旱性较强;抗旱性弱的植物,其水分利用效率也较低[26]。本研究中整个生长季柠条和沙柳δ13C值平均值分别为-27.08‰和-27.40‰,柠条δ13C值略大于沙柳,即水分利用效率柠条略高于沙柳。沙柳和柠条在旱季δ13C值均大于雨季,说明2种灌木的水分利用效率旱季大于雨季。降水作为该区柠条和沙柳最主要的水分来源,旱季降水较少,土壤含水量较低,2种植物可利用的水分较少,可能会导致柠条和沙柳受到一定程度的水分胁迫,因此它们通过提高水分利用效率来应对干旱的胁迫。雨季降水较旱季多,土壤含水量高,植物可利用的水分较充足,柠条和沙柳受到相对较小的水分胁迫,其δ13C值与旱季相比较小即水分利用效率较低。刘丽颖等[27]通过对共和盆地不同林龄乌柳林(SalixcheilophilaSchneid.)的水分利用策略研究表明,旱季5a生乌柳由于水分来源较9a和25a生乌柳的水分来源少,因此采取提高水分利用效率的方法来适应干旱胁迫,而9a和25a生乌柳受到相对较小的水分胁迫,水分利用效率相对较低。
柠条和沙柳叶片δ13C值随生长季而减小的规律可能与叶片成熟度和其稳定碳同位素自然丰度有关。在生长初期,细胞生长比较活跃,以便合成大量有机质来满足植物叶片发育和植物构建的需要,相对而言,细胞内部CO2浓度处于“饥饿”状态,因而导致植物对δ13C值的识别和排斥降低,故植物叶片δ13C值较高。到了生长后期,叶片外部形态和内部结构发育趋于完全,内部生理代谢功能趋于完备,植物具备了较完善生理生化反应调控机制,能较有效地识别与排斥δ13C值,故植物叶片δ13C值较低。另外,可能也与空气温湿度升高有关。Farquhar等[28]研究发现随着空气温湿度的升高,植物蒸发增强,气孔打开,引起植物叶片内部CO2浓度升高,如果此时植物光合作用速率仍维持正常水平,植物对CO2的识别能力势必增强,从而使叶片碳同位素值降低。Garten等[29]、严昌荣等[30]和刘海燕等[31]分别对沼生栎(Quercusprinus)、暖温带落叶乔木和灌木沙柳叶片δ13C值的研究中也发现从生长初期到生长末期δ13C值有逐渐降低的趋势。
4 结 论
(1)柠条和沙柳的水分利用策略随季节变化,不同季节对不同土壤深度水分的利用比例存在差异。旱季柠条和沙柳均利用深层土壤水分,其中柠条对40~80 cm土层水分的利用率达48.1%,沙柳对40~80 cm土层水分的利用率达49%。雨季2种植物对各层次土壤水分的利用发生了明显的变化,表现为减少对深层土壤水分的利用,更倾向于利用浅层土壤水分,其中柠条更多的利用表层0~10 cm和浅层10~40 cm的土壤水分,利用率约为57.5%;沙柳利用浅层10~40 cm的土壤水,利用比例达76%。两种植物在旱季和雨季通过不同策略合理利用水源维持自身生长,使其组成的生态系统更好地适应风蚀水蚀区脆弱的生态环境。
(2)在整个生长季,柠条叶片δ13C值在(-29.60±0.90)‰~(-24.93±0.92)‰之间,平均值为-27.08‰;沙柳δ13C值变化范围(-28.28±0.45)‰~(-26.65±0.17)‰,平均值为-27.40‰。柠条δ13C值略大于沙柳,即长期水分利用效率表现为柠条略高于沙柳。两灌木叶片δ13C值的季节变化模式呈现总体下降的趋势。最高值出现在5月,最低值出现在9月。在土壤水分条件相对较差的旱季,柠条和沙柳叶片水势较小,其能通过降低气孔导度减小水分损失来获得较高的碳同化速率,维持较高的水分利用效率来适应干旱环境。
参考文献:
[1]唐克丽. 黄土高原水蚀风蚀交错区治理的重要性与紧迫性[J]. 中国水土保持, 2000, (11): 11-12.
TANG K L. Importance and urgency of harnessing the interlocked area with both water and wind erosion in the loess plateau [J].SoilandWaterConservationinChina, 2000, (11): 11-12.
[2]LI S G, ROMERO-SALTOS H, TSUJIMURA M,etal. Plant water sources in the cold semiarid ecosystem of the upper Kherlen River catchment in Mongolia: A stable isotope approach [J].JournalofHydrology, 2007,333(1): 109-117.
[3]DAI Y, ZHENG X J, TANG L S,etal. Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon, species in the Gurbantonggut Desert [J].PlantandSoil, 2015,389(1): 73-87.
[4]朱建佳, 陈辉, 邢星, 等. 柴达木盆地荒漠植物水分来源定量研究—以格尔木样区为例[J]. 地理研究, 2015,34(2): 285-292.
ZHU J J, CHEN H, XING X,etal. Quantification analysis of water sources of desert plants in Qaidam Basin: A case study of Golmud plot [J].GeographicalResearch, 2015,34(2): 285-292.
[5]丁亚丽, 陈洪松, 聂云鹏, 等. 基于稳定同位素的喀斯特坡地尾巨桉水分利用特征[J]. 应用生态学报, 2016,27(9): 2 729-2 736.
DING Y L, CHEN H S, NIE Y P,etal. Water use strategy ofEucalyptusurophylla×E.grandison karst hillslope based on isotope analysis [J].ChineseJournalofAppliedEcology, 2016,27(9): 2 729-2 736.
[6]查轩, 唐克丽. 水蚀风蚀交错带小流域生态环境综合治理模式研究[J]. 自然资源学报, 2000,15(1): 97-100.
CHA X, TANG K L. Study on comprehensive control model of small watershed ecoenvironment in water and wind crisscrossed erosion zone [J].JournalofNaturalResources, 2000,15(1): 97-100.
[7]林光辉. 稳定性同位素生态学[M]. 北京: 高等教育出版社, 2013.
[8]FARQUHAR G D, O’LEARY M H, BERRY J A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves [J].AustralianJournalofPlantPhysiology, 1982,13(2): 281-292.
[9]DEINES P. Chapter 9-the isotopic composition of reduced organic carbon [J].TerrestrialEnvironmentA, 1980,1: 329-406.
[10]MACFALL J S, JOHNSON G A, KRAMER P J. Comparative water uptake by roots of different ages in seedlings of loblolly pine (PinustaedaL.) [J].NewPhytologist, 1991,119(4): 551-560.
[11]曹燕丽, 卢琦, 林光辉. 氢稳定性同位素确定植物水源的应用与前景[J]. 生态学报, 2002,22(1): 111-117.
CAO Y L, LU Q, LIN G H. Review and perspective on hydrogen stable isotopes technique in tracing plant water sources researches [J].ActaEcologicaSinica, 2002,22(1): 111-117.
[12]牛西午. 柠条生物学特性研究[J]. 华北农学报, 1998,13(4): 122-129.
NIU X W. Biological characters of cultivars inCaragana[J].ActaAgriculturaeBoreali-Sinica, 1998,13(4): 122-129.
[13]刘健, 贺晓, 包海龙, 等. 毛乌素沙地沙柳细根分布规律及与土壤水分分布的关系[J]. 中国沙漠, 2010,30(6): 1 362-1 366.
LIU J, HE X, BAO H L,etal. Distribution of fine roots ofSalixpsammophilaand its relationship with soil moisture in Mu Us Sandland [J].JournalofDesertResearch, 2010,30(6): 1 362-1 366.
[14]SUN S J, MENG P, ZHANG J S,etal. Variation in soil water uptake and its effect on plant water status inJuglansregiaL. during dry and wet seasons [J].TreePhysiology, 2011,31(12): 1 378-1 389.
[15]SCHENK H J. The shallowest possible water extraction profile: a null model for global root distributions [J].VadoseZoneJournal, 2008,7(3): 1 119-1 124.
[16]徐庆, 王海英, 刘世荣. 变叶海棠及其伴生植物峨眉小檗的水分利用策略[J]. 生态学报, 2011,31(19): 5 702-5 710.
XU Q, WANG H Y, LIU S R. Water use strategies ofMalustoringoidesand its accompanying plant speciesBerberisaemulans[J].ActaEcologicaSinica, 2011,31(19):5 702-5 710.
[17]YANG H, AUERSWALD K, BAI Y F,etal. Complementarity in water sources among dominant species in typical steppe ecosystems of Inner Mongolia, China [J].PlantandSoil, 2011,340(1): 303-313.
[18]WILLIAMS D G, EHLERINGER J R. Intra-and interspecific variation for summer precipitation use in pinyon-juniper woodlands [J].EcologicalMonographs, 2000,70(4): 517-537.
[19]严昌荣, 韩兴国. 六种木本植物水分利用效率和其小生境关系研究[J]. 生态学报, 2001,21(11): 1 952-1 957.
YAN C R, HAN X G. Water use efficiency of six woody species in relation to micro-environmental factors of different habitats[J].ActaEcologicaSinica, 2001,21(11): 1 952-1 957.
[20]孙守家, 孟平, 张劲松, 等. 太行山南麓山区栓皮栎-扁担杆生态系统水分利用策略[J]. 生态学报, 2014,34(21): 6 317-6 325.
SUN S J, MENG P, ZHANG J S,etal. Ecosystems water use patterns ofQuercusvariabilisandGrewiabilobabased on stable hydrogen and oxygen isotopes in the south aspect of Taihang Mountains[J].ActaEcologicaSinica, 2014,34(21): 6 317-6 325.
[21]张佩, 袁国富, 庄伟, 等. 黑河中游荒漠绿洲过渡带多枝柽柳对地下水位变化的生理生态响应与适应[J]. 生态学报, 2011,31(22): 6 677-6 687.
ZHANG P, YUAN G F, ZHUANG W,etal. Ecophysiological responses and adaptation ofTamarixramosissimato changes in groundwater depth in the Heihe river basin[J].ActaEcologicaSinica, 2011,31(22): 6 677-6 687.
[22]HALL A E, RICHARDS R A, CONDON A G,etal. Carbon isotope discrimination and plant breeding [J].PlantBreedingReviews, 2010,12: 81-113.
[23]FARQUHAR G D, RICHARDS R A. Isotopic composition of plant carbon correlates with water-use efficiency of wheat genotypes [J].FunctionalPlantBiology, 1984,11(6): 539-552.
[24]张怡, 罗晓芳, 沈应柏. 干旱胁迫下四倍体刺槐幼苗水分利用效率及稳定碳同位素组成的研究[J]. 西北植物学报, 2009,29(7): 1 460-1 464.
ZHANG Y, LUO X F, SHEN Y B. Water use efficiency and carbon isotopic compositions inTetraploidClonesofBlackLocustunder drought resistance [J].ActaBot.Boreal.-Occident.Sin, 2009,29(7): 1 460-1 464.
[25]樊廷录, 马明生, 王淑英, 等. 不同基因型冬小麦旗叶的稳定碳同位素比值及其与产量和水分利用效率的关系[J]. 植物生态学报, 2011,35(2): 203-213.
FAN T L, MA M S, WANG S Y,etal. Stable carbon isotope ratio (δ13C) in flag leaves of different genotypes of winter wheat and its relation to yield and water use efficiency [J].ChineseJournalofPlantEcology, 2011,35(2): 203-213.
[26]潘佳, 李荣, 胡小文. 水分条件对红砂叶片碳同位素组成与光合特性和分枝生长的影响[J]. 西北植物学报, 2016,36(6): 1 190-1 198.
PAN J, LI R, HU X W. Effect of water conditions on carbon isotope composition, photosynthesis and branch growth ofReaumuriasoongorica[J].ActaBot.Boreal.-Occident.Sin, 2016,36(6): 1 190-1 198.
[27]刘丽颖, 贾志清, 朱雅娟, 等. 青海共和盆地不同林龄乌柳林的水分利用策略[J]. 林业科学研究, 2012,25(5): 597-603.
LIU L Y, JIA Z Q, ZHU Y J,etal. Water use strategy ofSalixcheilophilastands with different ages in Gonghe basin, Qinghai Province [J].ForestResearch, 2012,25(5): 597-603.
[28]FARQUHAR G D, And J R E, HUBICK K T. Carbon isotope discrimination and photosynthesis [J].AnnualReviewofPlantPhysiologyandPlantMolecularBiology, 2003,40(40): 503-537.
[29]GARTEN JR C T, TAYLOR JR G E. FOLIAR δ13C within a temperate deciduous forest: spatial, temporal, and species sources of variation [J].Oecologia, 1992,90(1): 1-7.
[30]严昌荣, 韩兴国, 陈灵芝, 等. 暖温带落叶阔叶林主要植物叶片中δ13C值的种间差异及时空变化[J]. 植物学报, 1998,40(9): 853-859.
YAN C R, HAN X G, CHEN L Z,etal. Foliar δ13C within temperate deciduous forest: its spatial change and interspecies variation [J].ActaBotanicaSinica, 1998,40(9): 853-859.
[31]刘海燕, 李吉跃, 赵燕, 等. 沙柳稳定碳同位素值的特点及其水分利用效率[J]. 干旱区研究, 2008,25(4): 514-518.
LIU H Y, LI J Y, ZHAO Y,etal. Study on δ13C values of desert plantSalixpsammophilaand water use efficiency [J].AridZoneResearch, 2008,25(4): 514-518.
