基于瞳孔光响应的光谱灵敏度模型研究初探
2018-01-06沈志豪韩秋漪张善端
沈志豪,徐 蔚,韩秋漪,张善端
(复旦大学电光源研究所,上海 200433)
基于瞳孔光响应的光谱灵敏度模型研究初探
沈志豪,徐 蔚,韩秋漪,张善端
(复旦大学电光源研究所,上海 200433)
本文综述了现有的非视觉生物效应光谱灵敏度研究,设计了一套新的实验装置和流程研究瞳孔光响应。根据稳定瞳孔收缩率和最大瞳孔收缩率数据,基于数据拟合和归一化处理,获得了两个基于瞳孔光响应的光谱灵敏度曲线,其峰值波长分别为482.8 nm和495.6 nm,对应的曲线半峰值宽度为95.1 nm和62.9 nm。
非视觉生物效应;生理表征;瞳孔光响应;光谱灵敏度
引言
众所周知,光对人的影响包括两部分。一方面,人眼的视觉细胞在光信号的刺激下形成神经冲动,传至大脑皮层产生视觉;另一方面,人体的生理参数和心理状况会受到光的影响,这些影响通常被称为光的非视觉生物效应,主要包括生理节律、激素的分泌与抑制、瞳孔光响应、警觉性影响等[1]。
目前的照明标准是基于满足人体的视觉需求而建立的,主要考虑的光参数为亮度、照度、色温、显色指数、均匀度和眩光等。然而光的非视觉生物效应对人的健康也会产生很大的影响,例如在夜晚,光会抑制人体褪黑激素的分泌,而研究表明褪黑激素在正常的生理浓度下可以抑制Ⅱ型抗炎细胞因子(具有促进肿瘤生长的活性)的产生[2]。此外,光的非视觉生物效应也能用于造福人类,例如通过特定光照强度和光谱能量分布的人工光以及自然光来修复人的昼夜节律紊乱的状况,帮助恢复人体的健康[3];利用动态光策略,在工厂环境下实现工作与休息的良好协调,保证工人的工作效率和健康[4]。因此,未来的照明设计和实践将综合考虑视觉需求和非视觉生物效应的影响。为了更好地制定照明新标准,我们需要基于光参数来量化非视觉生物效应的影响,其中包括非视觉生物效应与光谱之间的关系,即非视觉生物效应光谱灵敏度模型的研究。
1 现有的非视觉生物效应的光谱灵敏度研究
目前人们对于非视觉生物效应的光谱灵敏度开展了一些研究,实验中的特征参量包含一个或多个生理参数。这些生理参数包括光致褪黑激素的抑制、瞳孔光响应、生理节律、警觉反应、心跳、体温等,其中被采用最多的生理表征是前两个。
早在2001年,Brainard等人[5]就通过实验得到了光致褪黑素抑制的作用谱。实验选取受试者在凌晨接受全视野的单色光刺激,然后抽取受试者血液测量,比较光刺激前后血液中的褪黑激素的含量,最后通过数据处理得到了光致褪黑素抑制与单色光波长之间的关系。实验表明光致褪黑素抑制的峰值波长为464 nm。Thapan等人[6]也在同一年得到了光致褪黑素抑制的作用谱,其峰值波长为459 nm(457~462 nm,相关系数r2=0.73)。这两个研究组对于光致褪黑素抑制的光谱灵敏度的研究,为后来视网膜上第三种感光细胞的发现奠定了基础。
2002年,美国《科学》杂志同期发表Hattar等人[7]和Berson等人[8]的2篇研究论文,揭示哺乳动物视网膜上除了视锥细胞和视杆细胞外,还有本征感光视网膜神经节细胞(ipRGC),即第三种感光细胞,证实了褪黑激素调控人体生理节律等非视觉生物效应的生理基础。此后非视觉生物效应成为研究热点。
除了以光致褪黑素抑制作为生理表征,科研人员也常常以瞳孔的光响应作为研究对象。瞳孔的光响应,是指瞳孔直径随着人眼处照度的变化而产生变化的生理效应。2007年,Gamlin等人[9]先后对猕猴和人类在不同波长单色光下的瞳孔光响应进行了实验研究,获得不同波长下达到相同瞳孔收缩率的人眼处辐照度,从而得到了光谱灵敏度曲线。2012年,McCormick等人[10]以Lolliguncula brevis(一种乌贼)为实验对象,研究其在不同波长下的瞳孔收缩情况,从而探讨非视觉生物效应的光谱敏感性。
对比以上两种以不同生理参数为表征的实验研究,我们发现只有在光照达到一定强度时才会出现光致褪黑素抑制的现象。同时,给光刺激与褪黑素含量下降之间具有一定的时间差,适用性和实时性有待商榷。此外,若要增加实验数据量来优化这条光谱曲线模型,则通过采集人体血液来测量褪黑素含量的方法会使整个实验变得无比繁琐和冗长。相对而言,以瞳孔光响应作为生理表征来进行实验,实时性更好,实验结果更直观,实验流程相对简单,耗时短,适用于利用大量实验来提高光谱灵敏度曲线的准确度。
回顾现有的非视觉生物效应的光谱灵敏度研究,出于研究目的的不同,这些实验所得的作用谱均有一定的应用限制,且研究多采用6~8个波长的单色光来进行实验,最终实验拟合曲线的准确性有待提高。本研究通过建立一套新的瞳孔实验装置和实验流程,实现高可行性和高重复性的瞳孔光响应实验;同时采用更多波长的单色光进行实验,以提高实验数据量,从而拟合出更加准确的基于瞳孔光响应的光谱灵敏度模型。
2 瞳孔光响应实验
本实验将对12种不同波长的窄带单色光刺激下的瞳孔光响应进行研究,记录瞳孔光响应的动态变化过程。
2.1 实验装置
实验装置如图1所示,根据实现的功能分为两个部分:单色光发生部分和瞳孔直径测量部分。
单色光发生部分主要由灯箱、氙灯电源、透镜、光栅单色仪和出射光管组成。灯箱包括短弧氙灯、散热风扇和排风管。所用短弧氙灯(深圳凯世光电,KDX-2000SD)的功率2 000 W,工作电压24.3 V,额定电流75 A。氙灯位于定制的灯箱中部,所产生的热量由散热风扇和排风管及时排出,以保证灯箱内的温度适当,氙灯正常工作而不至过热。灯箱正对单色仪的侧面开孔,孔对氙灯中心所张立体角与透镜对氙灯中心所张的立体角相同,且该立体角与单色仪的数值孔径匹配。氙灯电弧发出的光从灯箱上的孔射出后,经透镜聚焦,氙灯电弧1∶1成像在光栅单色仪(上海光学仪器厂,44 W,Czerny-Turner型结构)的入射缝上;光进入单色仪内部经光栅分光,所需要的特定波长的单色光再从出射缝射出到出射光管。出射光管内部涂黑,另一端放置漫射板,获得相对均匀的面光源。
瞳孔测量部分由暗室、颚托、额托和眼动仪组成。为了保证实验受试者仅接受特定波长的单色光的刺激,我们用木板搭建了一个暗室。实验时暗室内唯一的光源就是来自于出射光管一端的漫射板的单色光。颚托和额托用于固定实验受试者头部的位置,保证在实验时不发生变动。此外,本研究采用Tobbi Glasses眼镜式眼动仪来记录瞳孔光响应的动态变化过程,易携带和佩戴,采样频率为30 Hz,瞳孔数据测量结果精确。在每次实验测量前,通过定标来确定受试者瞳孔的初始大小,从而缩小实验的误差。每次实验记录的数据会单独保存为一个文件,在实验结束后通过眼动仪配套软件批量导出至计算机,进行下一步的数据处理。
2.2 受试者
共有4位受试者参与了基于瞳孔光响应的实验初步研究。受试者1男3女,年龄23~25周岁,每个人都完成了450~620 nm区间内12个不同波长、5种不同强度的单色光下的实验,获得共60组瞳孔收缩数据。所有的受试者具有正常的矫正视力和正常的色觉,身体健康,无眼科疾病和精神类疾病,实验周期内未服用任何药物且具有正常和自然的作息习惯。
2.3 实验方法和流程
受试者在实验当天于规定的时间到达实验室,先对实验室环境进行适应,并在暗室内根据受试者的个体情况调节座椅、颚托和额托的位置,使得受试者的左眼完全处于出射光管一端的漫射板的单色光视野内。完成相关准备工作后,受试者用托吡卡胺眼药水对自己的左眼进行散瞳,每隔5 min滴一次,共滴5次,以达到完全散瞳的效果(专业眼科医生的建议)。完成散瞳后,实验受试者戴上自制的遮光镜进行暗适应,遮光镜由黑色泳镜外加黑色胶带缠绕而成,保证在佩戴时间内受试者处于完全黑暗中。
在进行10 min的暗适应后,第一位受试者在实验工作人员的带领下前往暗室,在工作人员指导下摘下遮光镜佩戴眼动仪,并进行定标。眼动仪记录给光前20 s,光刺激20 s和撤光后30 s共70 s的受试者右眼瞳孔动态变化数据。完成一次测量后,受试者立即戴上遮光镜进行下一轮实验的暗适应;而第二位受试者开始进行本轮测试。采用这种交替实验的方法,可以缩短实验周期。在两轮实验的间隔期,实验工作人员通过更换减光片来调节所给光的强度,通过调节光栅单色仪来变换出射光管的单色光波长。
在每轮实验前,确定受试者眼睛的位置后,用光谱照度计(杭州科兴光电有限公司,CL-200F)来测量人眼处的辐照度大小,并将数据导入计算机。实验中,共测量了450~620 nm区间内12个不同波长单色光刺激下的瞳孔变化情况。对于每个波长的单色光,辐照度从小到大依次测量,完成5个不同强度的实验。
3 波长和辐照度对瞳孔收缩率的影响
眼动仪内的瞳孔数据通过专业软件导入计算机,通过数据处理,得到一些初步的结果。
通过分析某一波长一定强度的单色光刺激下瞳孔直径的动态变化曲线,我们发现瞳孔光响应分为几个阶段:在给光后,瞳孔会快速收缩至最小值;然后在持续给光阶段,瞳孔会从最小值逐渐略微增大至稳定值;最后在撤光后,瞳孔又缓慢恢复。
本研究根据实验数据计算瞳孔收缩率(pupil contraction rate,PCR),计算公式如下:
其中,D1为给光前瞳孔直径;D2为给光后瞳孔收缩后的直径。数据处理时,给光前瞳孔直径取给光前倒数第99至倒数第50共50个采样点的瞳孔平均直径;而给光后收缩瞳孔的直径取瞳孔最小值前后共5个采样点的瞳孔平均值。不同波长不同强度的单色光刺激下,瞳孔收缩率最大值数据如表1所示。
根据表1,我们发现在同一波长的单色光照射下,瞳孔收缩率随辐照度增加而逐渐增大;辐照度愈大,瞳孔收缩现象愈加明显,但瞳孔收缩率趋向饱和。
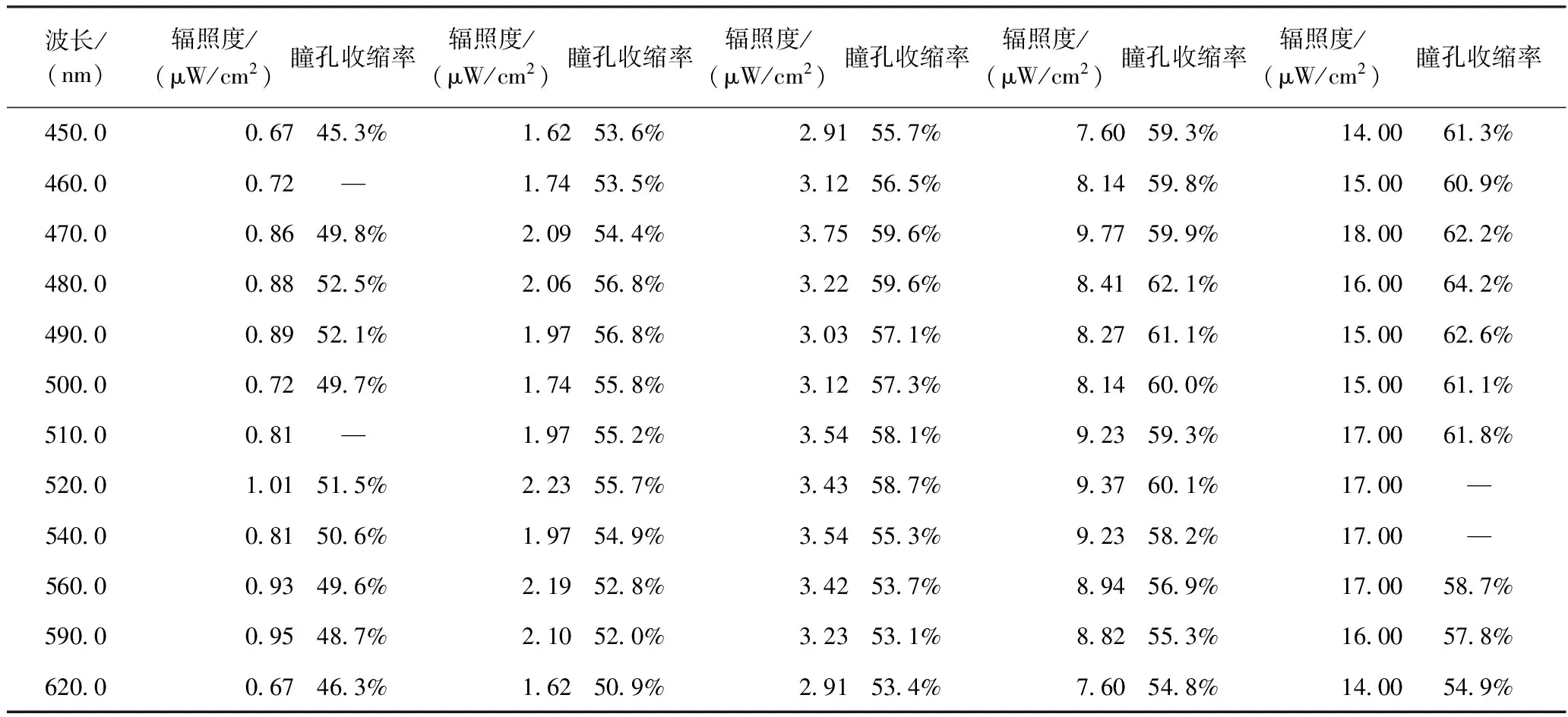
表1 不同波长不同强度的单色光刺激下的瞳孔最大收缩率Table 1 Pupil constriction rate under different wavelength and irradiance
此外,在辐照度相近的条件下,不同波长的瞳孔收缩情况也有比较明显的差异。如表1中所示,在辐照度同为16 μW/cm2,480 nm单色光照射下瞳孔收缩率比590 nm单色光照射下大6.4%,说明波长是影响人眼瞳孔收缩率的一个非常重要的因素,也即人眼的瞳孔收缩具有明显的光谱响应差异。
4 光谱响应模型的推导
基于实验所得不同波长不同强度单色光下的瞳孔收缩率数据,我们通过数学推导和归一化处理,得到合理的光谱响应模型。
光度学中,光通量Φ是指能够被人的视觉系统所感受到的那部分光辐射功率的大小的度量,用来表征光量的多少,计算公式如下:
其中,Km代表最大光谱光视效能,明视觉条件下Km=683 lm/W;P(λ)代表绝对光谱能量分布;V(λ)代表明视觉光谱光视效率函数。参考式(2),我们可以类比得到非视觉生物效应的计算公式:
其中,C(λ)即为需要推导的基于瞳孔收缩的非视觉生物效应的光谱灵敏度响应曲线。
根据建立明视觉光谱光视效率函数的基础,以及实验心理学中物理量与感受量之间的对数关系,相应的由非视觉生物效应引起的瞳孔收缩率PCR可以表示为
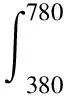
由此可得,在不同波长的单色光下,取相同的瞳孔收缩率,对该瞳孔收缩率下的辐照度作归一化处理,即可得到光谱灵敏度响应曲线C(λ)。
根据实验所得数据,在同一波长下,随着人眼处辐照度的增加,人眼的瞳孔稳定收缩率也会增大。在低辐照度条件下,瞳孔收缩率的变化比较剧烈;而随着辐照度的增加,瞳孔收缩率逐渐趋于缓和。我们用Hill方程对同一波长下瞳孔收缩率PCR与辐照度E之间的关系进行拟合,公式如下:
根据所得的各波长不同辐照度下的瞳孔最大收缩率,用计算机软件拟合,得到不同波长下的瞳孔收缩率与人眼处辐照度的函数关系以及曲线。各波长下拟合参数如表2所示。

表2 不同波长下瞳孔收缩率与人眼处辐照度的函数参数Table 2 The parameters of Pupil constriction rateunder different monochromatic light
各波长的瞳孔收缩率与人眼处的辐照度之间的拟合曲线如图2所示。
根据实验数据,设定55%为瞳孔收缩率的响应灵敏度归一化指标,根据式(5)和表2拟合得到的函数,可以求得各个波长单色光照射条件下在55%瞳孔收缩率时人眼处所需辐照度值。根据公式(3),可得当PCR相同时,
由实验数据,可得480 nm波长下达到55%瞳孔收缩率的所需辐照度最小,因此令此时的C(λ)=1,则可以得到归一化条件下各波长下的C(λ)值,如表3所示。

图2 各波长的瞳孔收缩率与人眼处的辐照度之间的拟合曲线Fig.2 Pupil constriction rate PCR as a function of irradiance E under different monochromatic light
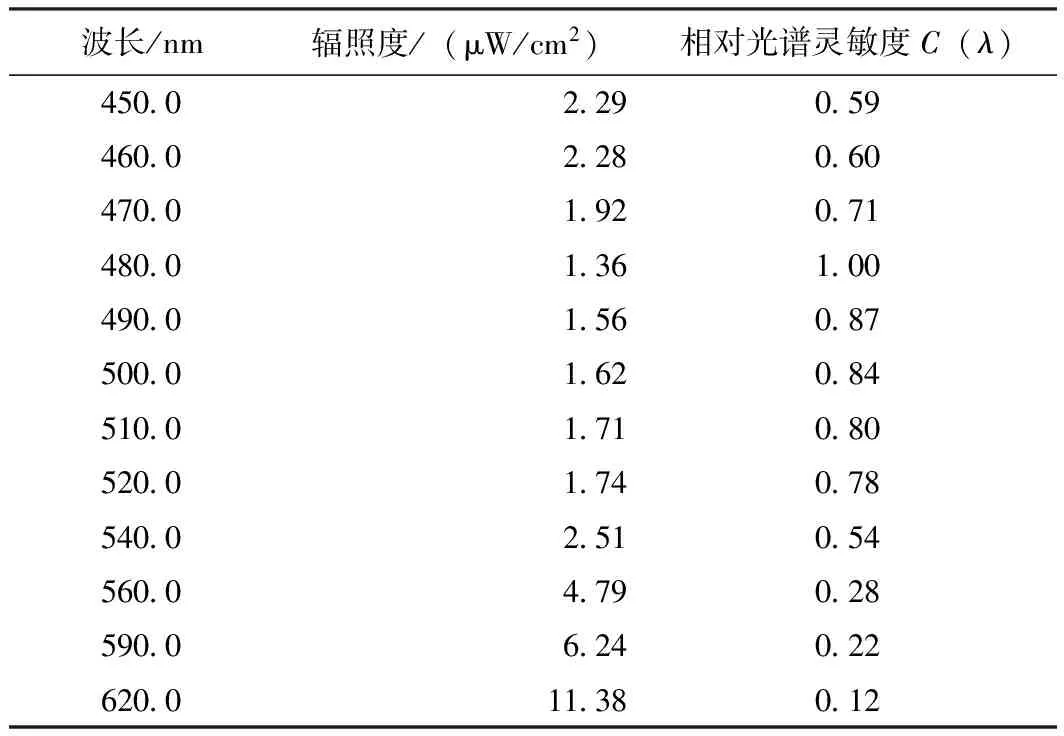
表3 不同波长下达到55%瞳孔收缩率时对应的相对光谱灵敏度Table 3 Relative spectral sensitivity C(λ) at PCR=55% for different monochromatic light
分析各类光谱响应灵敏度曲线,发现其分布规律非常符合高斯函数。因此C(λ)函数模型采用高斯分布公式[11-12]:
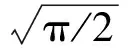
将上述不同波长及其对应的相对光谱灵敏度值代入式(7),求得峰值波长λmax和待定系数D,得到瞳孔收缩率的光谱响应分布函数式如下:
即峰值波长为495.6 nm,曲线半高宽根据公式(8)计算可得为95.1 nm,对应的光谱响应灵敏度曲线的拟合相关系数r2=0.84,如图3(a)所示。
式(10)为以最大瞳孔收缩率值为数据基础得到的拟合曲线,若根据给光期间瞳孔收缩率的稳定值为数据基础,可得到另一条拟合曲线,光谱响应分布函数式如下:
其峰值波长为482.8 nm,曲线半高宽根据公式(8)计算可得为62.9 nm,对应的光谱响应灵敏度曲线的拟合相关系数r2=0.75,如图3(b)所示。
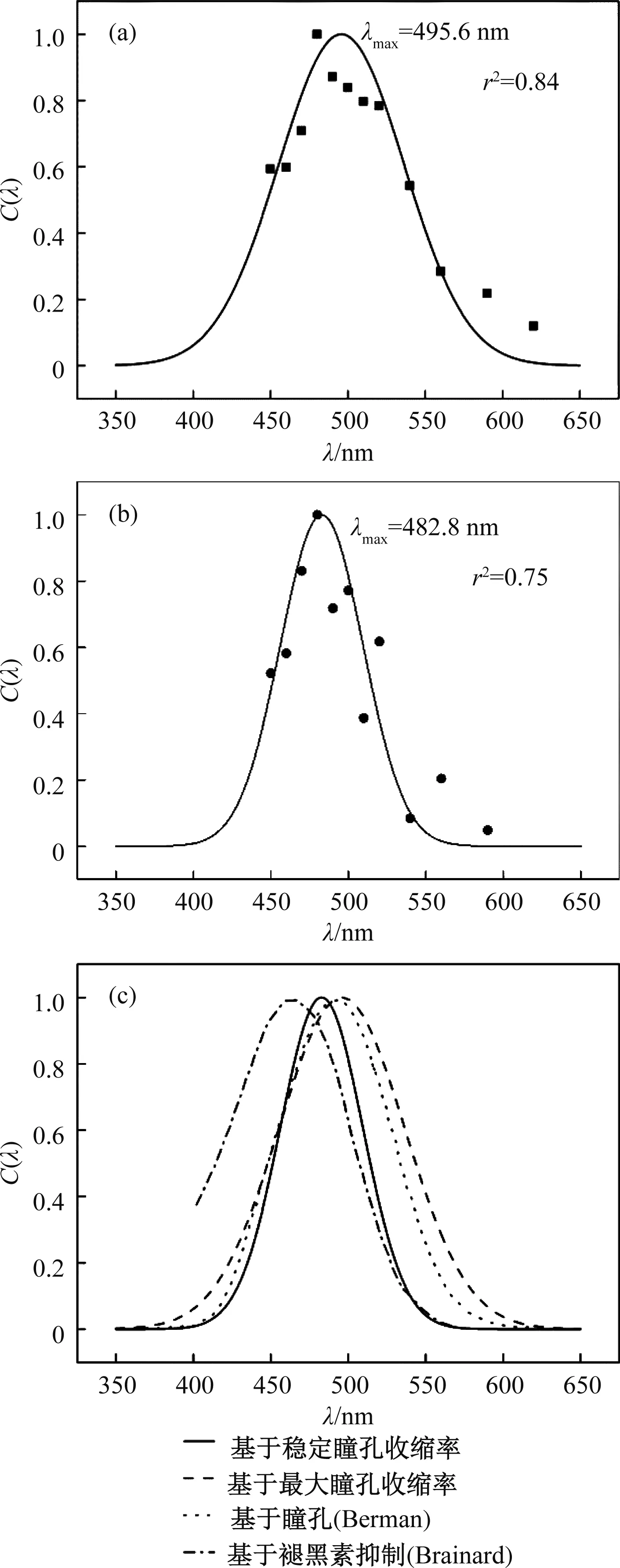
图3 基于瞳孔光响应的光谱灵敏度曲线图Fig.3 Spectral sensitivity curve based on pupillary light reflex
如图3(c)所示,将本文分别基于稳定瞳孔收缩率和最大瞳孔收缩率数据所得的光谱灵敏度曲线与Berman于2008年提出的基于瞳孔光响应的光谱灵敏度曲线[13]、Brainard于2001年实验所得的基于光致褪黑素抑制的光谱灵敏度曲线[5]进行了对比。一方面,本文基于最大瞳孔收缩率数据所得的光谱灵敏度曲线(λm=495.6 nm)与Berman所得的曲线(λm=491 nm)非常接近,而基于光致褪黑素抑制的光谱灵敏度曲线的峰值波长则为464 nm。
根据现有的研究结果[14-16],我们知道视锥细胞、视杆细胞和本征感光视网膜神经节细胞共同参与了瞳孔光响应的生理过程。本文所得的光谱灵敏度曲线与光致褪黑素抑制曲线相比,表现出波长红移现象,也恰恰证明了这一结论。另一方面,基于稳定瞳孔收缩率拟合所得曲线比基于最大瞳孔收缩率拟合所得曲线要更偏向短波方向,更靠近光致褪黑素抑制的光谱灵敏度曲线,其可能的原因是在光刺激导致瞳孔收缩的初期,视网膜神经节细胞所起的作用不明显。而在瞳孔的稳定收缩期,视网膜神经节细胞更多地参与进来,且由于其光谱响应对短波更为敏感,从而导致了最终的曲线向短波偏移。
5 结论
在照明设计和照明实践中,非视觉生物效应的影响越来越无法忽视,如何量化照明光参数和非视觉生物效应生理参数之间的关系是一个重要的研究方向。基于此目的,我们建立了一套新的瞳孔光响应实验装置和实验流程,来实现更准确、重复更好的瞳孔光响应实验。我们对人眼的瞳孔光响应这一生理参数进行了初步研究,测量了瞳孔收缩率与眼睛处辐照度和波长的关系。根据稳定瞳孔收缩率和最大瞳孔收缩率数据,基于公式推导和数据归一化处理,建立了基于瞳孔光响应的光谱灵敏度模型,其峰值波长为482.8 nm和495.6 nm,与传统的视觉感光细胞的光谱灵敏度曲线有较大差异。
未来,在本文研究的基础上增加受试者数量,进一步优化非视觉生物效应的光谱灵敏度模型,将其与照明设计方法和照明标准结合起来,更好地满足不同场所的照明设计。
[1] 罗明,郑诗琪,叶 鸣.动态光对人警觉度与表现的影响[J].照明工程学报,2016,27(6):1-5.
[2] MOCELLIN S,MARINCOLA F M,YOUNG H A.Interleukin-10 and the immune response against cancer:a counterpoint[J].Journal of Leukocyte Biology,2005,78(5):1043-1051.
[3] 王茜,郝洛西,曾堃.健康光照环境的研究现状及应用展望[J].照明工程学报,2012,23(3):12-17.
[4] 何思琪.工厂环境下的工作-休息排程与动态光策略研究初探[J].照明工程学报,2016,27(6):53-60.
[5] BRAINARD G C,HANIFIN J P,GREESON J M,et al.Action spectrum for melatonin regulation in humans:Evidence for a novel circadian photoreceptor[J].The Journal of Neuroscience,2001,21(16):6405-6412.
[6] THAPAN K,ARENDT J,SKENE D J.An action spectrum for melatonin suppression:evidence for a novel non-rod,non-cone photoreceptor system in humans[J].Journal of Physiology,2001,535(1):261-267.
[7] HATTAR S,LIAO H W,TAKAO M,et al.Melanopsin-containing retinal ganglion cells:architecture,projections,and intrinsic photosensitivity[J].Science,2002,295:1065-1070.
[8] BERSON D M,DUNN F A,TAKAO M.Phototransduction by retinal ganglion cells that set the circadian clock[J].Science,2002,295:1070-1073.
[9] GAMLIN P D,MCDOUGAL D H,POKORNY J,etal.Human and macaque pupil responses driven by melanopsin-containing retinal ganglion cells[J].Vision Research,2007,47(7):946-954.
[10] MCCORMICK L R,COHEN J H.Pupil light reflex in the Atlantic brief squid,Lolliguncula brevis[J].Journal of Experimental Biology,2012,215(Pt 15):2677-2683.
[11] XU W,VAN BOMMEL W.Inferior vs superior:Inferior retinal light exposure is more effective in pupil contraction in humans[J].Light & Engineering,2011,19(2):14-17.
[12] 徐蔚.基于瞳孔收缩的非视觉感光系统的研究[D].上海:复旦大学,2011.
[13] BERMAN S M.A new retinal photoreceptor should affect lighting practice[J].Lighting Research and Technology,2008,40(4):373-376.
[14] TSUJIMURA S I,UKAI K,OHAMA D,et al.Contribution of human melanopsin retinal ganglion cells to steady-state pupil responses[J].Proceedings of the Royal Society B,2010,277:2485-2492.
[15] GOOLEY J J,MIEN I H,HILAIRE M A S,et al.Melanopsin and rod-cone photoreceptors play different roles in mediating pupillary light responses during exposure to continuous light in humans[J].The Journal of Neuroscience,2012,32(41):14242-14253.
[16] MCDOUGAL D H,GAMLIN P D.The influence of intrinsically photosensitive retinal ganglion cells on the spectral sensitivity and response dynamics of the human pupillary light reflex[J].Vision Research,2010,50:72-87.
ElementaryResearchonSpectralSensitivityModelBasedonPupillaryLightReflex
SHEN Zhihao,XU Wei,HAN Qinyi,ZHANG Shanduan
(DepartmentofLightSourceandIlluminatingEngineering,FudanUniversity,Shanghai200433,China)
This paper reviewed the existed research on spectral sensitivity of non-visual biological effects,and designed a set of new experimental setup and testing procedure.According to the experimental results of stable pupil contraction rate and maximum pupil contraction rate,two spectral sensitivity models are established based on pupillary light reflex,the peak wavelengths of which are 482.8 nm and 495.6 nm,and their corresponding full width at half maximum are 95.1 nm and 62.9 nm.
non-visual biological effects;physiological symptoms;pupillary light reflex;spectral sensitivity
国家自然科学基金项目(61308080),上海杨浦区国家创新型试点城区建设与管理资金项目(2014YPCX01-003)
TM923
A
10.3969/j.issn.1004-440X.2017.06.024
