新疆天山野果林准噶尔山楂种群结构与动态分析
2018-02-13吕海英艾克拜尔依米提
杨 蕾,吕海英,李 进,马 雪,艾克拜尔·依米提
(新疆特殊环境物种保护与调控生物学实验室,干旱区植物逆境生物学重点实验室,新疆师范大学 生命科学学院,乌鲁木齐 830054)
种群结构分析是植物生态学研究的核心问题之一[1-2],它能够反映出种群的数量动态、未来发展趋势以及种群与环境之间的相互关系[3-4]。大多数研究在种群数量统计中通常采用静态生命表,直观地呈现出种群各年龄段实际存活数和死亡数,以生存分析函数进一步阐明种群各生长阶段变化规律和更新策略[5-6]。时间序列模型可以从不同空间和时间的变化中揭示种群建立与发展的特点,预测种群未来发展趋势[7]。因此,种群结构与种群动态分析能够为野生濒危植物种群的保护、管理及生态恢复提供重要的理论指导意义。
准噶尔山楂(Crataegussongarica)为蔷薇科山楂属光核组唯一一种植物,在中国仅分布于新疆伊犁霍城县大、小西沟、果子沟以及伊宁县吉尔格朗沟等山区,现已被录入中国优先保护物种名录,是国家三级重点保护野生植物、新疆特有珍稀濒危植物[8-9]。在新疆天山野果林中丰富的植被资源,是多种栽培果树的野生种质起源地,其中准噶尔山楂具有较高的营养价值与药用价值[10-13],可用作栽培果树的砧木和育种材料,利用价值极高。此外,准噶尔山楂作为天山野果林的主要组成树种之一,对于当地的水土保持、生态系统稳定以及中亚天山野果林生物多样性的保护研究具有重要意义[14-15]。但根据调查显示天山野果林的实际现存总面积相比50年前缩减近30%[16],在恶劣的气候变化加之大量的人为干扰下,多种野生植物资源数量缩减,实际面积逐年递减。而鉴于天山野果林生态系统的特殊性和重要性,对天然分布范围极为狭小的准噶尔山楂进行系统地种群调查显得尤为重要,研究中从年龄结构、静态生命表和生存分析等方面对准噶尔山楂种群结构特征进行分析,并结合时间序列预测种群未来发展趋势,为准噶尔山楂种群的合理开发与管理采取保护措施,并为生态系统恢复提供重要的理论指导[14,17]。
1 研究区概况
研究区位于新疆伊犁霍城县大西沟境内,地处天山野果林带最东端81°31′~80°40′E,40°30′~44°35′N,为温带湿润型气候,太阳年辐射总量在5 600~5 800 MJ/m2,年日照时数3 150 h,年均降雨量约500 mm左右,冬季处于逆温层内,逆温层分布海拔上限可达1 700~2 300 m,最大的逆温强度为0.4 ℃/100 m[18]。大西沟地处山地草原带的深切峡谷,是伊犁谷地中最为温和、适宜和赋有“海洋性”气候特色的地段[19-20],主要生长着喜温暖湿润的落叶阔叶树种,多以混交林存在[18,20]。其中与准噶尔山楂种群伴生的落叶阔叶乔木包括有野生樱桃李(Prunusdivaricata)、野山杏(Armeniacavulgaris)、新疆野苹果(Malussieversii)等,树下灌木主要有野蔷薇(Rosamultiflora)、黑果悬钩(Rubuscaesius)、截萼忍冬(Loniceraaltmannii)、天山樱桃(CerasustianschanicaPojark.)、异果小檗(Berberisheteropoda)等,草本植物主要有小飞蓬(Erigeroncanadensis)、大羊茅(Festucagigantea)、荨麻(Urticafissa)、短柄草(Brachypodiumsylvaticum)、短距凤仙花(Impatiensbrachycentra)、水金凤(Impatiensnoli-tangere)等。
2 研究方法
2.1 野外调查方法
2016年和2017年7月分别在霍城县大西沟境内进行2次野外实际踏查,通过采用典型样地法,选取准噶尔山楂种群分布集中且林分保持较为良好的地段设置21个50 m×50 m大样方,共计面积5.25 hm2,样地设置如图1所示。对样方内准噶尔山楂植株进行逐木测量,利用卷尺、游标卡尺等测量工具统计植株胸径(DBH≥2 cm)、树高、冠幅、枝下高以及幼苗(DBH<2 cm)基径、株高、个体数,并通过GPS记录样地的海拔、坡度等生境特征。

图1 新疆天山野果林准噶尔山楂种群采样点示意图Fig.1 The population samples of Crataegus songarica populations of Tianshan Wild Fruit Forest in Xinjiang
2.2 种群径级划分和静态生命表编制
根据2015至2018年期间实验室盆栽试验结果,发现2年生实生幼苗最大基径不超过1 cm,进而参考其他径级划分方法[21],结合野外实际数据,将准噶尔山楂种群共划分为13个胸径级,小于2 cm为第1径级,2~5 cm为第2径级,5~8 cm为第3径级,依此类推,胸径大于35 cm全部归为最大级,即第13径级。运用“空间代替时间”的方法确定准噶尔山楂种群年龄结构,相应径级对应相应龄级,如第1径级对应Ⅰ龄级[19]。其中Ⅰ龄级为幼苗(DBH=0~2 cm),Ⅱ~Ⅲ龄级为幼树(DBH=2~8 cm),Ⅲ~Ⅷ龄级为成体树(DBH=8~20 cm),Ⅷ龄级后的为老龄阶段(DBH>20 cm)。
准噶尔山楂种群静态生命表的参数值:x为龄级;ax表示在x龄级内现有个体数;ax′为匀滑修正后在x龄级内的现有个体数;lx表示在x龄级开始时标准化存活个体数(以1 000为基数),lx=(ax/a0)×1 000;dx是从x到x+1龄级间隔期内标准化死亡个体数,dx=lx-lx+1;qx是从x到x+1龄级间隔期间的死亡率,qx=dx/lx;Lx为从x到x+1龄级间隔期间还存活的个体数,Lx=(lx+lx+1)/2;Tx是从x龄级到超过x龄级的总个体数,Tx=∑Lx;ex是进入x龄级个体的期望寿命,ex=Tx/lx;Kx是消失率,Kx=lnlx- lnlx+1。在编制种群静态生命表过程中,各径级个体数量相差较大使生命表中出现死亡率为负值的情况,这不能够满足编制静态生命表的前提条件,对此本实验对准噶尔山楂种群的调查数据采用了匀滑技术处理[22-23]。
2.3 种群年龄结构的数量变化动态(Vpi)分析
根据准噶尔山楂种群静态生命表中的数据,采用陈晓德[24]的量化方法定量描述准噶尔山楂数量种群动态变化。具体方法如下:
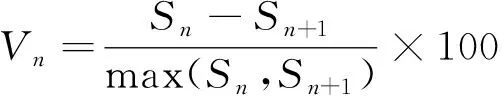
式中:Vn表示种群从n到n+1级的个体数量变化动态,Vpi表示整个种群结构的数量变化动态指数,Sn、Sn+1分别表示第n、n+1年龄级种群个体数。
当考虑外部干扰时:
式中:K表示种群年龄级数量,Vpi与Vn为正、零、负值的意义分别反映相邻年龄级个体数量或整个种群增长、稳定、衰退的结构动态关系。P为种群对外界干扰所承担风险的概率,只有当P值为最大时才对种群动态Vpi构成最大影响。
2.4 生存分析
在准噶尔山楂种群生命表的基础上,引入生存分析中的生存率函数(Si)、累计死亡率函数(Fi)、死亡密度函数(fi)以及危险率函数(λi)其公式如下[25-26]:
Si=P1×P2×P3…Pi(Pi为存活率)
Fi=1-Si
fi=(Si-1-Si)/h(h为龄级宽度)
λi=2qi/hi(1+Pi)(qi为死亡率)
根据以上4个函数的估算值绘制生存率曲线、积累死亡率曲线、死亡密度曲线和危险率曲线。
2.5 种群动态的时间序列预测模型
选用一次性移动平均法对准噶尔山楂种群的龄级结构进行模拟和预测[27-28]。
式中:n表示需要预测的未来时间年限,Mt表示未来n年t龄级的种群大小,XK为当前K龄级的种群大小。本研究分别对2年、4年、6年、8年以及10年龄级时间后准噶尔山楂种群数量进行时间序列预测。
3 结果与分析
3.1 静态生命表分析
将所有样地个体存活数经匀滑修正后得到ax′,据此编制准噶尔山楂种群静态生命表(表1)。由表1可以看出,在5.25 hm2的大面积调查中,准噶尔山楂种群共计1 926株,最大径级为13,对应最大DBH为35.03 cm,其中Ⅰ龄级个体达1 262株,具备丰富的幼苗数量,Ⅲ龄级至Ⅵ龄级各龄级数量在90株左右,从Ⅶ龄级开始各龄级数量均不足50株,整体数量偏少;Ⅱ~Ⅳ龄级准噶尔山楂种群处于生理活动旺盛期,Ⅲ龄级的ex达到最大值4.952 ,从Ⅸ龄级开始ex值逐渐递减、死亡率升高,Ⅻ龄级死亡率高达0.778,除了与准噶尔山楂自身生理特性有关外,种群所经历的较强环境压力(Kx=1.505)以及种群自疏效应共同影响着种群的数量分布。
3.2 年龄结构分析
以龄级为横坐标,以静态生命表中各龄级存活株数(ax)的百分比为纵坐标,绘制准噶尔山楂种群年龄结构图。由图2可以看出,准噶尔山楂种群各径级植株数量随径级的增加而逐渐减少,第1径级个体数量最多,占总数的65.5%,随后在第2径级数量发生骤减,总比例降至6.5%;种群内成体植株的各径级数量偏少,第3~8径级共占总数的22.2%;从第8径级之后,准噶尔山楂种群数量开始持续下降,进入老龄化阶段。准噶尔山楂种群总体表现为幼龄个体居多,中龄、老龄个体逐级递减的倒“J”型分布,属于增长型年龄结构。
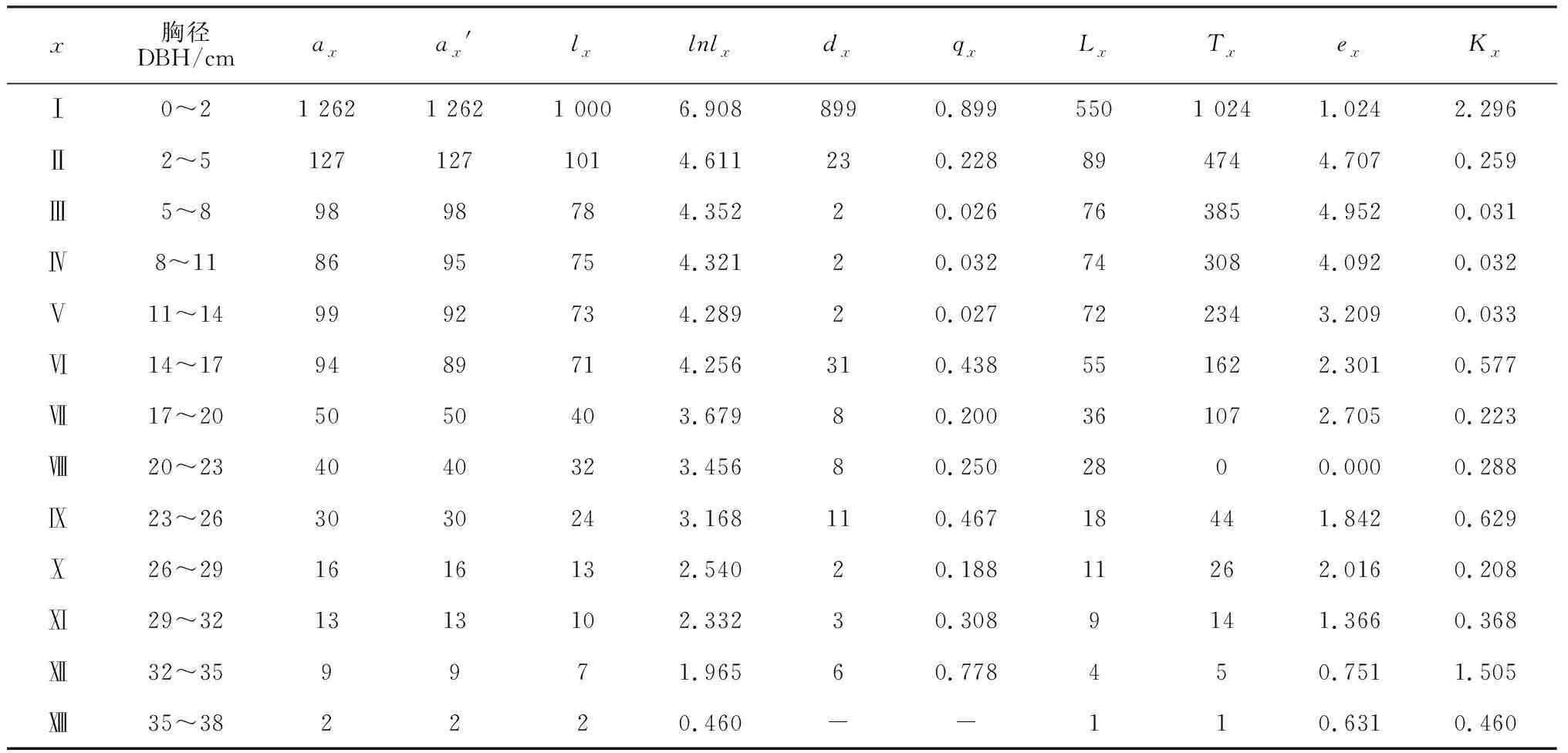
表1 新疆天山野果林准噶尔山楂种群的静态生命表
注:x为龄级;ax表示在x龄级内现有个体数;ax′为匀滑修正后在x龄级内的现有个体数;lx表示在x龄级开始时标准化存活个体数(以1 000为基数),lx=(ax/a0)×1 000;dx是从x到x+1龄级间隔期内标准化死亡个体数,dx=lx-lx+1;qx是从x到x+1龄级间隔期间的死亡率,qx=dx/lx;Lx为从x到x+1龄级间隔期间还存活的个体数,Lx=(lx+lx+1)/2;Tx是从x龄级到超过x龄级的总个体数,Tx=∑Lx;ex是进入x龄级个体的期望寿命,ex=Tx/lx;Kx是消失率,Kx=lnlx- lnlx+1
Note:x. Age class;ax. Number of survival individuals in agex;ax′. Number of survival individuals after smoothing amend in agex;lx.Proportion individuals surviving from the beginning to agex;dx.Number of dead individuals from agextox+1;qx.Mortality rate fromxtox+1;Lx.Mean number of survival individuals fromxtox+1;Tx.Total number of survival individuals fromxtox+1;ex.Life expectancy at agex;Kx.Age specific mortality
3.3 存活曲线分析
存活曲线能够直观反映出种群个体在各年龄级的存活状况,据此本研究将准噶尔山楂种群原始数据整理绘制存活曲线(图3),将准噶尔山楂种群存活曲线与经典存活曲线相对照,对比结果显示该种群幼年阶段死亡率极高,存活曲线介于Deevey-Ⅱ和Ⅲ型之间。为进一步确定准噶尔山楂种群存活曲线类型,本研究根据数学模型检验估算,用指数方程式:Nx=N0e-bx描述Deevey-Ⅱ型存活曲线,以幂函数式:Nx=N0x-b描述Deevey-Ⅲ型存活曲线[31]。模型检验(表2)结果显示,准噶尔山楂种群模型的F检验值和判定系数R2值中指数函数均大于幂函数,因此推定准噶尔山楂种群存活曲线表现为Deevey-Ⅱ。
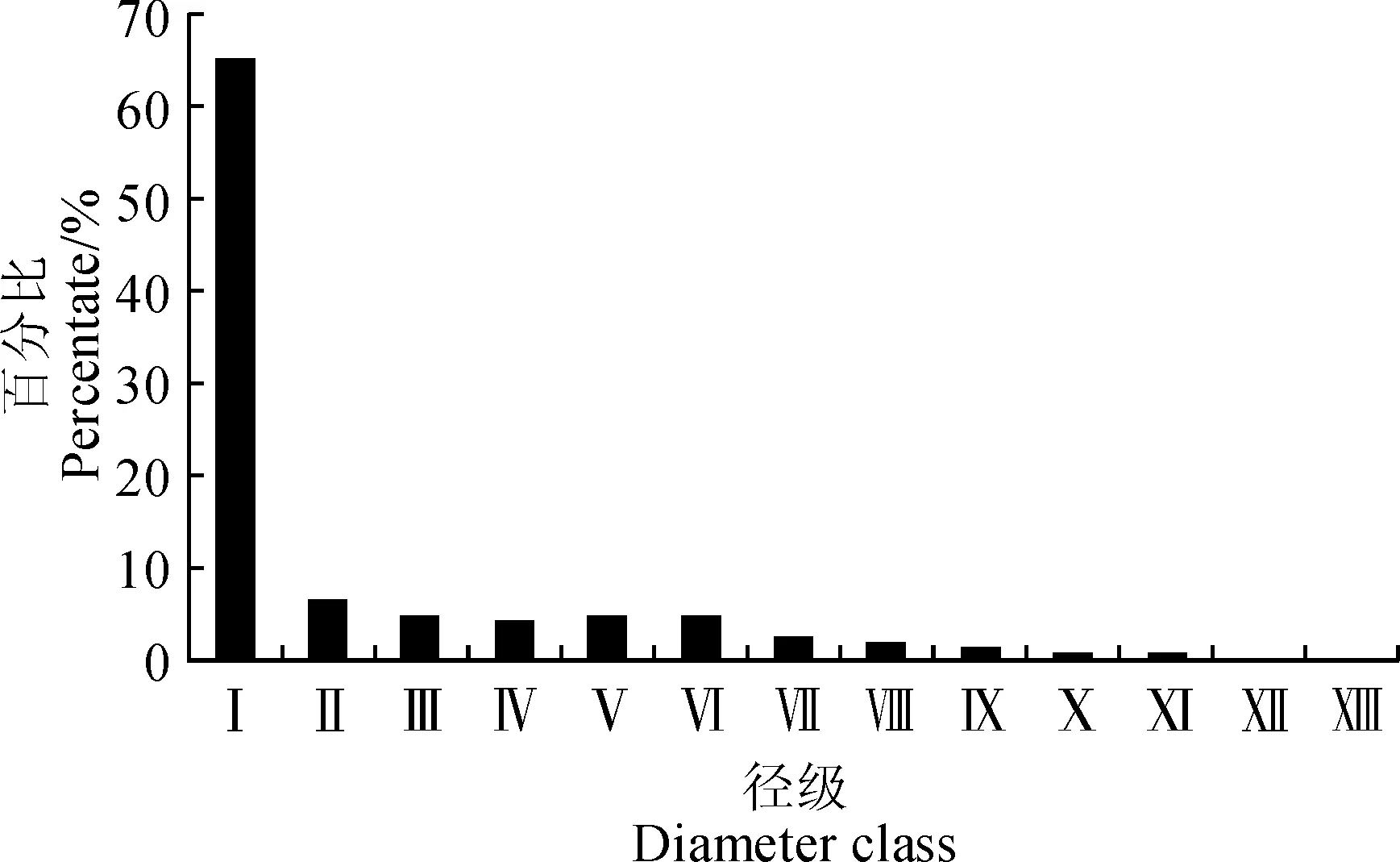
Ⅰ~.径级(代替龄级);DBH .胸径;Ⅰ.DBH=2 cm; Ⅱ. 2 cm

图3 新疆天山野果林准噶尔山楂种群的存活曲线Fig.3 Survival curve of C. songarica populations in Tianshan Wild Fruit Forest of Xinjiang
3.4 死亡率与消失率曲线分析
从图4可以看出,准噶尔山楂种群的死亡率(qx)与消失率(Kx)曲线变化趋势相似。波动中共出现4个高峰,其中Ⅰ龄级出现最大值,死亡率为0.899、消失率为2.296;在Ⅲ龄级出现最低谷,消失率为0.031,与种群期望寿命结果相对应;在其生活史中剩下3次环境筛的强烈筛选分别出现在Ⅵ龄级、Ⅸ龄级以及Ⅻ龄级,其所对应的死亡率依次为0.438、0.467、0.778,随着龄级的增加,种群表现为环境选择压力与高死亡率相对应的生长规律。
3.5 种群数量动态分析

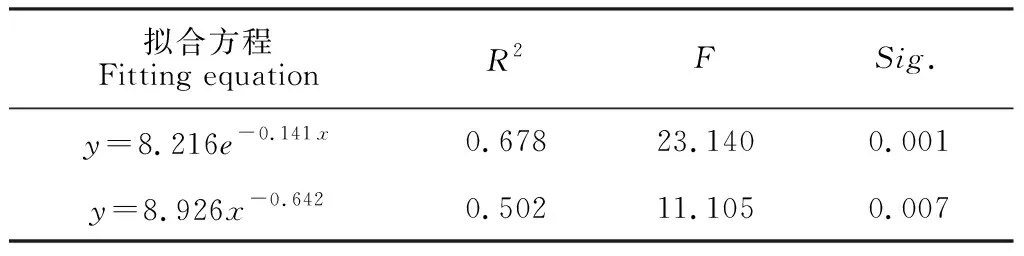
表2 新疆天山野果林准噶尔山楂种群存活曲线的检验
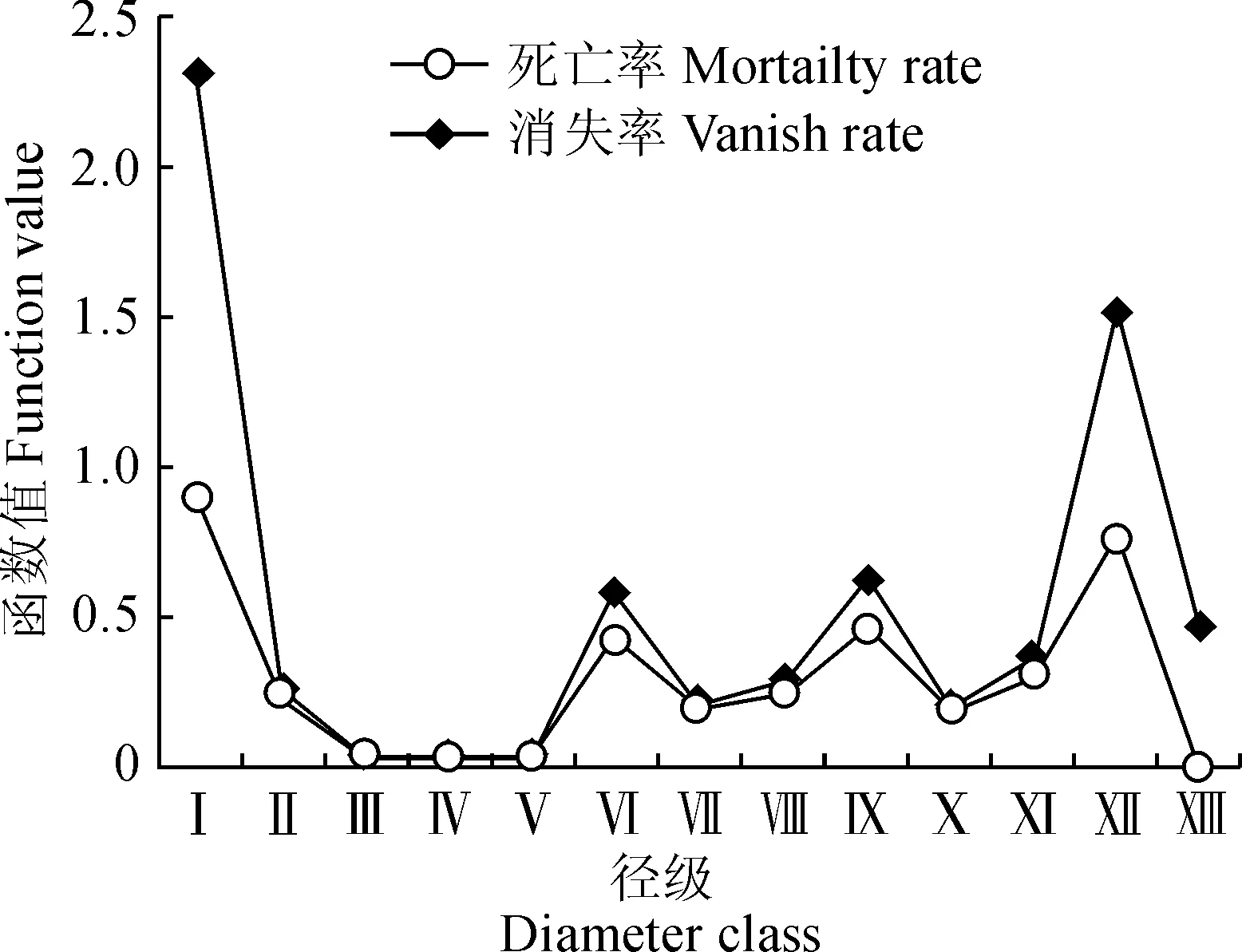
图4 新疆天山野果林准噶尔山楂种群的死亡率 与消失率曲线Fig.4 Curves of mortality and disappear rates of C. songarica populations in Tianshan Wild Fruit Forest of Xinjiang
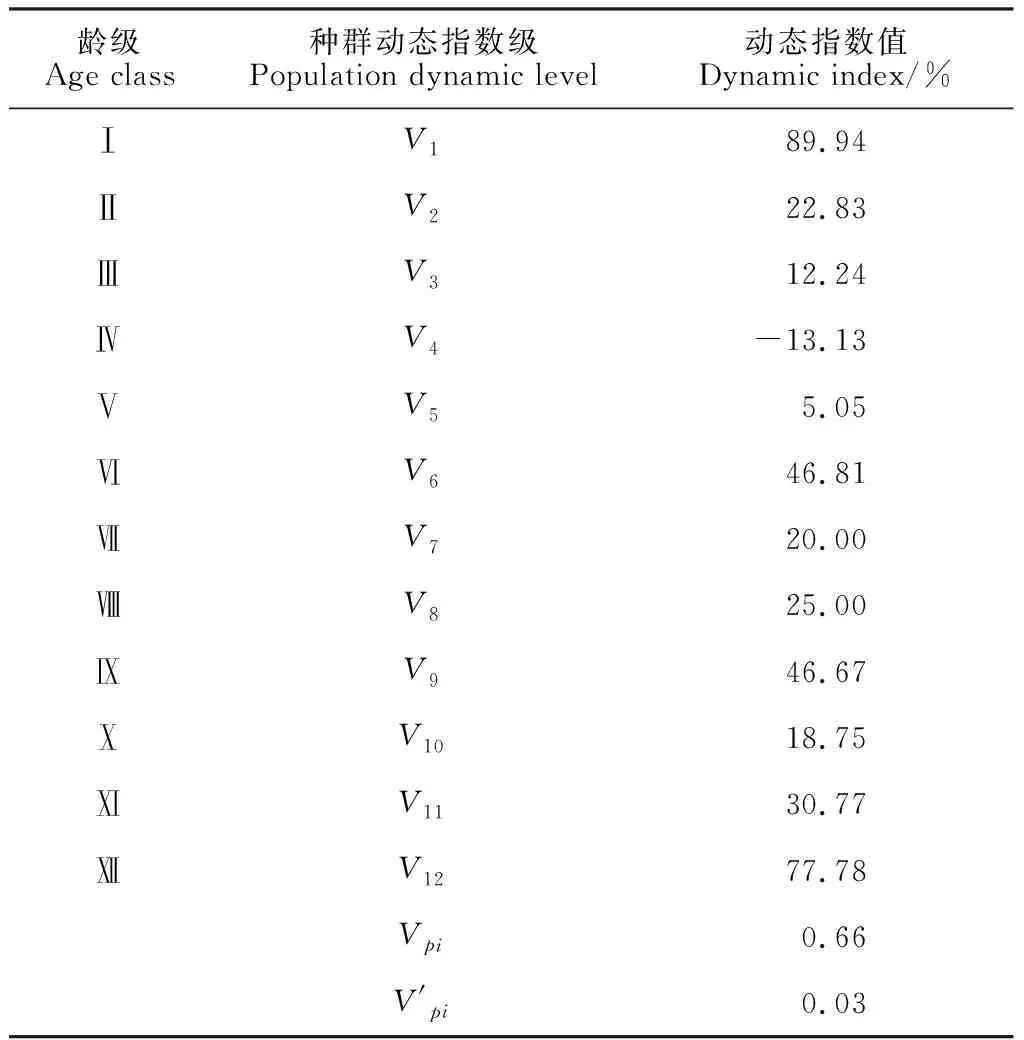
龄级Age class种群动态指数级Population dynamic level动态指数值Dynamic index/%ⅠV189.94ⅡV222.83ⅢV312.24ⅣV4-13.13ⅤV55.05ⅥV646.81ⅦV720.00ⅧV825.00ⅨV946.67ⅩV1018.75ⅪV1130.77ⅫV1277.78Vpi0.66V'pi0.03
3.6 生存分析
在准噶尔山楂种群的4个生存函数分析中分别以与径级相对应的龄级作为横坐标,以4个函数值为纵坐标作图(图5)。生存率函数曲线呈单调下降,相应的积累死亡率曲线前期小幅增加,后期趋于平稳,在特定龄级上生存率和死亡率两者互补,这说明准噶尔山楂种群在进入成熟期后,种群趋于稳定,但生存数量偏少;危险率函数曲线呈现单调递增的波动性变化,中后段龄级危险率随着龄级的增加而增大,与死亡率、消失率曲线变化趋势基本一致。因此,综合分析得出准噶尔山楂种群表现为前期减少、中期稳定、后期衰退的特点,与种群静态生命表、存活曲线、死亡率、消失率曲线结果相一致。
3.7 时间序列预测分析
采用一次移动平均法对准噶尔山楂的种群径级结构进行模拟和预测,其结果(表4)可以看出,在经历未来2、4、6、8、10龄级的时间之后,总体上准噶尔山楂种群各径级数量在预测序列中依次向后推移。2年后,准噶尔山楂种群幼树比例占62.3%,成体树占30.76%,老龄树占6.94%,4年后,成体树数量增长最多,占比例84.14%;6年后,成体树数量开始下降,老龄树比例占30.85%;8年后老龄树数量达到53.97%,与现在数量相比增长了50.34%;同时在10年后准噶尔山楂剩余量仅为353株,剩余率为18.33%。由此可见,尽管现有准噶尔山楂种群幼苗储量较丰富,但极高的死亡率和环境筛选强度,使种群的补充量不足,种群整体老龄个体数量呈现增长趋势。

图5 新疆天山野果林准噶尔山楂种群生存率函数Si、 累计死亡率函数Fi、死亡密度函数fi、危险率函数λiFig.5 Curves of survival rate (Si), accumulated mortality rate function (Fi), deadly density rate function (fi), risk rate function (λi) of C. songarica populations in Tianshan Wild Fruit Forest of Xinjiang
4 讨 论
在野生濒危植物种群的研究中年龄结构可以直观地反映出种群现存状态、数量动态特征,以及与环境之间的相互关系[29]。从年龄结构来看准噶尔山楂种群呈倒“J”型分布,属于增长型年龄结构,幼苗数量丰富,成体植株数量偏少,在这种情况下种群现阶段的稳定维持较为乐观,但由于准噶尔山楂种群幼苗在Ⅰ龄级中高达89.9%的死亡率,加之生物因素与非生物因素的双重影响,使种群在未来的发展中数量始终无法得到保障。然而,植物幼苗阶段作为种群更新过程中最重要和敏感的阶段,幼苗的数量变化能够反映种群的动态特征和未来发展趋势[30]。在调查中发现,准噶尔山楂种群的幼苗多聚集分布在母树树冠之下及其周围,虽然幼苗数量丰富,但发育转换生成的幼树偏少,随着幼苗的继续生长,个体间对养分、水分以及光照的争夺愈加强烈,而小环境中高大树木和草本植物的遮蔽,使光环境的异质性不断变化,幼苗在较高的郁闭度下无法充分接受光照。当不能满足幼苗生长所需的临界点时,负密度制约效应加强,种群死亡率开始升高。从Ⅱ龄级到Ⅴ龄级,准噶尔山楂种群个体的死亡率明显较低,这表明个体在该区间竞争压力小,自疏作用较弱。这也可能是由于准噶尔山楂种群幼树密度较低,环境容纳量能够兼容现有种群规模。Ⅴ龄级之后,准噶尔山楂种群生活史再次经历3个死亡高峰,分别在Ⅵ龄级、Ⅸ龄级以及Ⅻ龄级。这个可能是由于准噶尔山楂种群生长于特殊的山区地理位置与人为破坏,使得严重的洪水冲击、水土流失现象加剧了成体树、老龄树的毁坏程度。调查结果也发现,主干折断和整株倒伏是导致大部分准噶尔山楂种群成体死亡的直接原因,其中倒木现象比例更大。
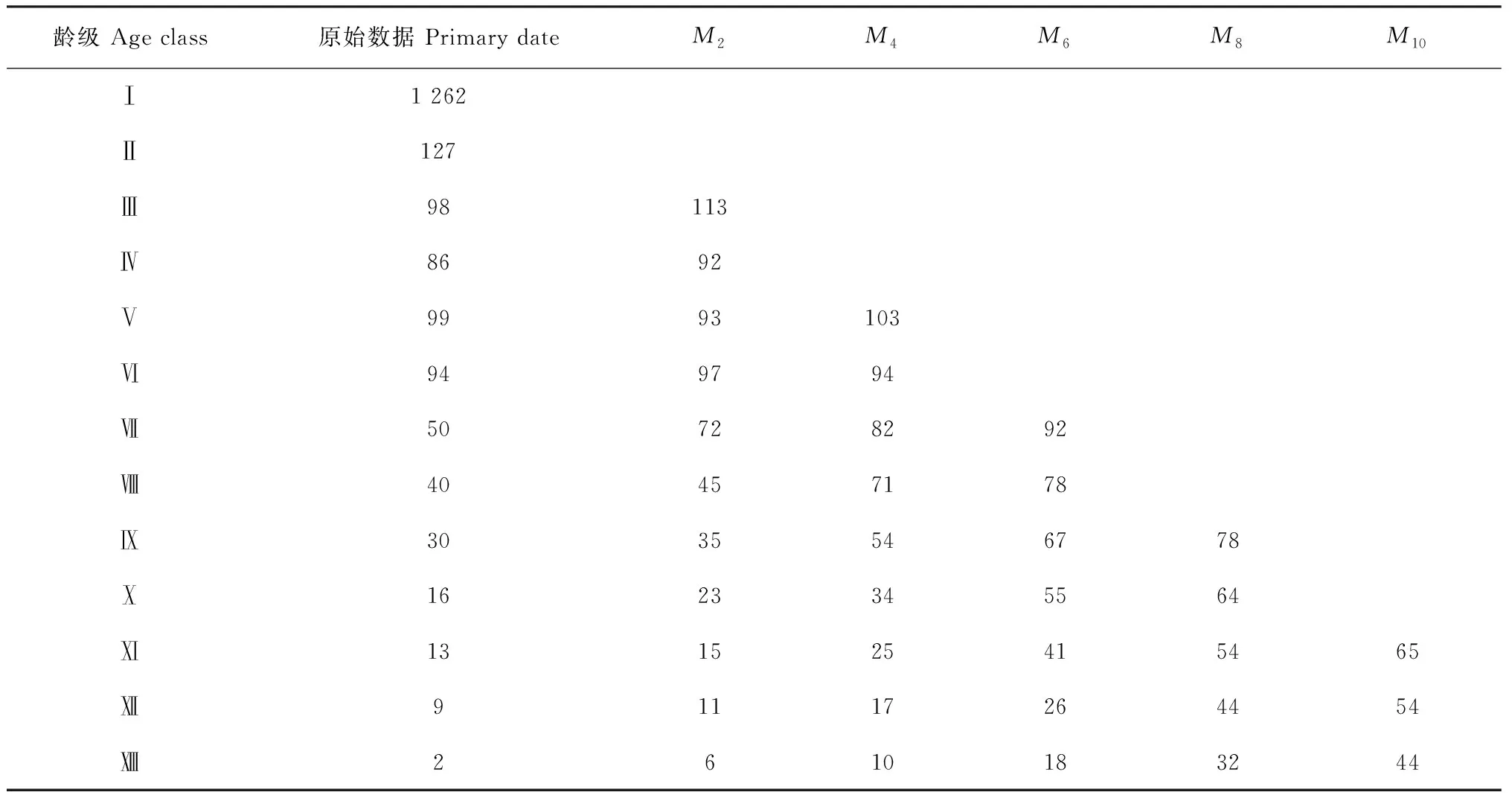
表4 新疆天山野果林准噶尔山楂种群数量动态时间序列预测
注:一次移动平均;M2、M4、M6、M8、M10分别表示对经历2、4、6、8和10年龄级时间后准噶尔山楂种群的数量预测
Note:Single moving average; M2, M4, M6, M8, M10represent the predicted number ofC.songaricapopulation through 2, 4, 6, 8, 10 size class time, respectively
存活曲线能够直观反映出生命表中种群个体在各年龄级的存活状况[31]。本研究中准噶尔山楂种群的存活曲线均趋于Deevey-Ⅱ型,在Ⅱ型中B1亚型种群的各时期成活量相差较大;B2亚型种群的各个年龄的死亡率基本相同,呈标准的对角线型;B3亚型种群幼年时期的死亡率较高,而成年后的死亡率则有所降低[32]。准噶尔山楂种群属于Deevey-Ⅱ型的B3亚型,这与同属于天山野果林重要组成树种的霍城县新疆野生樱桃李种群[19]、巩留县新疆野核桃种群[33]以及新源县新疆野苹果[16]种群存活曲线研究结果相似,这表明在伊犁地区天山野果林的野生果树资源趋同效应较为明显。而在田润炜等[34]的研究中,塔城地区天山野果林的新疆野苹果种群存活曲线为Deevey-Ⅰ型,各龄级的死亡率相近,种群幼苗个体较少,总体表现为衰退型种群。这与伊犁地区野生植物存活曲线存在差异,但在天山野果林大环境背景下,共同反映出不同地区不同树种之间均存在种源缺乏、人为干扰超负荷的更新危机。在实地调查中,准噶尔山楂种群林下牛、羊牲畜啃食幼苗的现象极为严重,在这种没有及时和足够的幼龄个体补充的前提下,准噶尔山楂种群趋于衰退的风险升高。另外,从生存分析来看,准噶尔山楂种群总体危险率函数曲线呈现一定波动性,波动过程中前期锐减,中期稳定,后期衰退,与刘忠权等[35]对新疆野苹果研究中前期衰退、中期相对稳定、后期不波动的生存分析特点相似,加之准噶尔山楂种群总体外部干扰指数(0.03%)接近于零,极易受外界环境影响,在未来发展中种群仍然表现出一定的不稳定性[36]。因此,结合准噶尔山楂种群自身的生物条件和环境条件,对人为干扰较大的准噶尔山楂种群分布区,进行实施禁牧及围封育林措施,为野生濒危植物准噶尔山楂种群的生长发育创造条件,促进种群良好的更新和维持。
时间序列预测分析具有较高的准确性,在准噶尔山楂种群时间序列分析中种群各径级株数在预测序列中依次向后推移,老龄个体逐渐增多,与甘肃省裸果木(Gymnocarposprzewalskii)种群[37]、海南省野生荔枝(Litchichinensis)种群[38]的时间序列预测结果相似。种群极易受到人为干扰的影响,种群幼苗存活率极低的“瓶颈”现象,不仅与准噶尔山楂种群自身的更新机制有关,更重要的是生存环境遭受破坏的影响。近年来当地大力开发旅游业,每年9月挂果期间福寿山景区日游客量可达2 000人次,景区开设的野果采摘等旅游特色对准噶尔山楂果实造成严重破坏性,这种生境破坏和人为砍伐造成准噶尔山楂种群结构发生变化,种群的生存受到极大的威胁,导致准噶尔山楂种群更新能力不足,受损群体难以复壮和扩展。
在研究区中,自然地质灾害、过度放牧啃食以及不合理的生态旅游开发等都是准噶尔山楂种群发展不稳定的重要影响因素。天山野果林中准噶尔山楂具有较强的抗寒、抗旱性以及耐贫瘠特性,是山楂育种的重要种质资源,且根据调查显示,现阶段准噶尔山楂种群仅分布于大、小西沟、果子沟以及吉尔格朗沟等山区,与前人研究中巩留县、新源县均有分布的研究结果不相符[39],极小的分布范围与极少的种群数量,急需对准噶尔山楂种群进行有效的保护和利用。因此,针对准噶尔山楂种群生存现状,提出以下2点保护建议:(1)对于准噶尔山楂幼苗、幼树采用适当人工抚育,促进其种群的天然更新,对缓坡准噶尔山楂植株,进行林下杂灌清除,对陡坡准噶尔山楂采用点状采伐,确保林下光线和营养有效分配,有利种群天然更新;(2)加强当地生境的保护,减少人为采摘、严禁放牧,逐步提升准噶尔山楂种群的自然恢复能力,为天山野果林野生植被资源的长期稳定发展提供有力保障。
