Formin’ bridges between microtubules and actin filaments in axonal growth cones
2018-01-05IstvánF?ldi,SzilárdSzikora,JózsefMihály
PERSPECTIVE
Formin’ bridges between microtubules and actin filaments in axonal growth cones
It is well established that guidance of axons during neuronal development is regulated by a variety of extracellular signals,governing cytoskeletal dynamics in growth cones. The actin and microtubule (MT) cytoskeleton have both been shown to play important roles. However, a growing body of work suggests that a critical issue is the proper coordination of changes within these two major cytoskeletal systems (reviewed in Cammarata et al., 2016). Previous studies led to the identification of a number of proteins involved in actin-microtubule crosstalk in neurons, in particular, several MT plus-end tracking proteins or+TIPs were shown to be at work. In addition to the well-known+TIP proteins, members of the formin protein family were also identified as regulators of actin and MT dynamics, initially in non-neuronal cells (reviewed in Bartolini and Gundersen,2010) and more recently in neurons (Szikora et al., 2017). Most formins, belonging to the diaphanous related formin (DRF)subfamilies, share a conserved domain structure. They consist of the formin homology domains FH1 (required for pro filin-actin interaction) and FH2 (required for actin assembly and protein dimerization), and a set of regulatory domains (GBD, DID,DD, DAD) (Figure 1A), which provide the means to control the spatial and temporal activity of the FH2 domain. Although formins were initially described as F-actin barbed end binding proteins, exhibiting an actin nucleation and elongation activity,subsequent work revealed that many formins have the potential to affect MT organization and/or dynamics in cellular systems,including neurons. Despite these advances, until recently, the contribution of formins to the government of neuronal actin-microtubule crosstalk remained largely elusive.
Capture and coalignment of growth cone MTs and filopodial actin bundles:Growth cone actin filaments and MTs are in a constant state of turnover, especially in the growth cone periphery. Of the various changes, MT capture followed by coalignment with filopodial actin bundles is thought to be a key feature of the growth cone cytoskeleton. According to the current view,co-aligned growing MTs transiently couple to the constant actin retrograde flow, removing them from the growth cone periphery during steady states. Whilst in the presence of attractive guidance cues, the on-axis pioneer MTs stabilize along the stalled filopodial actin bundles prior to growth cone turning/advance (Figure 1B-B”). A recent study (Preciado López et al.,2014) revealed that growing MTs with the appropriate plus end protein complex can selectively capture actin bundles while they cross-over individual actin filaments. They used an artificial protein (TipAct) which contains a series of actin-binding calponin homology domains and an SxIP motif to bind EB proteins.In the presence of EB proteins, TipAct is localized to the growing microtubule ends where it possesses a weak actin binding affinity, sufficient to con fine its localization to actin rich regions,e.g.,actin bundles. It was shown that actin bundles can capture and redirect growing MTs and they can even force MTs to grow along them in tight associationin vitro. They demonstrated that the MT tip complex plays an essential role in the initial interaction; however, additional mutual lattice binders are probably necessary for a more stable co-alignment between actin filaments and MTs. By employing a sensibly designed synthetic protein, these experiments helped to gain insight into the general mechanism of actin-MT co-alignment. However, among other proteins, several formins were already identi fied with the ability to bind actin and MTs simultaneously. This raised the possibility that formins might mediate actin-MT interactions,including MT capture and co-alignment. Accordingly, INF2 has been shown to co-aggregate MTs and actin filaments in vitro (Gaillard et al., 2011), and furthermore mDia formins were found to be essential in cortical microtubule capture in breast carcinoma cells (Daou et al., 2014). In line with these findings,we recently demonstrated a role for Drosophila DAAM, a DRF type of formin, during the coordinated regulation of actin and MT cytoskeletons in primary neurons (Szikora et al., 2017).TIRFM and co-sedimentation experiments demonstrated that DAAM has the capacity to simultaneously bind and co-align the two cytoskeletal filaments in vitro. Moreover, we found that DAAM is localized along overlapping MTs and actin filaments in primary neurons. Also, the loss of DAAM effectively reduces the retrograde MT translocation frequency, but only weakly affects the retrograde actin flow. Together, these data strongly suggest that DAAM is involved in the coordination/co-alignment of the two cytoskeletal systems. Whether DAAM is pivotal in all neurons or if formins act redundantly in this system, remains to be tested.
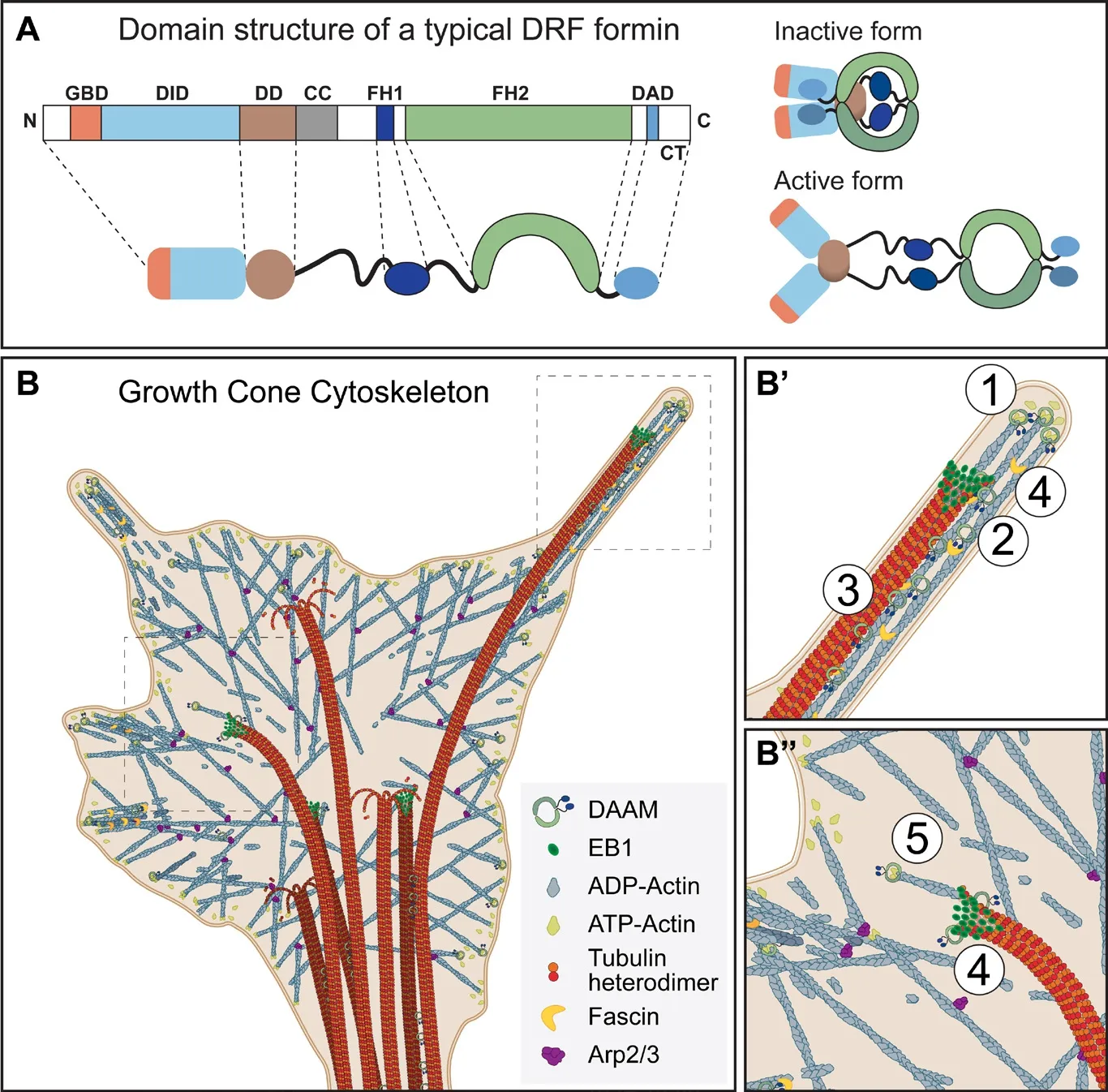
Figure 1 Formin mediated actinmicrotubule interactions in the growth cone(A) Domain structure and regulation of a typical Diaphanous-related formin (DRF).The domains abbreviated are as follows:GTPase binding domain (GBD), diaphanous inhibitory domain (DID), dimerization domain (DD), coiled-coil region (CC),formin homology 1 (FH1) and 2 (FH2)domains, followed by the diaphanous auto-regulatory domain (DAD) and the C-terminal tail (CT). DRFs are regulated by auto-inhibition, where binding of the DAD to the DID domain keeps the molecule closed and inactive. Binding of an activated Rho GTPase to the GBD releases the DAD from DID, subsequently opening and activating the protein. (B) Schematic model of the growth cone cytoskeleton. (B’, B”)Proposed functions of DAAM in neuronal growth cones: ① actin barbed-end binding,actin assembly, ② F-actin bundling, ③F-actin and MT coalignment, ④ EB1 binding, ⑤facilitated actin filament formation at the MT plus-ends through interactions with EB1.
Molecular mechanisms of formin mediated actin-MT crosstalk:Interestingly, the major MT interaction domain of formins appears to be the FH2 domain (also required for actin binding),although some formins contain an additional MT binding motif in their C-terminal tail (CT) region (also implicated in actin interactions). Thus, formins contain multiple actin and MT binding regions which place them into an ideal position to crosslink the actin and MT cytoskeleton directly. On the other hand, some formins were shown to bind or form complexes with MT binding proteins, such as +TIPs, providing a possibility for indirect MT binding. Together, these findings raised numerous points that need to be resolved. With regard to MT binding domains, although not conserved in all formins, the case of the CT domain is clear in the sense that it is capable of ionic interactions with the acidic MT surface. By contrast, little is known about how the FH2 domain binds to MTs. Conserved amino acid residues in the FH2 domain do not appear to in fluence MT binding in vitro in mDia1, Capu or DAAM (Bartolini et al., 2008; Roth-Johnson et al., 2014; Szikora et al., 2017),and no other motifs are implicated in mediating interactions between MTs and formin FH2 domains. In the absence of available mutations that would selectively impair MT binding, it has not been possible to assess the in vivo relevance of direct MT binding. However, in order to clearly discriminate the actinand MT-speci fic functions of formins, structural studies focusing on mapping MT-binding surfaces of FH2 domains would be a particular interest of the field. Because the FH2 domain is involved in actin and MT binding as well, another critical question is whether the binding of these two cytoskeletal elements is mutually exclusive/competitive or simultaneous. In some cases formins were shown to bind to MTs and actin via overlapping surfaces, therefore they compete for formin binding (Gaillard et al., 2011; Roth-Johnson et al., 2014). Whereas some formins promote the co-alignment of actin and MT filaments suggesting they interact with actin and MTs via different surfaces (Gaillard et al., 2011; Szikora et al., 2017). Thus, current data provide support for both scenarios, presumably suggesting a context/formin dependent regulation. However, the in vivo signi ficance of these interactions needs to be evaluated further.
Formin mediated regulation of MT dynamics in neurons:Stabilization of pioneer MTs in the growth cone periphery is thought to be a key step in axon guidance. Accordingly, orientation of the pioneer MTs in the growth cone seem to predict the direction of neurite turning. Furthermore, the localized application of the MT stabilizing drug, taxol, is sufficient to induce growth cone turning. The dynamic instability of MTs is characterized by the speed and proportion of growth and shrinkage rates of their growing ends. These events can be directly regulated at growing ends by +TIPs or indirectly by lattice binders capable of stabilizing or destabilizing MT protofilaments. In vitro studies have shown that the main effect of formins on MT dynamics is to contribute to stabilization, most likely via side binding of the filaments. Some formins are capable of protecting preassembled MTs against depolymerization, or by inducing the formation of stable MTs or bundle MT filaments, which may also promote MT stabilization (Bartolini et al., 2008; Gaillard et al., 2011; Szikora et al., 2017). The side binding of formins seems to slightly reduce the MT growth rate and strongly reduce shrinkage. The increased lifetime could facilitate posttranslational modification (acetylation, detyrosination) of the MTs affecting the binding of MAPs, which in turn can regulate the dynamic instability of the MTs.
An alternative mode of MT stabilization by formins would be through the +TIP complex. Notably, mDia1 and mDia2 were shown to interact with EB1 and APC via their FH1-FH2 region. The formation of this complex promotes MT stabilization, which has an important role in cell migration (Wen et al.,2004). Remarkably, mDia2 appears to promote MT stabilization independently of its actin assembly function (Bartolini et al.,2008). Similar observations were made in Drosophila primary neurons, where we demonstrated that a significant portion of DAAM localizes to MT plus ends promoting MT stabilization(independent of its actin nucleation activity), and co-purifies with EB1 from S2 cell lysates (Szikora et al., 2017). Based on our unpublished data, MT stabilization is only achieved with truncated constructs relieved from auto-inhibition, whereas the full length form was inactive in this assay. These observations suggest that the auto-inhibitory interaction typical for the DRF family may not simply block actin nucleation, but also prevents certain types of MT interactions required for stabilization. If proven, this could represent a novel way to coordinate actin assembly and MT organization by activating a single formin type of protein. Signi ficantly, the above mentioned +TIPs bind to the FH1-FH2 region of formins, although the exact binding sites and molecular hierarchy of the formin/+TIP complex formation are not well defined yet. The involvement of the FH2 domain, not only in actin and MT but also in +TIP binding,suggests the existence of a complex regulatory system. This system consists of formins and formin-interacting proteins that build an intricate system to regulate multiple aspects of MT dynamics and actin-MT crosstalk. In this regard, an important recent study demonstrated that an mDia1/CLIP-170/EB1 module triggers an accelerated actin polymerization from MT plus ends(Henty-Ridilla et al., 2016). These authors also found CLIP-170 increased the elongation rate of actin filaments assembled by other formins (mDia2, DAAM1, INF1 and INF2) suggesting a general regulatory function for CLIP-170 in formin-mediated actin polymerization. This discovery provides further proof for the versatile regulatory potential of formins coupled to +TIPs.Nevertheless, the exact composition, speci ficity and regulation of these complexes in neurons and other cell types remain to be determined. It would be important to see if the +TIP/formin interactions are tissue, cell type, developmental stage or context dependent. Is there specificity of regulation at this level or is there the possibility of multiple, highly redundant interactions ?
Guidance signaling:In order to find their correct target sites,neuronal growth cones need to navigate in a complex cellular environment exhibiting a large number of guidance cues. These cues control growth cone behavior orchestrated by coordinated remodeling of the actin and MT cytoskeleton. Given that formins coupled with +TIPs are important elements of connecting the regulation of actin and MT dynamics in neurons, it’s essential to understand the connection between this MT plus end cytoskeletal effector module and navigation systems. Previously we have shown that DAAM is regulated by Rac, downstream of the PCP navigation system in mushroom body neurons (Gombos et al., 2015). However, the Rho GTPase family was linked to all other major axonal guidance systems as well, and additionally, it was implicated in +TIP regulation in several other cellular contexts (Fukata et al., 2002). Taking this together with the expression pattern of vertebrate DAAM orthologs, known to be abundant in the developing central nervous system (CNS),a Rho-DAAM/formin-(+)TIP pathway might serve as an evolutionary conserved, critical component of actin-MT coordination downstream of axonal guidance signaling. Nevertheless,future studies are required to clarify the molecular mechanisms whereby Rho GTPases ensure a concerted regulation of a number of +TIPs and possibly a number of formins, under the control of guidance signaling.
Conclusions:Formins, as well as +TIPs, were both known to be capable of mediating actin-MT interactions in variousin vitroand cellular systems. Curiously, research during the past few years has established that despite their independent abilities to link actin filaments to MTs, +TIPs often collaborate with formins to induce coordinated changes of the two major cytoskeletal systems. Thus, a +TIP/formin based module has emerged as a general mechanism of linking actin and MT dynamics, which is essential for several cellular processes such as migration, phagocytosis, neuronal dendritic branching and axonal growth (Wen et al., 2004; Henty-Ridilla et al., 2016; Szikora et al., 2017). Due to the physiological importance of this mode of cytoskeleton regulation, particularly in the context of neuronal growth and regeneration, further biochemical and most importantlyin vivostudies are required to understand how these cytoskeleton regulatory elements associate with each other and how upstream signals control their activity.
This work was supported by the Hungarian Science Foundation (OTKA) (K109330 to JM), the Hungarian Brain Research Program (KTIA_NAP_13-2-2014-0007 to JM), the National Research, Development and Innovation Office (GINOP-2.3.2-15-2016-00001 and GINOP-2.3.2-15-2016-00032 to JM), and by an MTA Postdoctoral Fellowship (to IF).
István Földi, Szilárd Szikora, József Mihály*Institute of Genetics, Biological Research Centre, Hungarian Academy of Sciences, MTA-SZBK NAP B Axon Growth and Regeneration Group, Szeged, Hungary
*Correspondence to:József Mihály, mihaly.jozsef@brc.mta.hu.
orcid:0000-0003-3399-2424 (József Mihály)
How to cite this article:Földi I, Szikora S, Mihály J (2017) Formin’bridges between microtubules and actin filaments in axonal growth cones.Neural Regen Res 12(12):1971-1973.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 3.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as the author is credited and the new creations are licensed under identical terms.
Open peer review report:Reviewer: Yen-chung Chang, National Tsing Hua University, China.
Comments to authors: It is a nice and short reviewer article. This article summarizes the most recent advancements regarding the role(s) played by formin in joining the actin and microtubule filaments in axonal growth cones. In addition, the author also point out several possible lines of related research in the future, including the studies of how formins interact with +TIP proteins and of how formin/+TIP complex contributes to linking the actin and microtubule filaments in growth cones. The findings of several recent and relevant articles relating to the above-described subject have been discussed in the article. This article also includes a good background introduction which helps guide readers from different fields to read through the article.
Bartolini F, Gundersen GG (2010) Formins and microtubules. Biochim Biophys Acta 1803:164-173.
Bartolini F, Moseley JB, Schmoranzer J, Cassimeris L, Goode BL, Gundersen GG (2008) The formin mDia2 stabilizes microtubules independently of its actin nucleation activity. J Cell Biol 181:523-536.
Cammarata GM, Bearce EA, Lowery LA (2016) Cytoskeletal social networking in the growth cone: How +TIPs mediate microtubule-actin cross-linking to drive axon outgrowth and guidance. Cytoskeleton (Hoboken)73:461-476.
Daou P, Hasan S, Breitsprecher D, Baudelet E, Camoin L, Audebert S, Goode BL, Badache A (2014) Essential and nonredundant roles for Diaphanous formins in cortical microtubule capture and directed cell migration. Mol Biol Cell 25:658-668.
Fukata M, Watanabe T, Noritake J, Nakagawa M, Yamaga M, Kuroda S, Matsuura Y, Iwamatsu A, Perez F, Kaibuchi K (2002) Rac1 and Cdc42 capture microtubules through IQGAP1 and CLIP-170. Cell 109:873-885.
Gaillard J, Ramabhadran V, Neumanne E, Gurel P, Blanchoin L, Vantard M,Higgs HN (2011) Differential interactions of the formins INF2, mDia1,and mDia2 with microtubules. Mol Biol Cell 22:4575-4587.
Gombos R, Migh E, Antal O, Mukherjee A, Jenny A, Mihály J (2015) The formin DAAM functions as molecular effector of the planar cell polarity pathway during axonal development in Drosophila. J Neurosci 35:10154-10167.doi:10.1523/JNEUROSCI.3708-14.2015.
Henty-Ridilla JL, Rankova A, Eskin JA, Kenny K, Goode BL (2016) Accelerated actin filament polymerization from microtubule plus ends. Science 352:1004-1009.
Preciado López M, Huber F, Grigoriev I, Steinmetz MO, Akhmanova A,Koenderink GH, Dogterom M (2014) Actin-microtubule coordination at growing microtubule ends. Nat Commun 5:4778.
Roth-Johnson EA, Vizcarra CL, Bois JS, Quinlan ME (2014) Interaction between microtubules and the Drosophila formin Cappuccino and its effect on actin assembly. J Biol Chem 289:4395-4404.
Szikora S, Földi I, Tóth K, Migh E, Vig A, Bugyi B, Maléth J, Hegyi P, Kaltenecker P, Sanchez-Soriano N, Mihály J (2017) The formin DAAM is required for coordination of the actin and microtubule cytoskeleton in axonal growth cones. J Cell Sci 130:2506-2519.
Wen Y, Eng CH, Schmoranzer J, Cabrera-Poch N, Morris EJ, Chen M, Wallar BJ, Alberts AS, Gundersen GG (2004) EB1 and APC bind to mDia to stabilize microtubules downstream of Rho and promote cell migration. Nat Cell Biol 6:820-830.
2017-09-26
10.4103/1673-5374.221148
杂志排行

中国神经再生研究(英文版)的其它文章
- Roles of neural stem cells in the repair of peripheral nerve injury
- Conductive polymer scaffolds to improve neural recovery
- The contribution of oligodendrocytes and oligodendrocyte progenitor cells to central nervous system repair in multiple sclerosis: perspectives for remyelination therapeutic strategies
- MicroRNAs in Parkinson’s disease and emerging therapeutic targets
- Surgical reconstruction of spinal cord circuit provides functional return in humans
- Environmental cues determine the fate of astrocytes after spinal cord injury